
Analysis of Citrus Tristeza Virus Incidences within Asian Citrus Psyllid (Diaphorina citri) Populations in Florida via High-Throughput Sequencing

 , , and
, , and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Collection for RNA Extraction and HTS Analysis
2.2. High-Throughput Sequencing (HTS) Assembly and Analysis
2.3. Statistical Analysis
2.4. Validation of CTV Presence in HTS Samples and Field ACPs by RT-PCR
2.5. CTV Strain Detection in Florida ACPs by Strain-Specific RT-PCR
2.6. Immunogold Labelling of CTV Virions
3. Results
3.1. HTS Generated a Persistent Presence of CTV Contigs in Total RNAs Extracted from Florida ACPs
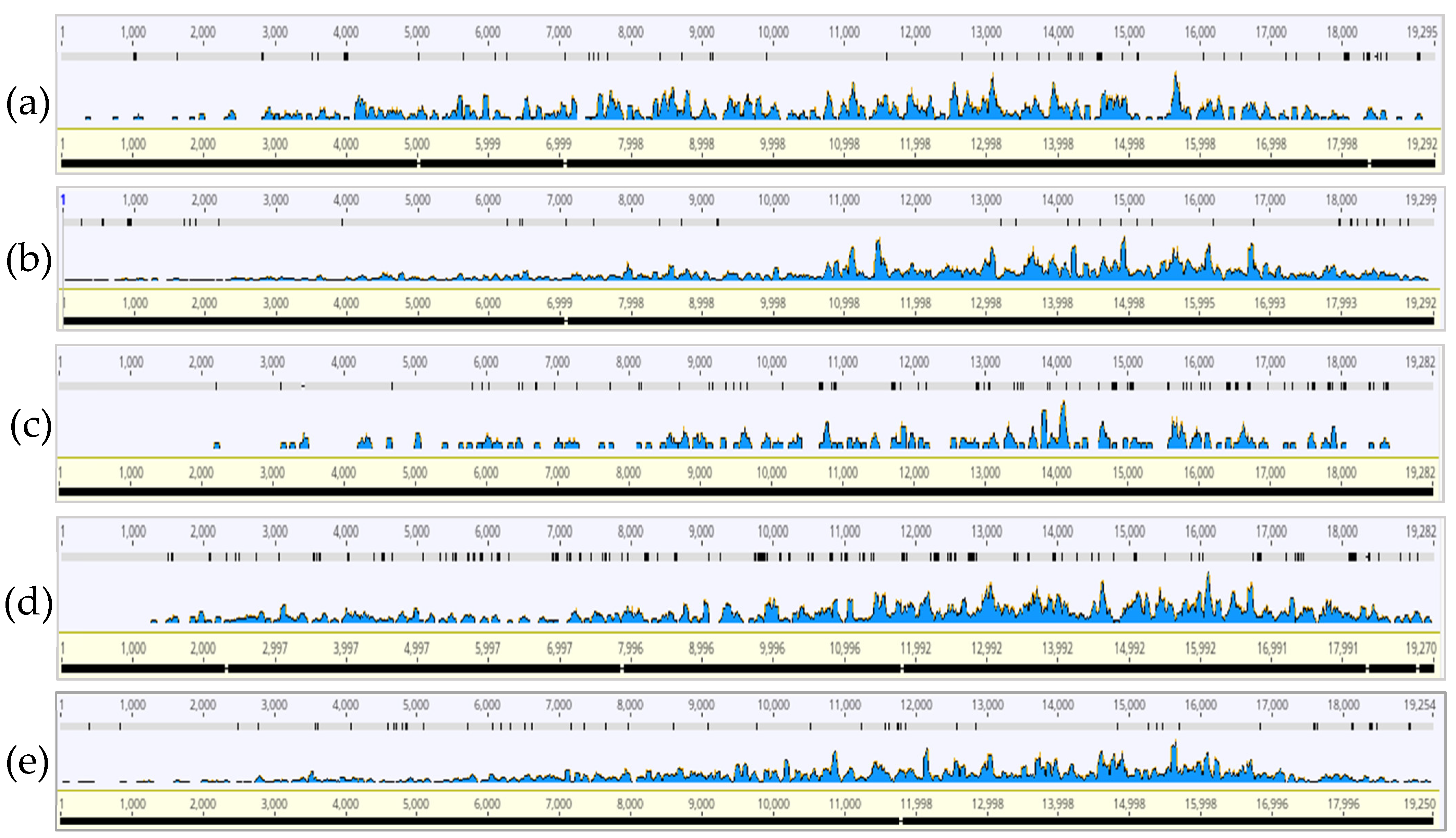
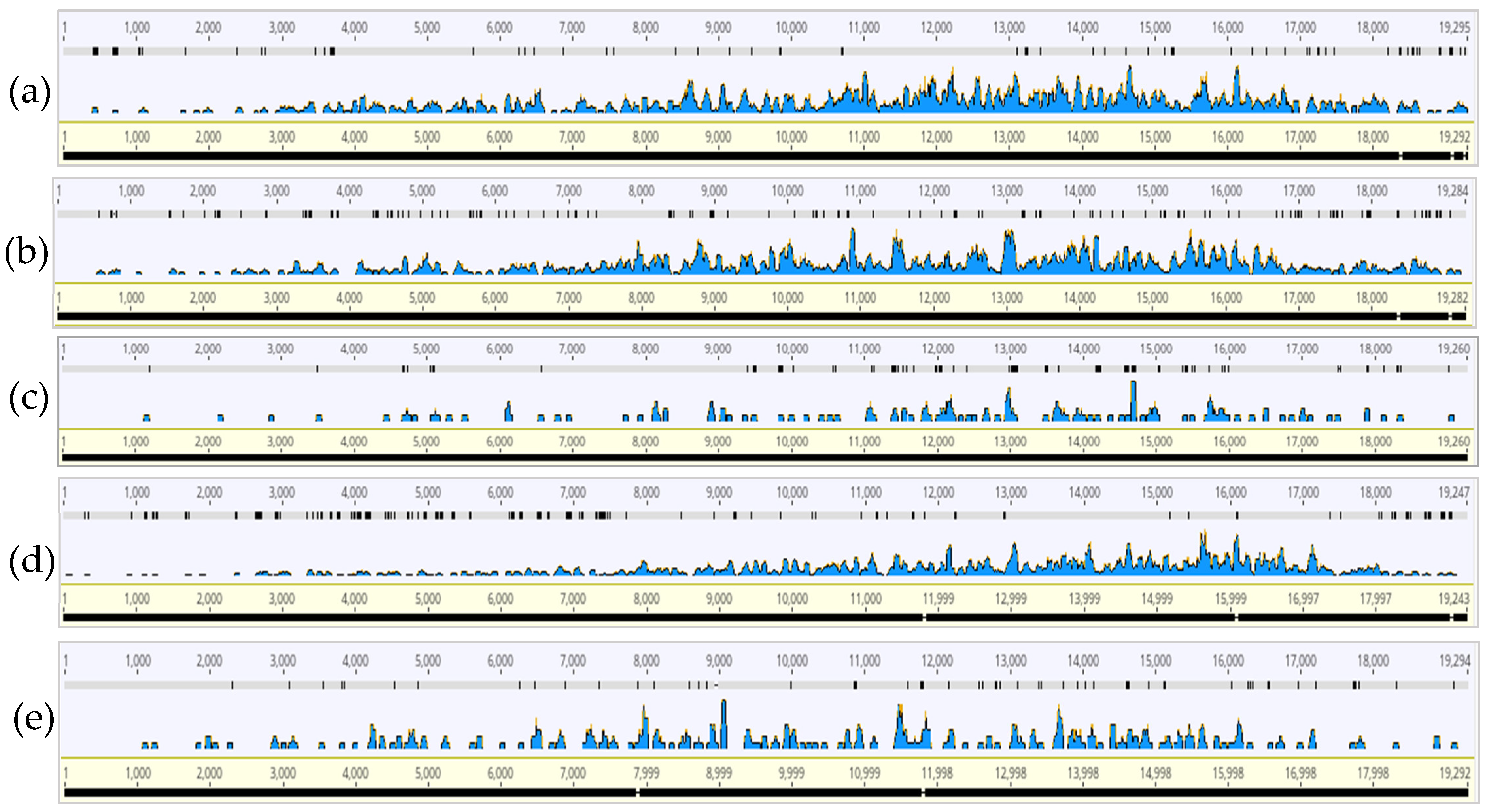
3.2. CTV Read Coverage Profiles of HTS Samples
3.3. RT-PCRs of Florida ACPs
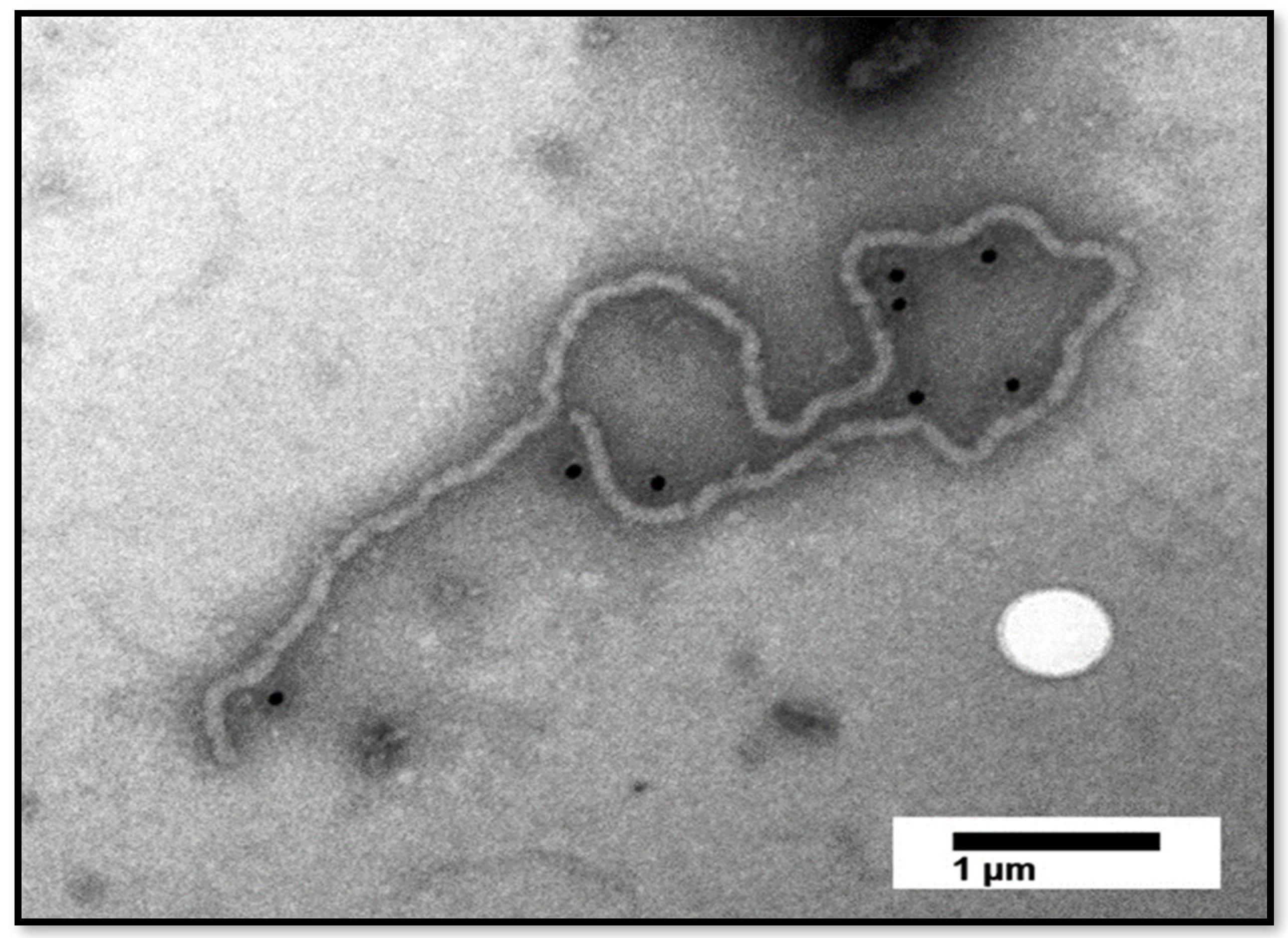
3.4. Immunogold-Labelling Detects the Presence of Large CTV Particles in Florida ACPs
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bové, J.M. Huanglongbing: A Destructive, Newly-Emerging, Century-Old Disease of Citrus. J. Plant Pathol. 2006, 88, 7–37. [Google Scholar] [CrossRef]
- Halbert, S.E.; Niblett, C.; Manjunath, K.L.; Lee, R.F.; Brown, L.G. Establishment of two new vectors of citrus pathogens in Florida. In Proceedings of the International Society of Citriculture IX Congress, Orlando, FL, USA, 3–7 December 2000; ASHS Press: Alexandria, VA, USA, 2003; pp. 1016–1017. [Google Scholar]
- Graham, J.; Gottwald, T.; Setamou, M. Status of Huanglongbing (HLB) outbreaks in Florida, California and Texas. Trop. Plant Pathol. 2020, 45, 265–278. [Google Scholar] [CrossRef]
- Nouri, S.; Salem, N.; Nigg, J.C.; Falk, B.W. Diverse Array of New Viral Sequences Identified in Worldwide Populations of the Asian Citrus Psyllid (Diaphorina citri) Using Viral Metagenomics. J. Virol. 2016, 90, 2434–2445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nouri, S.; Matsumura, E.E.; Kuo, Y.W.; Falk, B.W. Insect-specific viruses: From discovery to potential translational applications. Curr. Opin. Virol. 2018, 33, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Marutani-Hert, M.; Hunter, W.B.; Katsar, C.S.; Sinisterra, X.H.; Hall, D.G.; Powell, C.A. Reovirus-like sequences isolated from adult Asian citrus psyllid, (Hemiptera: Psyllidae: Diaphorina citri). Fla. Entomol. 2009, 92, 314–320. [Google Scholar] [CrossRef]
- Matsumura, E.E.; Nerva, L.; Nigg, J.C.; Falk, B.W.; Nouri, S. Complete genome sequence of the largest known flavi-like virus, Diaphorina citri flavi-like virus, a novel virus of the Asian citrus psyllid, Diaphorina citri. Genome Announc. 2016, 4, e00946-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nigg, J.C.; Nouri, S.; Falk, B.W. Complete genome sequence of a putative densovirus of the Asian Citrus Psyllid, Diaphorina citri. Genome Announc. 2016, 4, e00589-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varghese, F.S.; van Rij, R.P. Insect virus discovery by metagenomic and cell culture-based approaches. In Viral Metagenomics: Methods and Protocols, Methods in Molecular Biology; Pantaleo, V., Chiumenti, M., Eds.; Humana Press: New York, NY, USA, 2018; Volume 1746, pp. 197–213. ISBN 9781493976836. [Google Scholar]
- Britt, K.; Gebben, S.; Levy, A.; Al Rwahnih, M.; Batuman, O. The Detection and Surveillance of Asian Citrus Psyllid (Diaphorina citri)—Associated Viruses in Florida Citrus Groves. Front. Plant Sci. 2020, 10, 1687. [Google Scholar] [CrossRef] [Green Version]
- Dawson, W.O.; Garnsey, S.M.; Tatineni, S.; Folimonova, S.Y.; Harper, S.J.; Gowda, S. Citrus tristeza virus-host interactions. Front. Microbiol. 2013, 4, 88. [Google Scholar] [CrossRef] [Green Version]
- Grant, T.J. Evidence of tristeza, or quick decline, virus in Florida. Proc. Fla. State Hortic. Soc. 1952, 65, 28–31. [Google Scholar]
- Bar-Joseph, M.; Batuman, O.; Roistacher, C.N. The History of Citrus tristeza virus—Revisited. In Citrus Tristeza Virus Complex and Tristeza Diseases; Karasev, A.V., Hilf, M.E., Eds.; APS Press: St. Paul, MN, USA, 2010; pp. 3–26. [Google Scholar]
- Bar-Joseph, M.; Marcus, R.; Lee, R.F. Citrus Tristeza Virus. Annu. Rev. Phytopathol. 1989, 27, 291–316. [Google Scholar] [CrossRef]
- Bowman, K.D. Taking Stock of New Citrus Rootstocks. Grow. Prod. 2010. Available online: https://www.growingproduce.com/citrus/varieties-rootstocks/taking-stock-of-new-citrus-rootstocks/ (accessed on 10 April 2020).
- Folimonova, S.Y.; Robertson, C.J.; Garnsey, S.M.; Gowda, S.; Dawson, W.O. Examination of the responses of different genotypes of citrus to huanglongbing (Citrus Greening) under different conditions. Phytopathology 2009, 99, 1346–1354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grosser, J.W.; Gmitter, F.; Bowman, K.D. New rootstocks in the citrus breeding pipeline. Citrus Ind. 2020, 101, 8–11. [Google Scholar]
- Halbert, S.E.; Genc, H.; Cevik, B.; Brown, L.G.; Rosales, I.M.; Manjunath, K.L.; Pomerinke, M.; Davison, D.A.; Lee, R.F.; Niblett, C.L. Distribution and characterization of Citrus tristeza virus in South Florida following establishment of Toxoptera citricida. Plant Dis. 2004, 88, 935–941. [Google Scholar] [CrossRef] [PubMed]
- Harper, S.J.; Cowell, S.J.; Halbert, S.E.; Brlansky, R.H.; Dawson, W.O. CTV status in Florida. Citrus Ind. 2015, 36, 8–14. [Google Scholar]
- Harper, S.J.; Cowell, S.J. The past and present status of Citrus tristeza virus in Florida. J. Citrus Pathol. 2016, 3, 1–6. [Google Scholar] [CrossRef]
- Al Rwahnih, M.; Rowhani, A.; Westrick, N.; Stevens, K.; Diaz-Lara, A.; Trouillas, F.P.; Preece, J.; Kallsen, C.; Farrar, K.; Golino, D. Discovery of Viruses and Virus-like Pathogens in Pistachio using High-Throughput Sequencing. Plant Dis. 2018, 102, 1419–1425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [Green Version]
- Britt, K.; Stevens, K.; Gebben, S.; Levy, A.; Al Rwahnih, M.; Batuman, O. Partial Genome Sequence of a Novel Reo-Like Virus Detected in Asian Citrus Psyllid (Diaphorina citri) Populations from Florida Citrus Groves. Microbiol. Resour. Announc. 2021, 10, e00563-21. [Google Scholar] [CrossRef] [PubMed]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, C.; Manimaran, S.; Shen, Y.; Perez-Rogers, J.F.; Byrd, A.L.; Castro-Nallar, E.; Crandall, K.A.; Johnson, W.E. PathoScope 2.0: A complete computational framework for strain identification in environmental or clinical sequencing samples. Microbiome 2014, 2, 33. [Google Scholar] [CrossRef] [PubMed]
- Nouri, S.; Salem, N.; Falk, B.W. Complete genome sequence of Diaphorina citri-associated C virus, a novel putative RNA virus of the Asian Citrus Psyllid, Diaphorina citri. Genome Announc. 2016, 4, e00639-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [PubMed]
- Kruskal, W.H.; Wallis, W.A. Use of Ranks in One-Criterion Variance Analysis. J. Am. Stat. Assoc. 1952, 47, 583–621. [Google Scholar] [CrossRef]
- Dunn, O.J. Multiple Comparisons Using Rank Sums. Technometrics 1964, 6, 241–252. [Google Scholar] [CrossRef]
- Hilf, M.E.; Mavrodieva, V.A.; Garnsey, S.M. Genetic marker analysis of a global collection of isolates of Citrus tristeza virus: Characterization and distribution of CTV genotypes and association with symptoms. Phytopathology 2005, 95, 909–917. [Google Scholar] [CrossRef] [Green Version]
- Bernad, L.; Duran-Vila, N. A novel RT-PCR approach for detection and characterization of citrus viroids. Mol. Cell. Probes 2006, 20, 105–113. [Google Scholar] [CrossRef]
- Harper, S.J.; Cowell, S.J.; Dawson, W.O. With a little help from my friends: Complementation as a survival strategy for viruses in a long-lived host system. Virology 2015, 478, 123–128. [Google Scholar] [CrossRef] [Green Version]
- Dombrovsky, A.; Luria, N. The Nerium oleander aphid Aphis nerii is tolerant to a local isolate of Aphid lethal paralysis virus (ALPV). Virus Genes 2013, 46, 354–361. [Google Scholar] [CrossRef]
- Yokomi, R.K.; Selvaraj, V.; Maheshwari, Y.; Saponari, M.; Giampetruzzi, A.; Chiumenti, M.; Hajeri, S. Identification and characterization of Citrus tristeza virus isolates breaking resistance in Trifoliate Orange in California. Phytopathology 2017, 107, 901–908. [Google Scholar] [CrossRef] [Green Version]
- Bester, R.; Cook, G.; Maree, H.J. Citrus tristeza virus genotype detection using high-throughput sequencing. Viruses 2021, 13, 168. [Google Scholar] [CrossRef] [PubMed]
- Licciardello, G.; Scuderi, G.; Ferraro, R.; Giampetruzzi, A.; Russo, M.; Lombardo, A.; Raspagliesi, D.; Bar-Joseph, M.; Catara, A. Deep sequencing and analysis of small RNAs in sweet orange grafted on sour orange infected with two citrus tristeza virus isolates prevalent in Sicily. Arch. Virol. 2015, 160, 2583–2589. [Google Scholar] [CrossRef] [PubMed]
- Matsumura, E.E.; Coletta-Filho, H.D.; Nouri, S.; Falk, B.W.; Nerva, L.; Oliveira, T.S.; Dorta, S.O.; Machado, M.A. Deep sequencing analysis of RNAs from citrus plants grown in a citrus sudden death–affected area reveals diverse known and putative novel viruses. Viruses 2017, 9, 92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruiz-Ruiz, S.; Navarro, B.; Gisel, A.; Peña, L.; Navarro, L.; Moreno, P.; Di Serio, F.; Flores, R. Citrus tristeza virus infection induces the accumulation of viral small RNAs (21-24-nt) mapping preferentially at the 3′-terminal region of the genomic RNA and affects the host small RNA profile. Plant Mol. Biol. 2011, 75, 607–619. [Google Scholar] [CrossRef] [Green Version]
- Batuman, O.; Che, X.; Yang, G.; Mawassi, M.; Bar-Joseph, M. Interference or Insurance? More Questions than Answers on the Roles of the Multiple Defective RNAs of Citrus tristeza virus. In Citrus Tristeza Virus Complex and Tristeza Diseases; Karasev, A.V., Hilf, M.E., Eds.; APS Press: St. Paul, MN, USA, 2010; pp. 73–93. [Google Scholar]
- Dodds, J.A.; Bar-Joseph, M. Double-Stranded RNA from Plants Infected with Closteroviruses. Phytopathology 1983, 73, 419–423. [Google Scholar] [CrossRef]
- Hilf, M.E.; Karasev, A.V.; Pappu, H.R.; Gumpf, D.J.; Niblett, C.L.; Garnsey, S.M. Characterization of Citrus Tristeza Virus Subgenomic RNAs in Infected Tissue. Virology 1995, 208, 576–582. [Google Scholar] [CrossRef] [Green Version]
- Bordenstein, S.R.; Wernegreen, J.J. Bacteriophage flux in endosymbionts (Wolbachia): Infection frequency, lateral transfer, and recombination rates. Mol. Biol. Evol. 2004, 21, 1981–1991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masui, S.; Kamoda, S.; Sasaki, T.; Ishikawa, H. Distribution and evolution of bacteriophage WO in Wolbachia, the endosymbiont causing sexual alterations in Arthropods. J. Mol. Evol. 2000, 51, 491–497. [Google Scholar] [CrossRef]
- Schneider, W.; Roy, A.; Hartung, J.; Shao, J.; Brlansky, R. Citrus blight research update. Citrus Ind. 2015, 2015, 12–14. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Group | Region | Florida Counties Included | The Approximate Number of Nymph and Adult ACPs Used for Composite |
|---|---|---|---|
| 2019 | A | Lake, Orange, Seminole | 20 |
| 2019 | B | Pasco, Polk | 15 |
| 2019 | C | DeSoto, Hardee, Manatee | 20 |
| 2019 | D | Martin, Okeechobee, St. Lucie | 20 |
| 2019 | E | Charlotte, Hendry, Lee, Collier | 25 |
| 2020 | A | Lake, Orange, Osceola, Seminole | 30 |
| 2020 | B | Pasco, Polk | 20 |
| 2020 | C | DeSoto, Highlands, Manatee, Sarasota | 50 |
| 2020 | D | Indian River, Okeechobee, St. Lucie | 30 |
| 2020 | E | Charlotte, Glades, Hendry, Lee, Collier | 50 |
| Group | Region | Total Yield of Reads | Total Filtered Reads * | Assembled Contigs ≥200 nts |
|---|---|---|---|---|
| 2019 | A | 2,687,116,425 | 35,828,219 | 52,675 |
| 2019 | B | 2,794,686,600 | 37,262,488 | 60,531 |
| 2019 | C | 1,545,277,950 | 20,603,706 | 45,251 |
| 2019 | D | 2,835,048,375 | 37,800,645 | 64,968 |
| 2019 | E | 2,997,707,400 | 39,969,432 | 51,178 |
| 2020 | A | 2,878,296,525 | 38,377,287 | 50,506 |
| 2020 | B | 2,909,774,550 | 38,796,994 | 59,991 |
| 2020 | C | 3,180,154,650 | 42,402,062 | 50,093 |
| 2020 | D | 1,611,839,475 | 21,491,193 | 48,713 |
| 2020 | E | 1,440,971,850 | 19,212,958 | 44,465 |
| Virus | Region A | Region B | Region C | Region D | Region E |
|---|---|---|---|---|---|
| CTV 1 | 99.38, 4680, 0.24X | 98.25, 27,369, 1.42X | - | 98.31, 7739, 0.40X | 99.65, 12,742, 0.66X |
| DcACV 2 | 99.90, 7342, 1.75X | 99.91, 7662, 1.82X | 99.73, 4092, 0.98X | - | 99.74, 4187, 0.99X |
| DcFLV 3 | 97.89, 24,970, 0.90X | 98.89, 27,530, 0.99X | 99.00, 1601, 0.60X | - | - |
| DcDV 4 | - | 84.63, 1438, 0.28X | 84.63, 1382, 0.27X | 91.24, 1659, 0.33X | - |
| WO 5 | - | 92.7, 305, 0.004X | 92.39, 276, 0.004X | - | - |
| Virus | Region A | Region B | Region C | Region D | Region E | Dunn’s Test p < 0.05 * |
|---|---|---|---|---|---|---|
| CTV 1 | T36 6 1.00 (698) | T36 6 12.47 (8969) | T36 7 0.65 (259) | RB 8 2.17 (1580) | VT 9 3.26 (2518) | ab |
| DcACV 2 | 294.80 (44,287) | 120.90 (18,887) | 211.80 (18,298) | 0 (0) | 411.40 (68,947) | a |
| DcFLV 3 | 3.69 (3661) | 47.79 (49,343) | 0.81 (460) | 0 (0) | 0.09 (96) | ab |
| DcDV 4 | 1.16 (211) | 0.85 (460) | 0.91 (95) | 0.96 (184) | 0.81 (164) | ab |
| WO 5 | 0.09 (209) | 0.09 (218) | 0.12 (165) | 0.04 (93) | 0.06 (169) | b |
| Virus | Region A | Region B | Region C | Region D | Region E |
|---|---|---|---|---|---|
| CTV 1 | 98.86, 7628, 0.40X | 97.52, 9518, 0.49X | 100, 276, 0.01X | 97.99, 8445, 0.43X | 100, 427, 0.02X |
| DcACV 2 | 98.60, 7551, 1.80X | 99.62, 4166, 0.99X | 95.35, 7238, 1.74X | 99.80, 6021, 1.43X | 99.95, 4133, 0.98X |
| DcFLV 3 | - | - | 98.36, 66, 0.02X | - | - |
| DcDV 4 | 84.63, 999, 0.20X | 84.63, 1338, 0.60X | - | 84.63, 989, 0.20X | 84.63. 1033, 0.02X |
| DcRV 5 | - | - | - | - | 97.13, 19,515, 1.03X |
| WO 6 | - | - | - | 95.11, 554, 0.008X | - |
| Virus | Region A | Region B | Region C | Region D | Region E | Dunn’s Test p < 0.05 * |
|---|---|---|---|---|---|---|
| CTV 1 | T36 7 1.62 (1200) | T36 8 2.17 (1622) | T30 9 0.21 (170) | VT 10 4.38 (1817) | T36 7 0.71 (263) | ab |
| DcACV 2 | 165.51 (26,634) | 208.69 (33,949) | 51.33 (9126) | 514.51 (46,364) | 229.10 (18,458) | a |
| DcFLV 3 | 0.014 (15) | 0.001 (1) | 0.32 (372) | 0 (0) | 0.15 (78) | b |
| DcDV 4 | 1.16 (226) | 0.84 (166) | 0.44 (95) | 0.68 (74) | 0.78 (76) | ab |
| DcRV 5 | 0.001 (1) | 0 (0) | 0.02 (26) | 0 (0) | 514.19 (262,092) | b |
| WO 6 | 0.10 (241) | 0.05 (131) | 0.10 (268) | 0.13 (190) | 0.10 (131) | b |
| Florida County | ACP Sample (Number of Individuals) | Collection Date | Detected CTV Strains |
|---|---|---|---|
| Glades | Adult (5) | 10/27/2017 | T36, VT |
| Collier | Nymph (11) | 11/1/2017 | T36, VT |
| Orange | Adult (8) | 3/9/2018 | T36, VT |
| Collier | Adult (11) | 3/13/2020 | T30, T36, VT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Britt, K.; Gebben, S.; Levy, A.; Achor, D.; Sieburth, P.; Stevens, K.; Al Rwahnih, M.; Batuman, O. Analysis of Citrus Tristeza Virus Incidences within Asian Citrus Psyllid (Diaphorina citri) Populations in Florida via High-Throughput Sequencing. Insects 2022, 13, 275. https://doi.org/10.3390/insects13030275
Britt K, Gebben S, Levy A, Achor D, Sieburth P, Stevens K, Al Rwahnih M, Batuman O. Analysis of Citrus Tristeza Virus Incidences within Asian Citrus Psyllid (Diaphorina citri) Populations in Florida via High-Throughput Sequencing. Insects. 2022; 13(3):275. https://doi.org/10.3390/insects13030275
Chicago/Turabian StyleBritt, Kellee, Samantha Gebben, Amit Levy, Diann Achor, Peggy Sieburth, Kristian Stevens, Maher Al Rwahnih, and Ozgur Batuman. 2022. "Analysis of Citrus Tristeza Virus Incidences within Asian Citrus Psyllid (Diaphorina citri) Populations in Florida via High-Throughput Sequencing" Insects 13, no. 3: 275. https://doi.org/10.3390/insects13030275
APA StyleBritt, K., Gebben, S., Levy, A., Achor, D., Sieburth, P., Stevens, K., Al Rwahnih, M., & Batuman, O. (2022). Analysis of Citrus Tristeza Virus Incidences within Asian Citrus Psyllid (Diaphorina citri) Populations in Florida via High-Throughput Sequencing. Insects, 13(3), 275. https://doi.org/10.3390/insects13030275