
Antennal Morphology and Fine Structure of Flagellar Sensilla in Hippoboscid Flies with Special Reference to Lipoptena fortisetosa (Diptera: Hippoboscidae)



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Collection
2.2. SEM Procedures
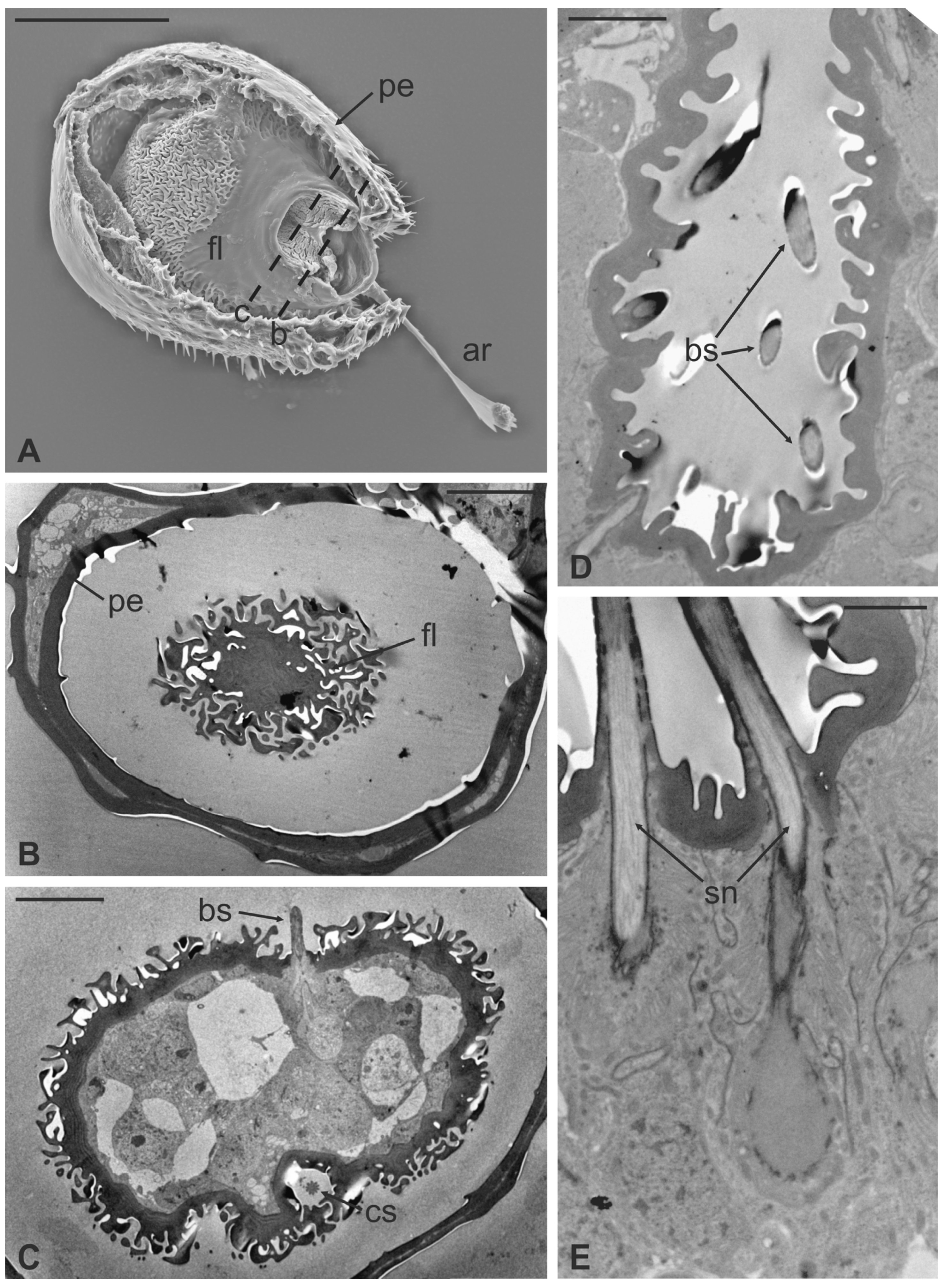
2.3. TEM Procedures
3. Results
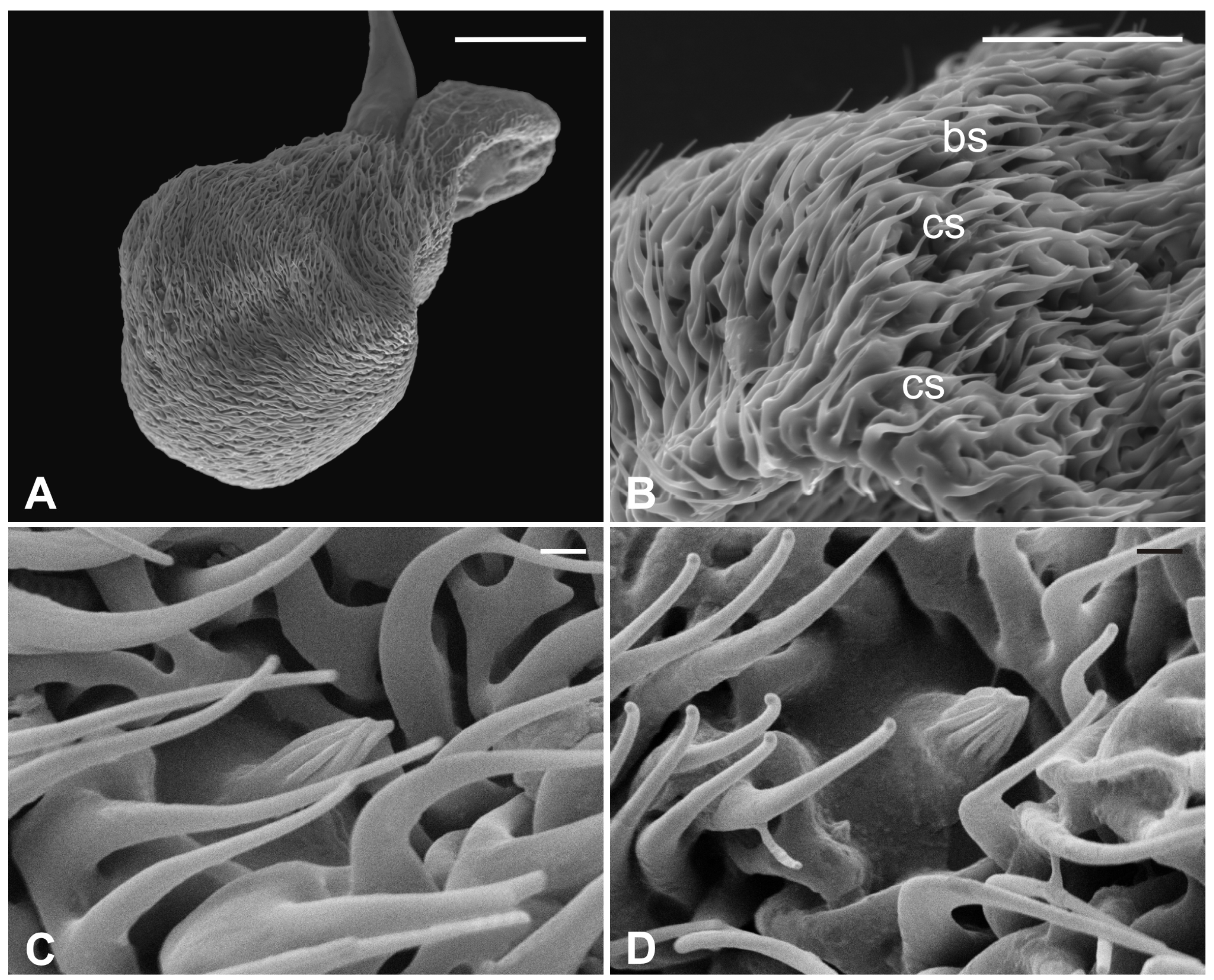
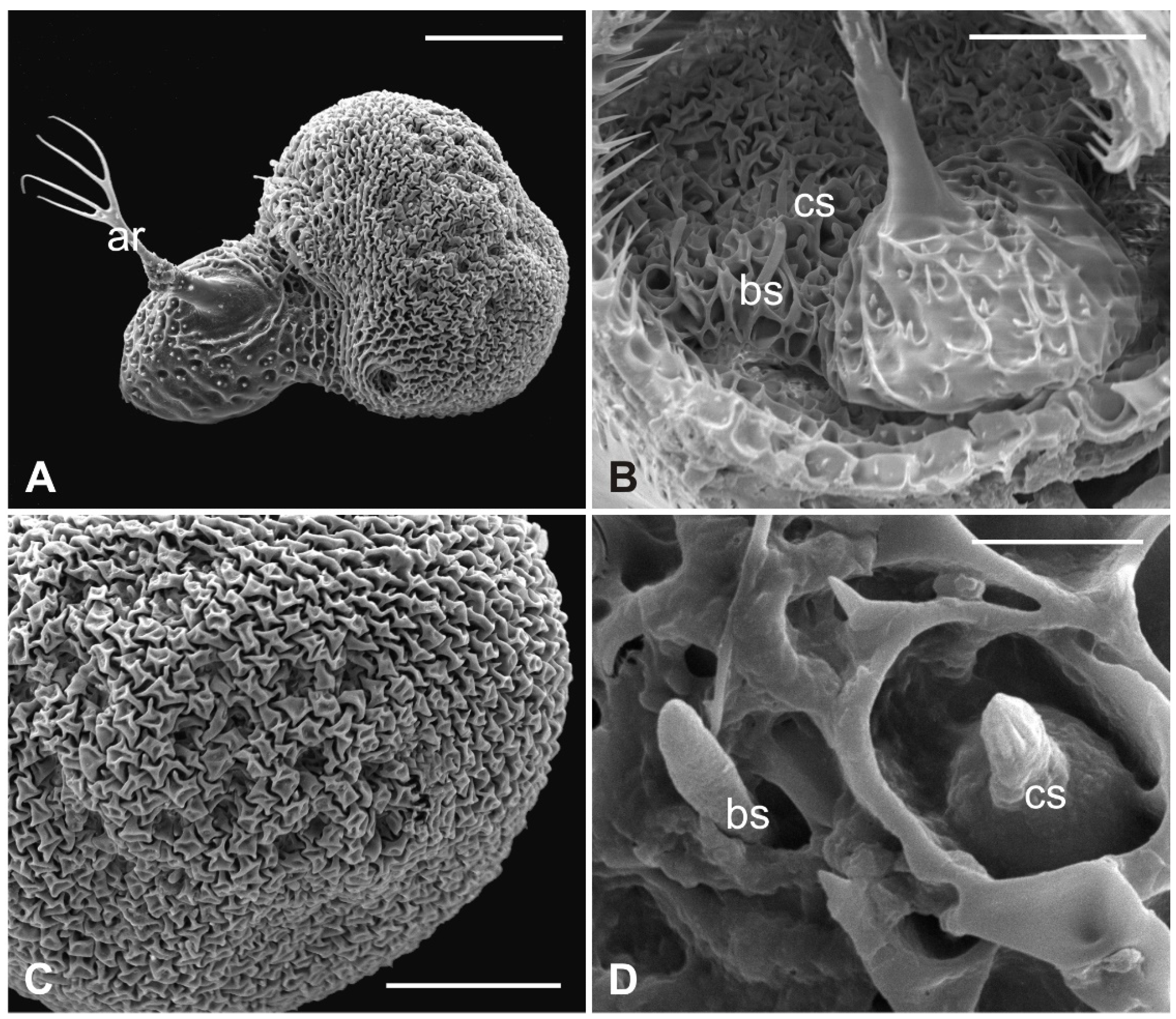
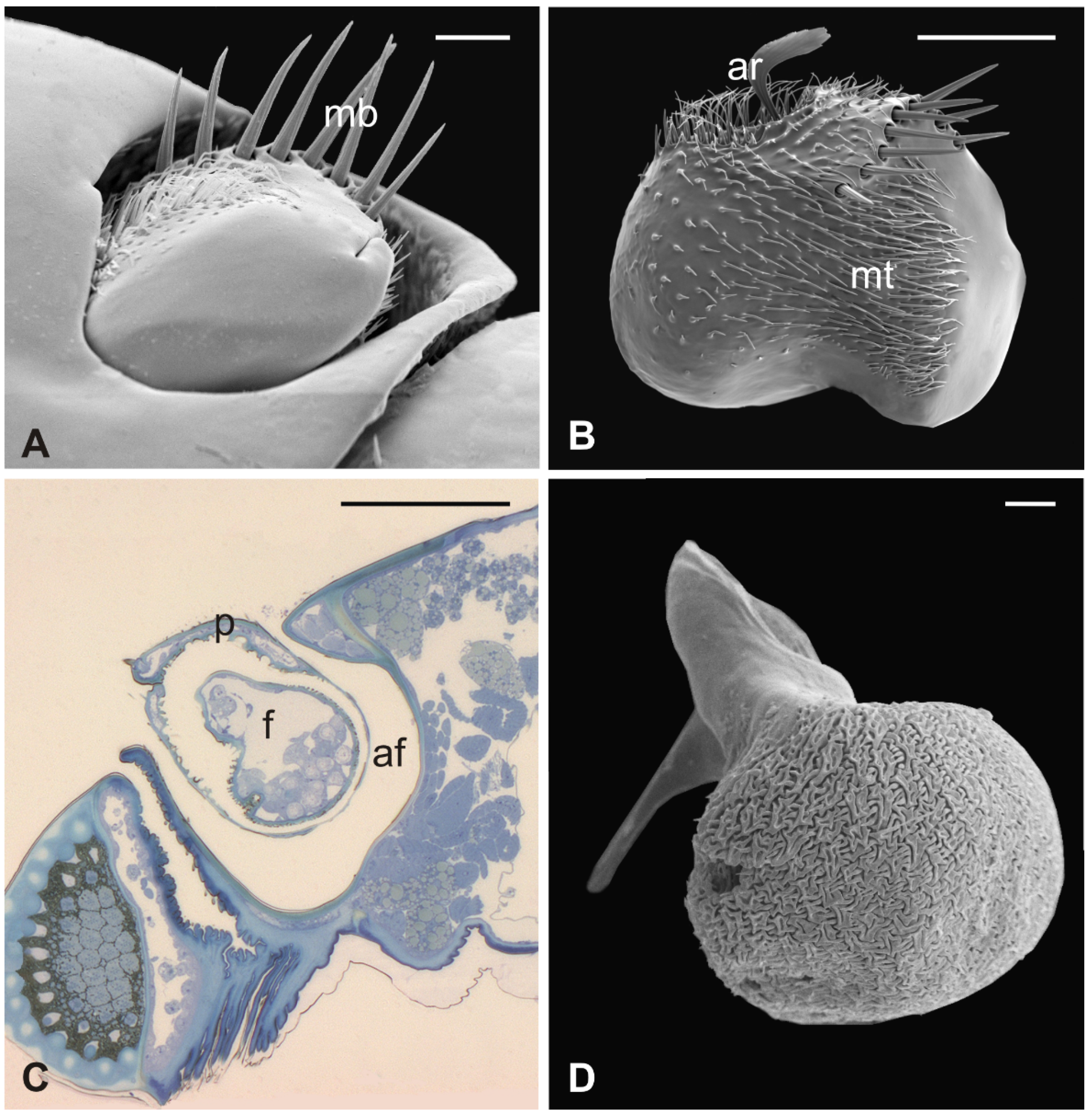
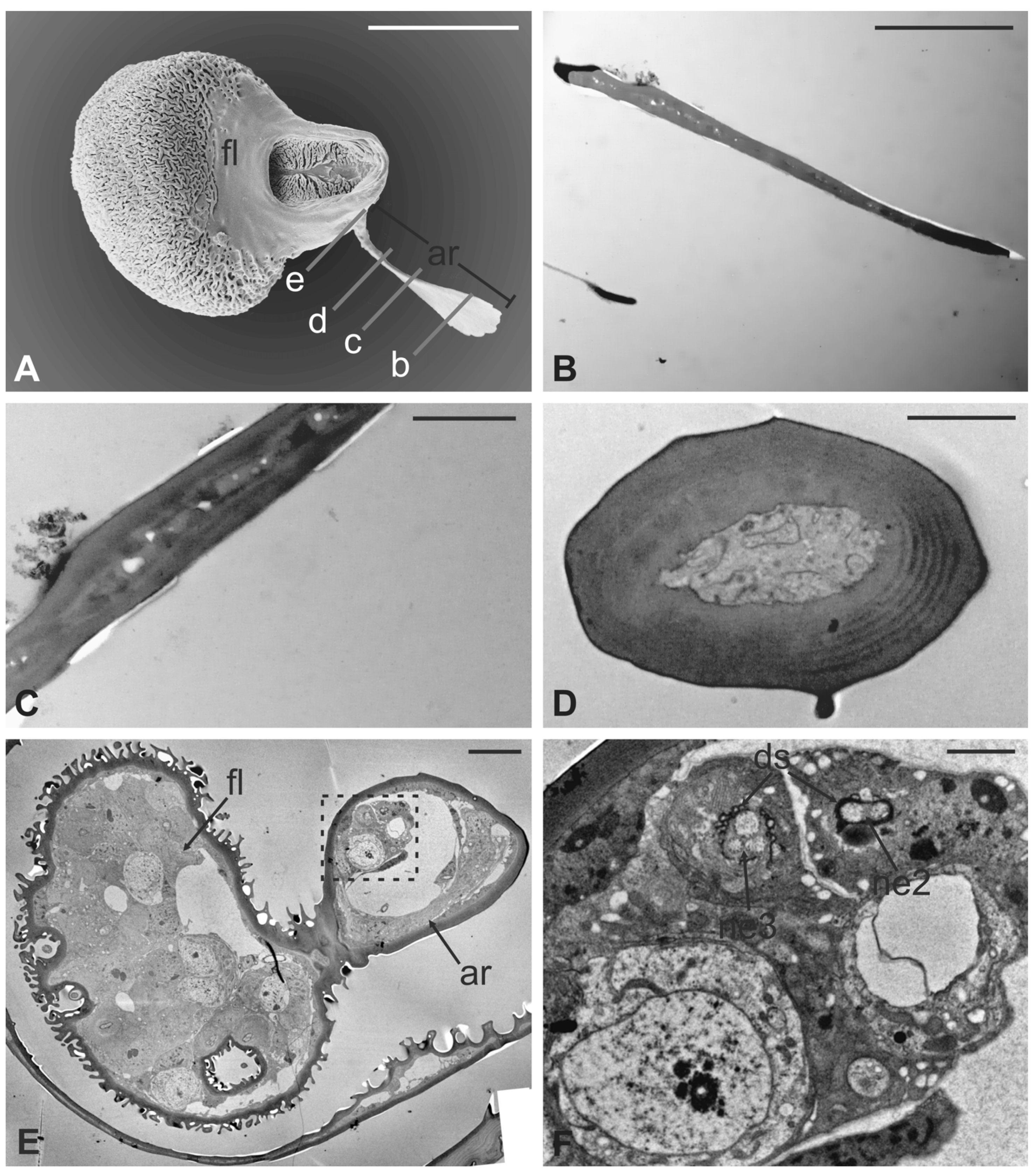
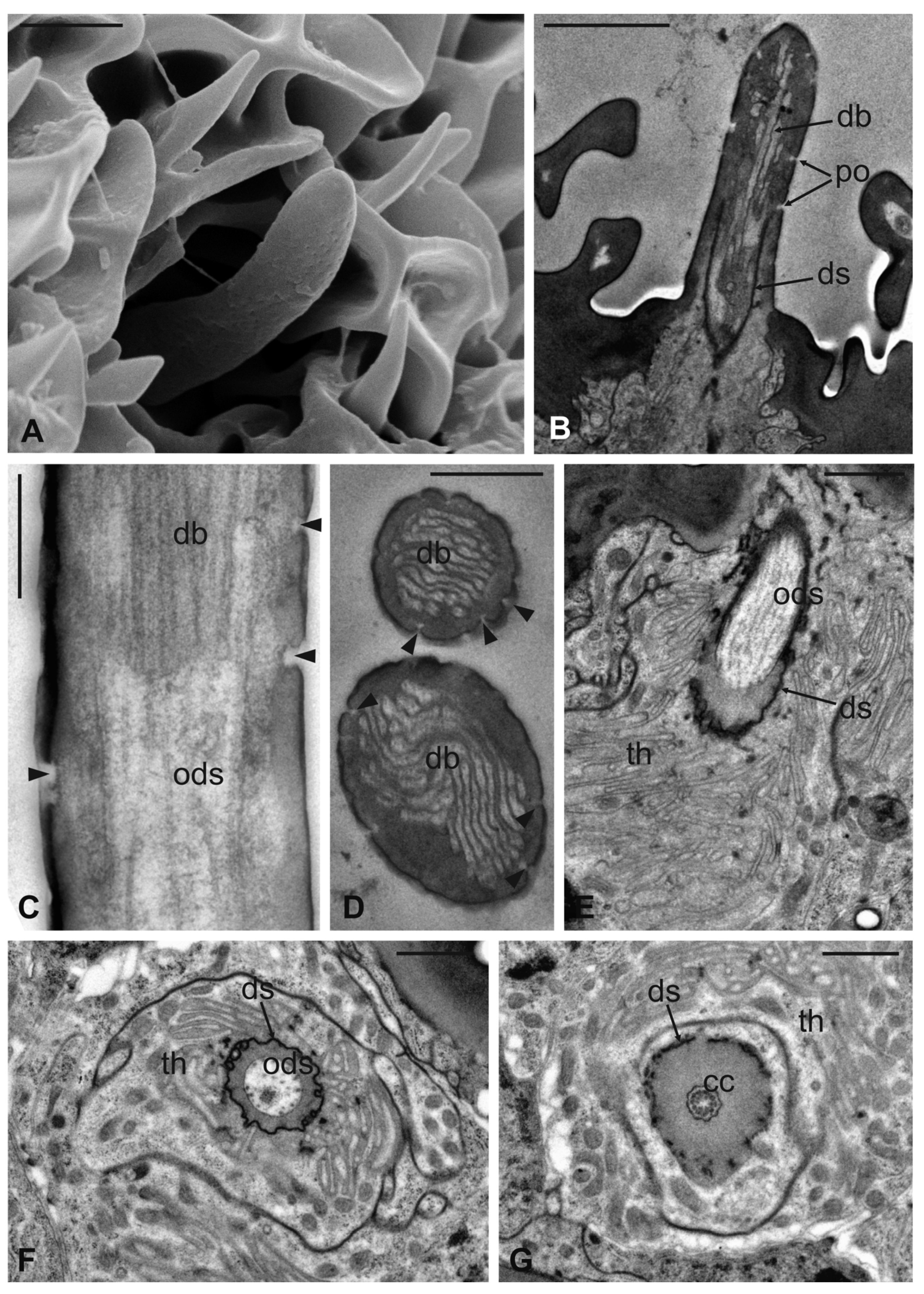
3.1. Pseudolynchia canariensis
3.2. Hippobosca equina
3.3. Lipoptena cervi
3.4. Lipoptena fortisetosa
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Ethical statement
References
- Reeves, W.K.; Lloyd, J.E. Louse flies, keds, and bat flies (Hippoboscoidea). In Medical and Veterinary Entomology, 3rd ed.; Mullen, G.R., Durden, L.A., Eds.; Elsevier: London, UK, 2019; pp. 421–438. [Google Scholar] [CrossRef]
- Hutson, A.M. Keds, flat-flies and bat-flies. Diptera, Hippoboscidae and Nycteribiidae. In Handbooks for the Identification of British Insects; Fitton, M.G., Ed.; Royal Entomological Society of London: London, UK, 1984; Volume 10, pp. 1–40. Available online: https://www.royensoc.co.uk/out-print-handbooks (accessed on 20 April 2021).
- Dick, C.W. Checklist of World Hippoboscidae (Diptera: Hippoboscoidea); Department of Zoology, Field Museum of Natural History: Chicago, IL, USA, 2006; pp. 1–7. Available online: http://fm1.fieldmuseum.org/aa/Files/cdick/Hippoboscidae_Checklist_20dec06.pdf (accessed on 30 January 2020).
- Bezerra-Santos, M.A.; Otranto, D. Keds, the enigmatic flies and their role as vectors of pathogens. Acta Trop. 2020, 209, 105521. [Google Scholar] [CrossRef] [PubMed]
- Dehghani Samani, A.; Pirali Kheirabadi, K.; Dehghani Samani, A. Prevalence and rate of parasitemia of Haemoproteus columbae in Columba livia domestica in Southwest of Iran. Iran. J. Parasitol. 2013, 8, 641–644. Available online: https://ijpa.tums.ac.ir/index.php/ijpa/article/view/443/525 (accessed on 4 January 2022). [PubMed]
- Selmi, R.; Dhibi, M.; Ben Said, M.; Ben Yahia, H.; Abdelaali, H.; Ameur, H.; Baccouche, S.; Gritli, A.; Mhadhbi, M. Evidence of natural infections with Trypanosoma, Anaplasma and Babesia spp. in military livestock from Tunisia. Trop. Biomed. 2019, 36, 742–757. Available online: https://msptm.org/files/Vol36No3/742-757-Selmi-R.pdf (accessed on 4 January 2022). [PubMed]
- Arafa, M.I.; Hamouda, S.M.; Rateb, H.Z.; Abdel-Hafeez, M.M.; Aamer, A.A. Oedematous Skin Disease (OSD) transmission among buffaloes. Glob. J. Med. Res. G Vet. Sci. Vet. Med. 2019, 19, 15–19. Available online: https://globaljournals.org/GJMR_Volume19/3-Oedematous-Skin-Disease.pdf (accessed on 4 January 2022).
- Halos, L.; Jamal, T.; Millard, L.; Girard, B.; Guillot, J.; Chomel, B.; Vayssier-Taussat, M.; Boulouis, H.J. Role of Hippoboscidae flies as potential vectors of Bartonella spp. infecting wild and domestic ruminants. Appl. Environ. Microbiol. 2004, 70, 6302–6305. [Google Scholar] [CrossRef]
- Kaunisto, S.; Kortet, R.; Härkönen, L.; Härkönen, S.; Ylönen, H.; Laaksonen, S. New bedding site examination-based method to analyse deer ked (Lipoptena cervi) infection in cervids. Parasitol. Res. 2009, 104, 919–925. [Google Scholar] [CrossRef]
- Kynkäänniemi, S.M.; Kettu, M.; Kortet, R.; Härkönen, L.; Kaitala, A.; Paakkonen, T.; Mustonen, A.M.; Nieminen, P.; Härkönen, S.; Ylönen, H.; et al. Acute impacts of the deer ked (Lipoptena cervi) infestation on reender (Rangifer tarandus tarandus) behaviour. Parasitol. Res. 2014, 113, 1489–1497. [Google Scholar] [CrossRef]
- Dehio, C.; Sauder, U.; Hiestand, R. Isolation of Bartonella schoenbuchensis from Lipoptena cervi, a blood-sucking arthropod causing deer ked dermatitis. J. Clin. Microbiol. 2004, 42, 5320–5323. [Google Scholar] [CrossRef]
- Duodu, S.; Madslien, K.; Hjelm, E.; Molin, Y.; Paziewska-Harris, A.; Harris, P.D.; Colquhoun, D.J.; Ytrehus, B. Bartonella infections in deer keds (Lipoptena cervi) and moose (Alces alces) in Norway. Appl. Environ. Microbiol. 2013, 79, 322–327. [Google Scholar] [CrossRef]
- Lee, S.H.; Kim, K.T.; Kwon, O.D.; Ock, Y.; Kim, T.; Choi, D.; Kwak, D. Novel detection of Coxiella spp., Theileria luwenshuni, and T. ovis endosymbionts in deer keds (Lipoptena fortisetosa). PLoS ONE 2016, 11, e0156727. [Google Scholar] [CrossRef]
- Szewczyk, T.; Werszko, J.; Steiner-Bogdaszewska, Ż.; Jeżewski, W.; Laskowski, Z.; Karbowiak, G. Molecular detection of Bartonella spp. in deer ked (Lipoptena cervi) in Poland. Parasit. Vectors 2017, 10, 487. [Google Scholar] [CrossRef] [PubMed]
- Werszko, J.; Steiner-Bogdaszewska, Ż.; Jeżewski, W.; Szewczyk, T.; Kuryło, G.; Wołkowycki, M.; Wróblewski, P.; Karbowiak, G. Molecular detection of Trypanosoma spp. in Lipoptena cervi and Lipoptena fortisetosa (Diptera: Hippoboscidae) and their potential role in the transmission of pathogens. Parasitology 2020, 147, 1629–1635. [Google Scholar] [CrossRef] [PubMed]
- Gałęcki, R.; Jaroszewski, J.; Bakuła, T.; Galon, E.M.; Xuan, X. Molecular detection of selected pathogens with zoonotic potential in deer keds (Lipoptena fortisetosa). Pathogens 2021, 10, 324. [Google Scholar] [CrossRef] [PubMed]
- Sato, S.; Kabeya, H.; Ishiguro, S.; Shibasaki, Y.; Maruyama, S. Lipoptena fortisetosa as a vector of Bartonella bacteria in Japanese sika deer (Cervus nippon). Parasite. Vector. 2021, 14, 73. [Google Scholar] [CrossRef]
- Andreani, A.; Sacchetti, P.; Belcari, A. Comparative morphology of the deer ked Lipoptena fortisetosa first recorded from Italy. Med. Vet. Entomol. 2019, 33, 140–153. [Google Scholar] [CrossRef]
- Andreani, A.; Sacchetti, P.; Belcari, A. Evolutionary adaptations in four hippoboscid fly species belonging to three different subfamilies. Med. Vet. Entomol. 2020, 34, 344–363. [Google Scholar] [CrossRef]
- Maa, T.C.; Peterson, B.V. Hippoboscidae. In Manual of Nearctic Diptera; McAlpine, J.F., Peterson, B.V., Shewell, G.E., Teskey, H.J., Vockeroth, J.R., Wood, D.M., Eds.; Research Branch, Agriculture Canada: Ottawa, ON, Canada, 1987; Volume 2, pp. 1271–1281. [Google Scholar]
- Zhang, D.; Liu, X.H.; Li, X.Y.; Cao, J.; Chu, H.J.; Li, K. Ultrastructural investigation of antennae in three cutaneous myiasis flies: Melophagus ovinus, Hippobosca equina, and Hippobosca longipennis (Diptera: Hippoboscidae). Parasitol. Res. 2015, 114, 1887–1896. [Google Scholar] [CrossRef]
- Gibson, G.; Torr, S.J. Visual and olfactory responses of haematophagous Diptera to host stimuli. Med. Vet. Entomol. 1999, 13, 2–23. [Google Scholar] [CrossRef]
- Cardé, R.T.; Gibson, G. Host finding by female mosquitoes: Mechanisms of orientation to host odours and other cues. In Olfaction in Vector-Host Interactions; Takken, W., Knols, B.G.J., Eds.; Wageningen Academic Publishers: Wageningen, The Netherlands, 2010; Volume 2, pp. 115–141. [Google Scholar] [CrossRef]
- Härkönen, L.; Härkönen, S.; Kaitala, A.; Kaunisto, S.; Kortet, R.; Laaksonen, S.; Ylönen, H. Predicting range expansion of an ectoparasite–the effect of spring and summer temperatures on deer ked Lipoptena cervi (Diptera: Hippoboscidae) performance along a latitudinal gradient. Ecography 2010, 33, 906–912. [Google Scholar] [CrossRef]
- Hennig, W. Die Verwandtschaftsbeziehungen der Pupiparen und die Morphologie der Sternalregion des Thorax der Dipteren. Arb. Morph. Taxon. Ent. Berl. 1941, 8, 231–2491. Available online: http://sdei.senckenberg.de/ (accessed on 4 January 2022).
- Peterson, B.V.; Wenzel, R.L. Nycteribiidae. In Manual of Nearctic Diptera; McAlpine, J.F., Peterson, B.V., Shewell, G.E., Teskey, H.J., Vockeroth, J.R., Wood, D.M., Eds.; Research Branch, Agriculture Canada: Ottawa, ON, Canada, 1987; Volume 2, pp. 1283–1291. [Google Scholar]
- Wenzel, R.L.; Peterson, B.V. Streblidae. In Manual of Nearctic Diptera; McAlpine, J.F., Peterson, B.V., Shewell, G.E., Teskey, H.J., Vockeroth, J.R., Wood, D.M., Eds.; Research Branch, Agriculture Canada: Ottawa, ON, Canada, 1987; Volume 2, pp. 1293–1301. [Google Scholar]
- Jobling, B. The structure of the head and mouth-parts in the Nycteribiidae (Diptera Pupipara). Parasitology 1928, 20, 254–272. [Google Scholar] [CrossRef]
- Jobling, B. A comparative study of the structure of the head and mouth parts in the Streblidae (Diptera Pupipara). Parasitology 1929, 21, 417–445. [Google Scholar] [CrossRef]
- McAlpine, J.F.; Peterson, B.V.; Shewell, G.E.; Teskey, H.J.; Vockeroth, J.R.; Wood, D.M. Manual of Nearctic Diptera, Volume 1, Monograph 27; Research Branch, Agriculture Canada: Ottawa, ON, Canada, 1981; pp. 1–674. [Google Scholar]
- Krinsky, W.L. Tsetse flies (Glossinidae). In Medical and Veterinary Entomology, 3rd ed.; Mullen, G.R., Durden, L.A., Eds.; Elsevier: London, UK, 2019; pp. 369–382. [Google Scholar] [CrossRef]
- Isaac, C.; Ravaiano, S.V.; Vicari Pascini, T.; Ferreira Martins, G. The antennal sensilla of species of the Palpalis group (Diptera: Glossinidae). J. Med. Entomol. 2015, 52, 614–621. [Google Scholar] [CrossRef] [PubMed]
- White, S.L.; Bay, D.E. Antennal olfactory sensilla of the horn fly, Haematobia irritans irritans (L.) (Diptera: Muscidae). J. Kans. Entomol. Soc. 1980, 53, 641–652. Available online: https://www.jstor.org/stable/25084087 (accessed on 4 January 2022).
- Been, T.H.; Schomaker, C.H.; Thomas, G. Olfactory sensilla on the antenna and maxillary palp of the sheep head fly, Hydrotaea irritans (Fallen) (Diptera: Muscidae). Int. J. Insect Morphol. 1988, 17, 121–133. [Google Scholar] [CrossRef]
- Pezzi, M.; Scapoli, C.; Mamolini, E.; Leis, M.; Bonacci, T.; Whitmore, D.; Krčmar, S.; Furini, M.; Giannerini, S.; Chicca, M.; et al. Ultrastructural characterization of sensilla and microtrichia on the antenna of female Haematopota pandazisi (Diptera: Tabanidae). Parasitol. Res. 2018, 117, 959–970. [Google Scholar] [CrossRef]
- Chapman, R.F. Chemoreception: The significance of receptor numbers. Adv. Insect Physiol. 1982, 16, 247–356. [Google Scholar] [CrossRef]
- Lehane, M.J. The Biology of Blood-Sucking in Insects, 2nd ed.; Cambridge University Press: Cambridge, UK, 2005; pp. 1–321. [Google Scholar] [CrossRef]
- Chahda, J.S.; Soni, N.; Sun, J.S.; Ebrahim, S.A.M.; Weiss, B.L.; Carlson, J.R. The molecular and cellular basis of olfactory response to tsetse fly attractants. PLoS Genet. 2019, 15, e1008005. [Google Scholar] [CrossRef]
- Clyne, P.; Grant, A.; O’Connell, R.; Carlson, J.R. Odorant response of individual sensilla on the Drosophila antenna. Invertebr. Neurosci. 1997, 3, 127–135. [Google Scholar] [CrossRef]
- Kurtovic, A.; Widmer, A.; Dickson, B.J. A single class of olfactory neurons mediates behavioural responses to a Drosophila sex pheromone. Nature 2007, 446, 542–546. [Google Scholar] [CrossRef]
- Yao, C.A.; Ignell, R.; Carlson, J.R. Chemosensory coding by neurons in the coeloconic sensilla of the Drosophila antenna. J. Neurosci. 2005, 25, 8359–8367. [Google Scholar] [CrossRef] [PubMed]
- Meijerink, J.; Braks, M.A.H.; Van Loon, J.J.A. Olfactory receptors on the antennae of the malaria mosquito Anopheles gambiae are sensitive to ammonia and other sweat-borne components. J. Insect Physiol. 2001, 47, 455–464. [Google Scholar] [CrossRef]
- Stange, G.; Stowe, S. Carbon-dioxide sensing structures in terrestrial arthropods. Microsc. Res. Tech. 1999, 47, 416–427. [Google Scholar] [CrossRef]
- Boo, K.S.; McIver, S.B. Fine structure of sunken thick-walled pegs (sensilla ampullacea and coeloconica) on the antennae of mosquitoes. Can. J. Zool. 1975, 53, 262–266. [Google Scholar] [CrossRef] [PubMed]
- Chu-Wang, I.W.; Axtell, R.C.; Kline, D.L. Antennal and palpal sensilla of the sand fly Culicoides furens (Poey) (Diptera: Ceratopogonidae). Int. J. Insect Morphol. 1975, 4, 131–149. [Google Scholar] [CrossRef]
- Kortet, R.; Härkönen, L.; Hokkanen, P.; Härkönen, S.; Kaitala, A.; Kaunisto, S.; Laaksonen, S.; Kekäläinen, J.; Ylönen, H. Experiments on the ectoparasitic deer ked that often attacks humans; preferences for body parts, colour and temperature. Bull. Entomol. Res. 2010, 100, 279–285. [Google Scholar] [CrossRef]
- Lourenço, S.I.; Palmeirim, J.M. How do ectoparasitic nycteribiids locate their bat hosts? Parasitology 2008, 135, 1205–1213. [Google Scholar] [CrossRef]
- Tangtrakulwanich, K.; Chen, H.; Baxendale, F.; Brewer, G.; Zhu, J.J. Characterization of olfactory sensilla of Stomoxys calcitrans and electrophysiological responses to odorant compounds associated with hosts and oviposition media. Med. Vet. Entomol. 2011, 25, 327–336. [Google Scholar] [CrossRef]
- Shanbhag, S.R.; Müller, B.; Steinbrecht, R.A. Atlas of olfactory organs of Drosophila melanogaster. 2. Internal organization and cellular architecture of olfactory sensilla. Arthropod Struct. Dev. 2000, 29, 211–229. [Google Scholar] [CrossRef]
- De Rose, F.; Corda, V.; Solari, P.; Sacchetti, P.; Belcari, A.; Poddighe, S.; Kasture, S.; Solla, P.; Marrosu, F.; Liscia, A. Drosophila mutant model of Parkinson’s disease revealed an unexpected olfactory performance: Morphofunctional evidences. Parkinson’s Dis. 2016, 2016, 3508073. [Google Scholar] [CrossRef]
- Setzu, M.D.; Poddighe, S.; Angioy, A.M. Sensilla on the antennal funiculus of the blow fly, Protophormia terraenovae (Diptera: Calliphoridae). Micron 2011, 42, 471–477. [Google Scholar] [CrossRef] [PubMed]
- Sukontason, K.; Sukontason, K.L.; Piangjai, S.; Boonchu, N.; Chaiwong, T.; Ngern-klun, R.; Sripakdeea, D.; Vogtsbergerb, R.C.; Olson, J.K. Antennal sensilla of some forensically important flies in families Calliphoridae, Sarcophagidae and Muscidae. Micron 2004, 35, 671–679. [Google Scholar] [CrossRef] [PubMed]
- Hu, F.; Zhang, G.N.; Jia, F.X.; Dou, W.; Wang, J.J. Morphological characterization and distribution of antennal sensilla of six fruit flies (Diptera: Tephritidae). Ann. Entomol. Soc. Am. 2010, 103, 661–670. [Google Scholar] [CrossRef]
- Liscia, A.; Angioni, P.; Sacchetti, P.; Poddighe, S.; Granchietti, A.; Setzu, M.D.; Belcari, A. Characterization of olfactory sensilla of the olive fly: Behavioral and electrophysiological responses to volatile organic compounds from the host plant and bacterial filtrate. J. Insect Physiol. 2013, 59, 705–716. [Google Scholar] [CrossRef] [PubMed]
- Zacharuk, R.Y. Ultrastructure and function of insect chemosensilla. Annu. Rev. Entomol. 1980, 25, 27–47. [Google Scholar] [CrossRef]
- Jeong, S.A.; Kim, J.; Byun, B.K.; Oh, H.W.; Park, K.C. Morphological and ultrastructural characterization of olfactory sensilla in Drosophila suzukii: Scanning and transmission electron microscopy. J. Asia-Pac. Entomol. 2020, 23, 1165–1180. [Google Scholar] [CrossRef]
- Schneider, D.; Steinbrecht, R.A. Checklist of insect olfactory sensilla. In Invertebrate Receptors; Carthy, J.D., Newell, G.E., Eds.; Symposia of the Zoological Society London, Academic Press: London, UK, 1968; Volume 23, pp. 279–297. [Google Scholar]
- Diehl, P.A.; Vlimant, M.; Guerenstein, P.; Guerin, P.M. Ultrastructure and receptor cell responses of the antennal grooved peg sensilla of Triatoma infestans (Hemiptera: Reduviidae). Arthropod Struct. Dev. 2003, 31, 271–285. [Google Scholar] [CrossRef]
- Romani, R.; Stacconi, M.V.R.; Riolo, P.; Isidoro, N. The sensory structures of the antennal flagellum in Hyalesthes obsoletus (Hemiptera: Fulgoromorpha: Cixiidae): A functional reduction? Arthropod Struct. Dev. 2009, 38, 473–483. [Google Scholar] [CrossRef]
- Shanbhag, S.R.; Müller, B.; Steinbrecht, R.A. Atlas of olfactory organs of Drosophila melanogaster. 1. Types, external organization, innervation and distribution of olfactory sensilla. Int. J. Insect Morphol. 1999, 28, 377–397. [Google Scholar] [CrossRef]
- de Bruyne, M.; Foster, K.; Carlson, J.R. Odor coding in the Drosophila antenna. Neuron 2001, 30, 537–552. [Google Scholar] [CrossRef]
- Fernandes, F.D.F.; Linardi, P.M.; Chiarini-Garcia, H. Morphology of the antenna of Dermatobia hominis (Diptera: Cuterebridae) based on scanning electron microscopy. J. Med. Entomol. 2002, 39, 36–43. [Google Scholar] [CrossRef] [PubMed]
- Stoffolano, J.G., Jr.; Rice, M.; Murphy, W.L. The importance of antennal mechanosensilla of Sepedon fuscipennis (Diptera: Sciomyzidae). Can. Entomol. 2013, 145, 265–272. [Google Scholar] [CrossRef]
- Foelix, R.F.; Stocker, R.F.; Steinbrecht, R.A. Fine structure of a sensory organ in the arista of Drosophila melanogaster and some other dipterans. Cell Tissue Res. 1989, 258, 277–287. [Google Scholar] [CrossRef] [PubMed]
- Andreani, A.; Rosi, M.C.; Guidi, R.; Jafrancesco, D.; Farini, A.; Belcari, A.; Sacchetti, P. Colour Preference of the Deer Ked Lipoptena fortisetosa (Diptera: Hippoboscidae). Insects 2021, 12, 845. [Google Scholar] [CrossRef] [PubMed]












Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Andreani, A.; Belcari, A.; Sacchetti, P.; Romani, R. Antennal Morphology and Fine Structure of Flagellar Sensilla in Hippoboscid Flies with Special Reference to Lipoptena fortisetosa (Diptera: Hippoboscidae). Insects 2022, 13, 236. https://doi.org/10.3390/insects13030236
Andreani A, Belcari A, Sacchetti P, Romani R. Antennal Morphology and Fine Structure of Flagellar Sensilla in Hippoboscid Flies with Special Reference to Lipoptena fortisetosa (Diptera: Hippoboscidae). Insects. 2022; 13(3):236. https://doi.org/10.3390/insects13030236
Chicago/Turabian StyleAndreani, Annalisa, Antonio Belcari, Patrizia Sacchetti, and Roberto Romani. 2022. "Antennal Morphology and Fine Structure of Flagellar Sensilla in Hippoboscid Flies with Special Reference to Lipoptena fortisetosa (Diptera: Hippoboscidae)" Insects 13, no. 3: 236. https://doi.org/10.3390/insects13030236
APA StyleAndreani, A., Belcari, A., Sacchetti, P., & Romani, R. (2022). Antennal Morphology and Fine Structure of Flagellar Sensilla in Hippoboscid Flies with Special Reference to Lipoptena fortisetosa (Diptera: Hippoboscidae). Insects, 13(3), 236. https://doi.org/10.3390/insects13030236