
The Supercooling Responses of the Solitary Bee Osmia excavata (Hymenoptera: Megachilidae) under the Biological Stress of Its Brood Parasite, Sapyga coma (Hymenoptera: Sapygidae)


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Insects
2.2. Measurement of Supercooling Points in Our Test Insects
2.3. Measurement of Body Weight, Water Content, and Fat Content
2.4. Statistical Analyses
3. Results
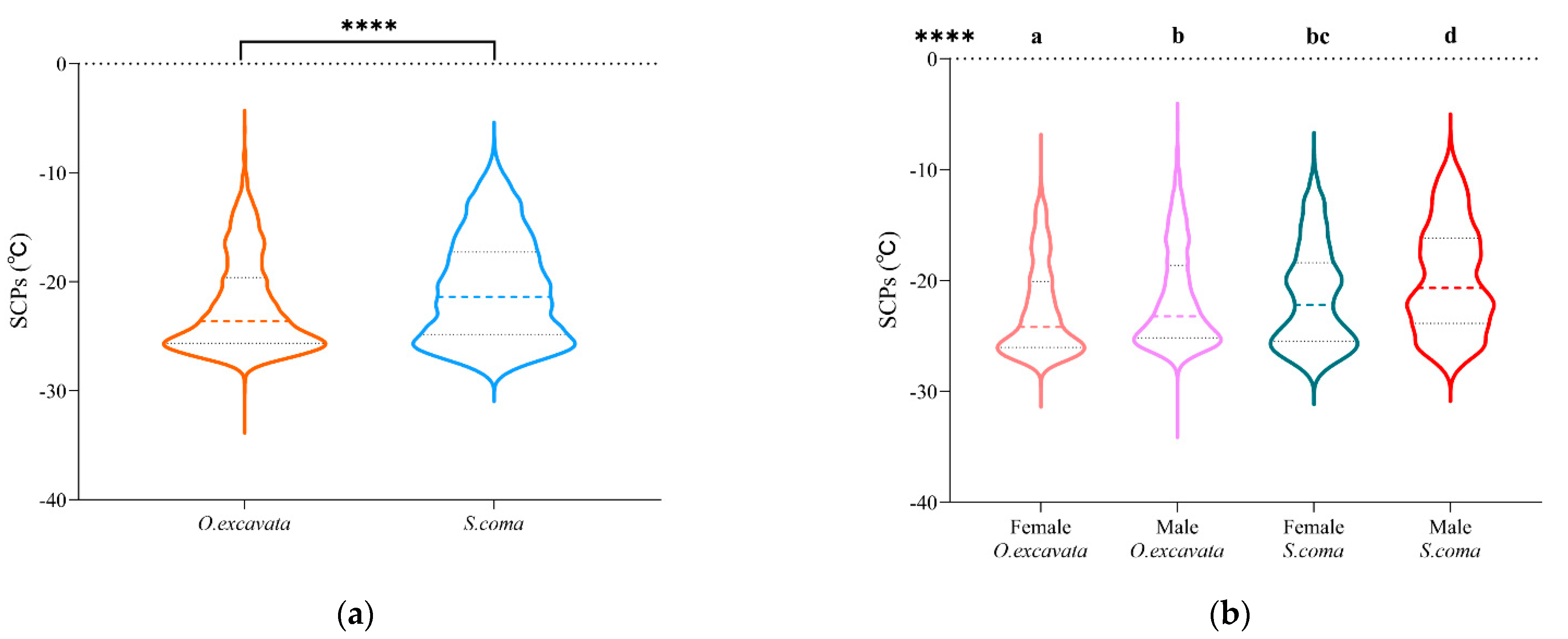
3.1. The Supercooling Points of O. excavata and S. coma
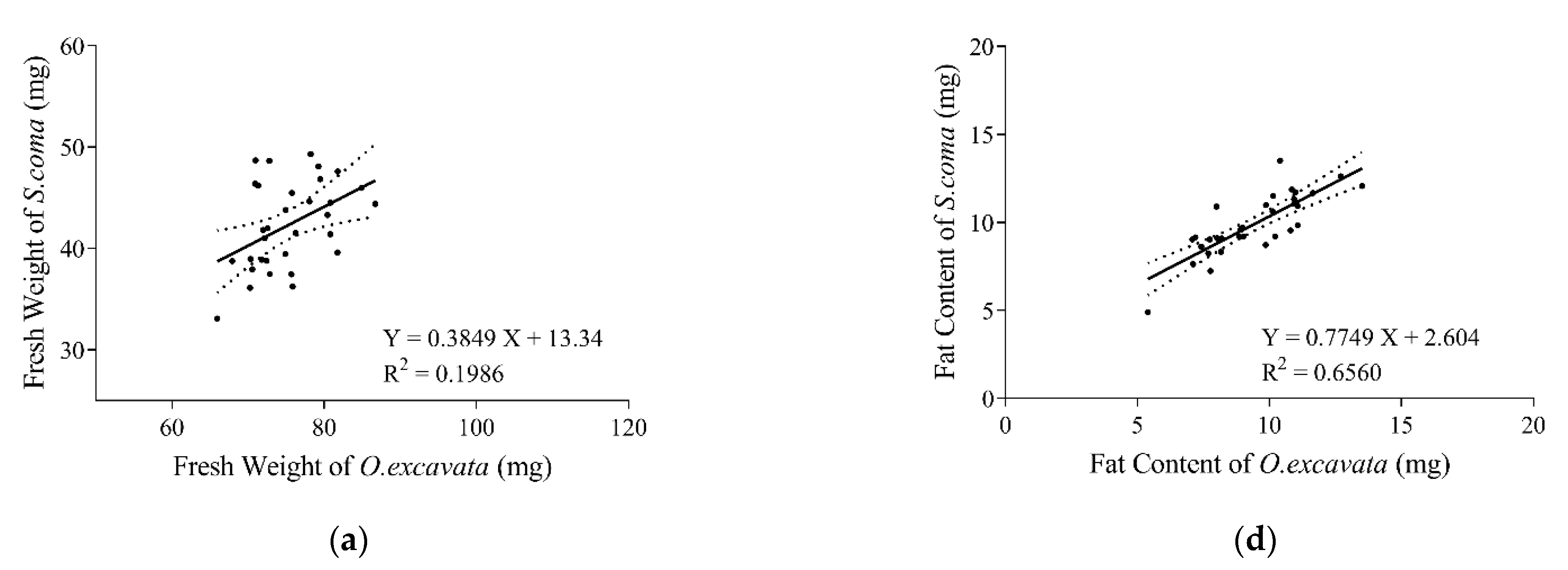
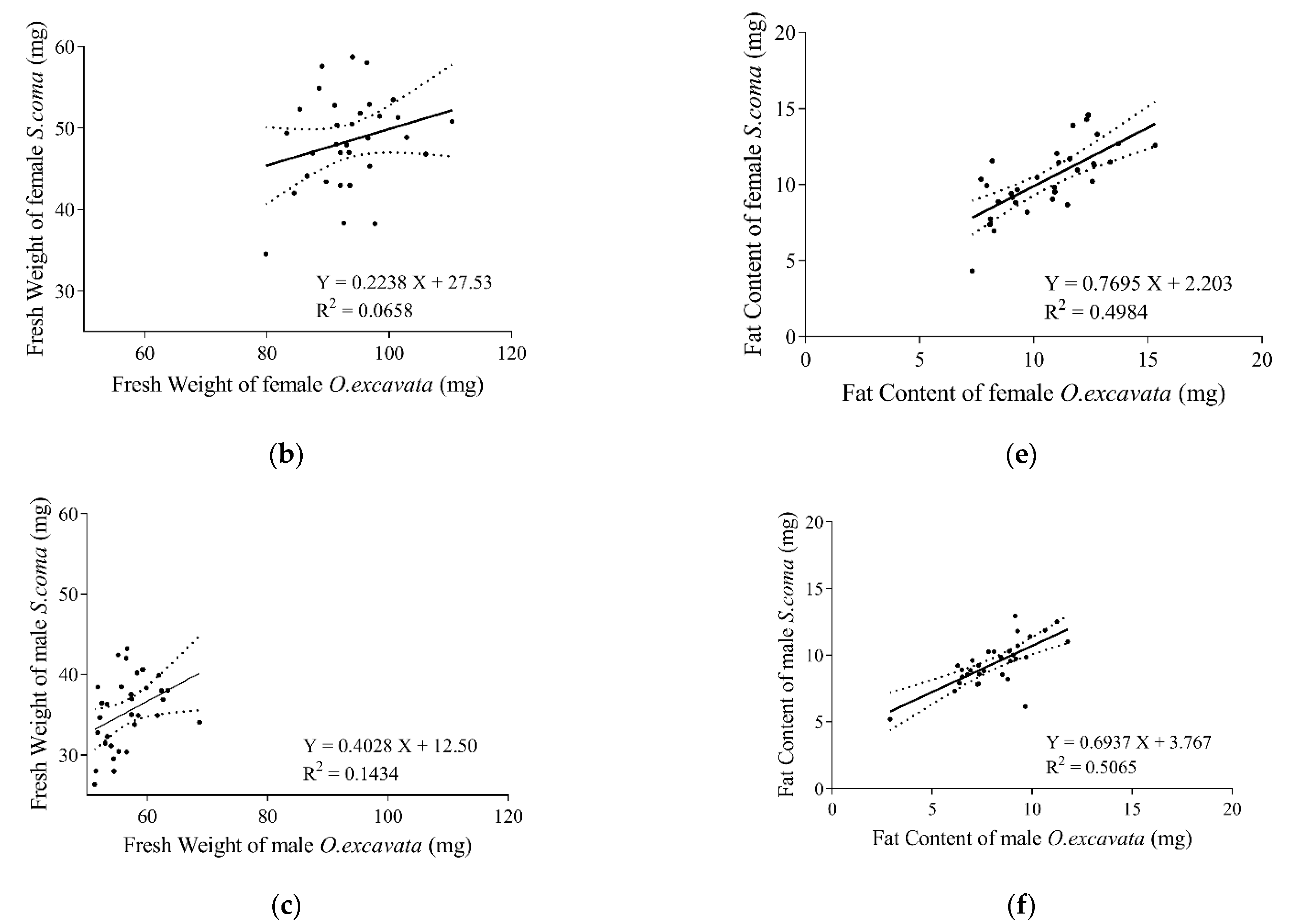
3.2. Relationship of Supercooling Points between O. excavata and S. coma
3.3. Physiological Parameters of the Solitary Bee and Its Brood Parasite
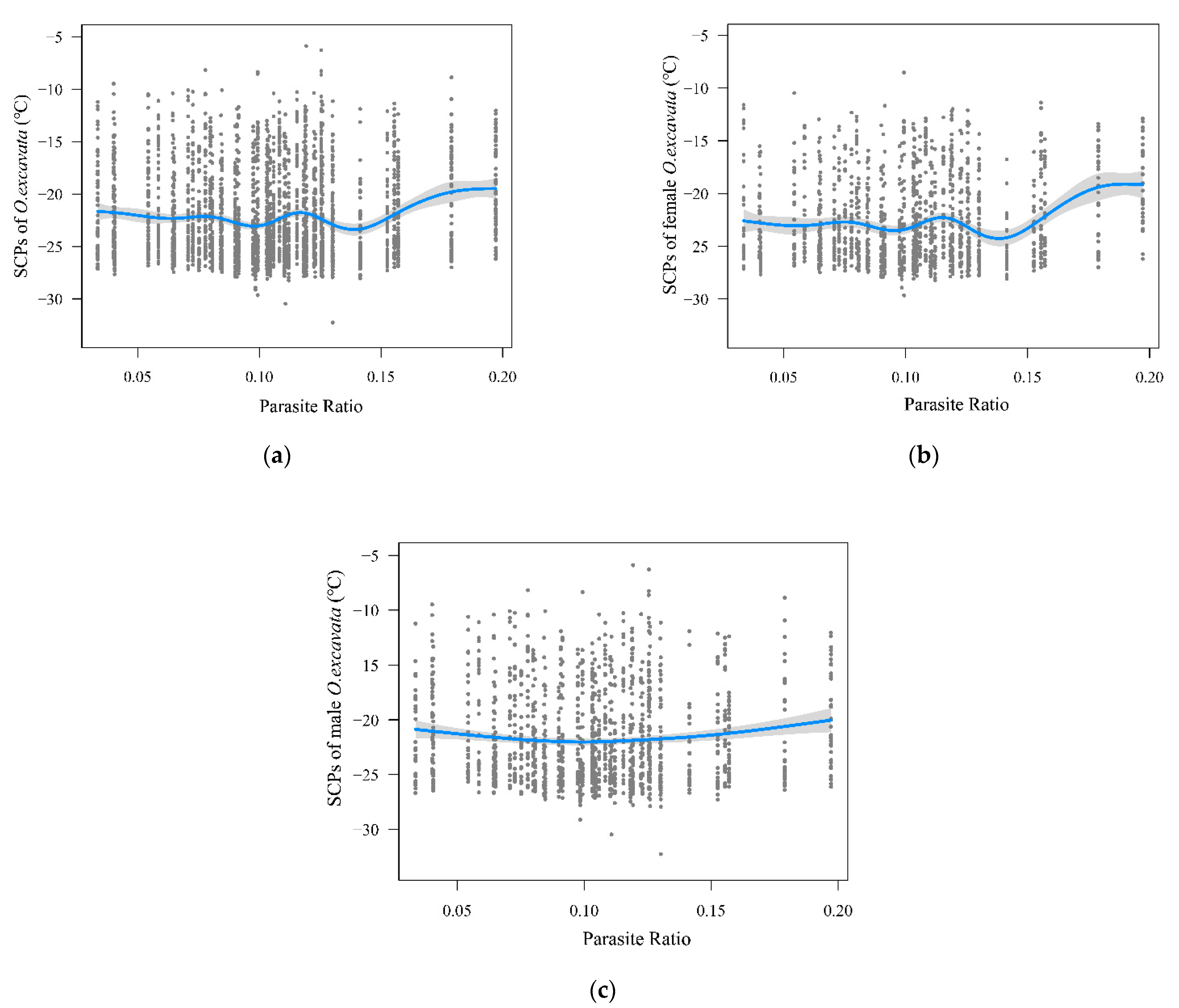
3.4. Relationship between Parasite Ratio and Supercooling Points of O. excavata
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lee, R.E.; Costanzo, J.P. Biological ice nucleation and ice distribution in cold-hardy ectothermic animals. Annu. Rev. Physiol. 1998, 60, 55–72. [Google Scholar] [CrossRef]
- Duman, J.G.; Wu, D.W.; Xu, L.; Tursman, D.; Olsen, T.M. Adaptations of Insects to Subzero Temperatures. Q. Rev. Biol. 1991, 66, 387–410. [Google Scholar] [CrossRef]
- Van Damme, V.; Berkvens, N.; Moerkens, R.; Berckmoes, E.; Wittemans, L.; De Vis, R.; Casteels, H.; Tirry, L.; De Clercq, P. Overwintering potential of the invasive leafminer Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae) as a pest in greenhouse tomato production in Western Europe. J. Pest Sci. 2015, 88, 533–541. [Google Scholar] [CrossRef]
- Costanzo, J.P.; Iverson, J.B.; Wright, M.F.; Lee, R.E. Cold Hardiness and Overwintering Strategies of Hatchlings in an Assemblage of Northern Turtles. Ecology 1995, 76, 1772–1785. [Google Scholar] [CrossRef]
- Costanzo, J.P.; Lee, R.E.; Ultsch, G.R. Physiological ecology of overwintering in hatchling turtles. J. Exp. Zool. Part A Ecol. Integr. Physiol. 2008, 309, 297–379. [Google Scholar] [CrossRef]
- Nicolai, A.; Vernon, P.; Lee, M.; Ansart, A.; Charrier, M. Supercooling ability in two populations of the land snail Helix pomatia (Gastropoda: Helicidae) and ice-nucleating activity of gut bacteria. Cryobiology 2005, 50, 48–57. [Google Scholar] [CrossRef]
- Sømme, L. Supercooling and winter survival in terrestrial arthropods. Comp. Biochem. Physiol. Part A Physiol. 1982, 73, 519–543. [Google Scholar] [CrossRef]
- Ouyang, F.; Liu, Z.; Yin, J.; Su, J.; Wang, C.; Ge, F. Effects of transgenic Bt cotton on overwintering characteristics and survival of Helicoverpa armigera. J. Insect Physiol. 2011, 57, 153–160. [Google Scholar] [CrossRef]
- Jakobs, R.; Gariepy, T.D.; Sinclair, B.J. Adult plasticity of cold tolerance in a continental-temperate population of Drosophila suzukii. J. Insect Physiol. 2015, 79, 1–9. [Google Scholar] [CrossRef]
- Vesala, L.; Hoikkala, A. Effects of photoperiodically induced reproductive diapause and cold hardening on the cold tolerance of Drosophila montana. J. Insect Physiol. 2011, 57, 46–51. [Google Scholar] [CrossRef]
- Wang, H.-S.; Zhou, C.-S.; Guo, W.; Kang, L. Thermoperiodic acclimations enhance cold hardiness of the eggs of the migratory locust. Cryobiology 2006, 53, 206–217. [Google Scholar] [CrossRef]
- Miller, K. Cold-hardiness strategies of some adult and immature insects overwintering in interior Alaska. Comp. Biochem. Physiol. Part A Physiol. 1982, 73, 595–604. [Google Scholar] [CrossRef]
- Ring, R.A. Freezing-tolerant insects with low supercooling points. Comp. Biochem. Physiol. Part A Physiol. 1982, 73, 605–612. [Google Scholar] [CrossRef]
- Lee, R.E.; Chen, C.-P.; Denlinger, D.L. A Rapid Cold-Hardening Process in Insects. Science 1987, 238, 1415–1417. [Google Scholar] [CrossRef]
- Forrest, J.R.K.; Chisholm, S.P.M. Direct benefits and indirect costs of warm temperatures for high-elevation populations of a solitary bee. Ecology 2017, 98, 359–369. [Google Scholar] [CrossRef]
- Zaragoza-Trello, C.; Vilà, M.; Bartomeus, I. Interaction between warming and landscape foraging resource availability on solitary bee reproduction. J. Anim. Ecol. 2021, 90, 2536–2546. [Google Scholar] [CrossRef]
- Marais, E.; Chown, S.L. Beneficial acclimation and the Bogert effect. Ecol. Lett. 2008, 11, 1027–1036. [Google Scholar] [CrossRef]
- Schebeck, M.; Hansen, E.M.; Schopf, A.; Ragland, G.J.; Stauffer, C.; Bentz, B.J. Diapause and overwintering of two spruce bark beetle species. Physiol. Entomol. 2017, 42, 200–210. [Google Scholar] [CrossRef]
- De Majo, M.S.; Zanotti, G.; Campos, R.E.; Fischer, S. Effects of Constant and Fluctuating Low Temperatures on the Development of Aedes aegypti (Diptera: Culicidae) from a Temperate Region. J. Med. Entomol. 2019, 56, 1661–1668. [Google Scholar] [CrossRef]
- Singh, K.P.; Kochar, E.; Prasad, N.G. Egg Viability, Mating Frequency and Male Mating Ability Evolve in Populations of Drosophila melanogaster Selected for Resistance to Cold Shock. PLoS ONE 2015, 10, e0129992. [Google Scholar] [CrossRef]
- Barat, M.; Vernon, P.; Tarayre, M.; Atlan, A. Overwintering strategy of two weevils infesting three gorse species: When cold hardiness meets plant-insect interactions. J. Insect Physiol. 2010, 56, 170–177. [Google Scholar] [CrossRef]
- Torchio, P.F.; Bosch, J. Biology of Tricrania stansburyi, a Meloid Beetle Cleptoparasite of the Bee Osmia lignaria propinqua (Hymenoptera: Megachilidae). Ann. Entomol. Soc. Am. 1992, 85, 713–721. [Google Scholar] [CrossRef]
- Seidelmann, K. The function of the vestibulum in nests of a solitary stem-nesting bee, Osmia rufa (L.). Apidologie 1999, 30, 19–29. [Google Scholar] [CrossRef][Green Version]
- Münster-Swendsen, M.; Calabuig, I. Interaction between the solitary bee Chelostoma florisomne and its nest parasite Sapyga clavicornis—Empty cells reduce the impact of parasites. Ecol. Entomol. 2000, 25, 63–70. [Google Scholar] [CrossRef]
- Groulx, A.F.; Forrest, J.R.K. Nesting aggregation as a predictor of brood parasitism in mason bees (Osmia spp.). Ecol. Entomol. 2018, 43, 182–191. [Google Scholar] [CrossRef]
- Liu, L.; Li, L.; Li, C.; Yuan, R.; Yu, Y.; Men, X.; Ye, B. Population investigation and restriction factors analyses of Osmia excavata Alfken in Jiaodong. Apicult. China 2018, 69, 68–71. (In Chinese) [Google Scholar]
- Wei, S.-G.; Wang, R.; Smirle, M.J.; Xu, H.-L. Release of Osmia excavata and Osmia jacoti (Hymenoptera: Megachilidae) for apple pollination. Can. Entomol. 2002, 134, 369–380. [Google Scholar] [CrossRef]
- Lu, H.; Dou, F.; Hao, Y.; Li, Y.; Zhang, K.; Zhang, H.; Zhou, Z.; Zhu, C.; Huang, D.; Luo, A. Metabarcoding Analysis of Pollen Species Foraged by Osmia excavata Alfken (Hymenoptera: Megachilidae) in China. Front. Ecol. Evol. 2021, 9. [Google Scholar] [CrossRef]
- Strachecka, A.; Chobotow, J.; Paleolog, J.; Łoś, A.; Schulz, M.; Teper, D.; Kucharczyk, H.; Grzybek, M. Insights into the biochemical defence and methylation of the solitary bee Osmia rufa L.: A foundation for examining eusociality development. PLoS ONE 2017, 12, e0176539. [Google Scholar] [CrossRef] [PubMed]
- Schenk, M.; Mitesser, O.; Hovestadt, T.; Holzschuh, A. Overwintering temperature and body condition shift emergence dates of spring-emerging solitary bees. PeerJ 2018, 6, e4721. [Google Scholar] [CrossRef]
- Slominski, A.H.; Burkle, L.A. Solitary Bee Life History Traits and Sex Mediate Responses to Manipulated Seasonal Temperatures and Season Length. Front. Ecol. Evol. 2019, 7, 314. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Simpson, G.L. Modelling Palaeoecological Time Series Using Generalised Additive Models. Front. Ecol. Evol. 2018, 6, 149. [Google Scholar] [CrossRef]
- Krunić, M.; Stanisavljević, L. Supercooling points and diapause termination in overwintering adults of orchard bees Osmia cornuta and O. rufa (Hymenoptera: Megachilidae). Bull. Entomol. Res. 2006, 96, 323–326. [Google Scholar] [CrossRef]
- Liu, C.; Bu, H.D.; Feng, Z.L.; Gu, G.J.; Zhang, W.J.; Cao, Y.; Zhao, D.Y. Relationship between physiological indicators and cold resistance on fruit pollinators Osmia excavata Alfken. Heilongjiang Agric. Sci. 2016, 2, 72–76. [Google Scholar]
- Polidori, C.; Boesi, R.; Borsato, W. Few, small, and male: Multiple effects of reduced nest space on the offspring of the solitary wasp, Euodynerus (Pareuodynerus) posticus (Hymenoptera: Vespidae). Comptes Rendus Biol. 2011, 334, 50–60. [Google Scholar] [CrossRef]
- Hahn, D.A.; Denlinger, D.L. Meeting the energetic demands of insect diapause: Nutrient storage and utilization. J. Insect Physiol. 2007, 53, 760–773. [Google Scholar] [CrossRef] [PubMed]
- Slominski, A.H.; Burkle, L.A. Asynchrony between solitary bee emergence and flower availability reduces flower visitation rate and may affect offspring size. Basic Appl. Ecol. 2021, 56, 345–357. [Google Scholar] [CrossRef]
- Torchio, P.F. In-Nest Biologies and Development of Immature Stages of Three Osmia Species (Hymenoptera: Megachilidae). Ann. Entomol. Soc. Am. 1989, 82, 599–615. [Google Scholar] [CrossRef]
- Hahn, D.A.; Martin, A.R.; Porter, S.D. Body size, but not cooling rate, affects supercooling points in the red imported fire ant, Solenopsis invicta. Environ. Entomol. 2008, 37, 1074–1080. [Google Scholar] [CrossRef]
- Rozsypal, J.; Koštál, V.; Berková, P.; Zahradníčková, H.; Šimek, P. Seasonal changes in the composition of storage and membrane lipids in overwintering larvae of the codling moth, Cydia pomonella. J. Therm. Biol. 2014, 45, 124–133. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Wang, M.-L.; Ding, N.; Ma, W.-H.; Li, Y.-N.; Lei, C.-L.; Wang, X.-P. Relationships between body weight of overwintering larvae and supercooling capacity; diapause intensity and post-diapause reproductive potential in Chilo suppressalis Walker. J. Insect Physiol. 2011, 57, 653–659. [Google Scholar] [CrossRef] [PubMed]
- Seidelmann, K.; Ulbrich, K.; Mielenz, N. Conditional sex allocation in the Red Mason bee, Osmia rufa. Behav. Ecol. Sociobiol. 2010, 64, 337–347. [Google Scholar] [CrossRef]
- Tepedino, V.J.; McDonald, L.L.; Rothwell, R. Defense against parasitization in mud-nesting Hymenoptera: Can empty cells increase net reproductive output? Behav. Ecol. Sociobiol. 1979, 6, 99–104. [Google Scholar] [CrossRef]
- McKinney, M.I.; Park, Y.-L. Nesting Activity and Behavior of Osmia cornifrons (Hymenoptera: Megachilidae) Elucidated Using Videography. Psyche J. Entomol. 2012, 2012, 814097. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yan, Z.; Wang, L.; Reddy, G.V.P.; Gu, S.; Men, X.; Xiao, Y.; Su, J.; Ge, F.; Ouyang, F. The Supercooling Responses of the Solitary Bee Osmia excavata (Hymenoptera: Megachilidae) under the Biological Stress of Its Brood Parasite, Sapyga coma (Hymenoptera: Sapygidae). Insects 2022, 13, 235. https://doi.org/10.3390/insects13030235
Yan Z, Wang L, Reddy GVP, Gu S, Men X, Xiao Y, Su J, Ge F, Ouyang F. The Supercooling Responses of the Solitary Bee Osmia excavata (Hymenoptera: Megachilidae) under the Biological Stress of Its Brood Parasite, Sapyga coma (Hymenoptera: Sapygidae). Insects. 2022; 13(3):235. https://doi.org/10.3390/insects13030235
Chicago/Turabian StyleYan, Zhuo, Lina Wang, Gadi V. P. Reddy, Shimin Gu, Xingyuan Men, Yunli Xiao, Jianwei Su, Feng Ge, and Fang Ouyang. 2022. "The Supercooling Responses of the Solitary Bee Osmia excavata (Hymenoptera: Megachilidae) under the Biological Stress of Its Brood Parasite, Sapyga coma (Hymenoptera: Sapygidae)" Insects 13, no. 3: 235. https://doi.org/10.3390/insects13030235
APA StyleYan, Z., Wang, L., Reddy, G. V. P., Gu, S., Men, X., Xiao, Y., Su, J., Ge, F., & Ouyang, F. (2022). The Supercooling Responses of the Solitary Bee Osmia excavata (Hymenoptera: Megachilidae) under the Biological Stress of Its Brood Parasite, Sapyga coma (Hymenoptera: Sapygidae). Insects, 13(3), 235. https://doi.org/10.3390/insects13030235