
Effect of Feed Containing Hermetia illucens Larvae Immunized by Lactobacillus plantarum Injection on the Growth and Immunity of Rainbow Trout (Oncorhynchus mykiss)


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. HIL, Bacteria Strain, and Immunization
2.2. Preparation of HIL and ImHIL Extract
2.3. qRT-PCR for AMP Expression Analysis in ImHIL
2.4. RDA and MIC for the Analysis of the Antibacterial Activity of ImHIL
2.5. Formulated Feed Manufacture and Analysis of the Proximate Composition of the Feeds
2.6. Fish, Rearing Conditions, and Sampling
2.7. Analysis of Somatic and Hematological Indices
2.7.1. Red Blood Cell (RBC) Count
2.7.2. Packed Cell Volume (PCV)
2.7.3. Insulin-Like Growth Factor-1 (IGF-1)
2.8. Analysis of Immunity Enhancement
2.8.1. Aspartate Aminotransferase (AST)
2.8.2. Lysozyme Activity
2.8.3. Immunoglobulin M (IgM)
3. Results and Discussion
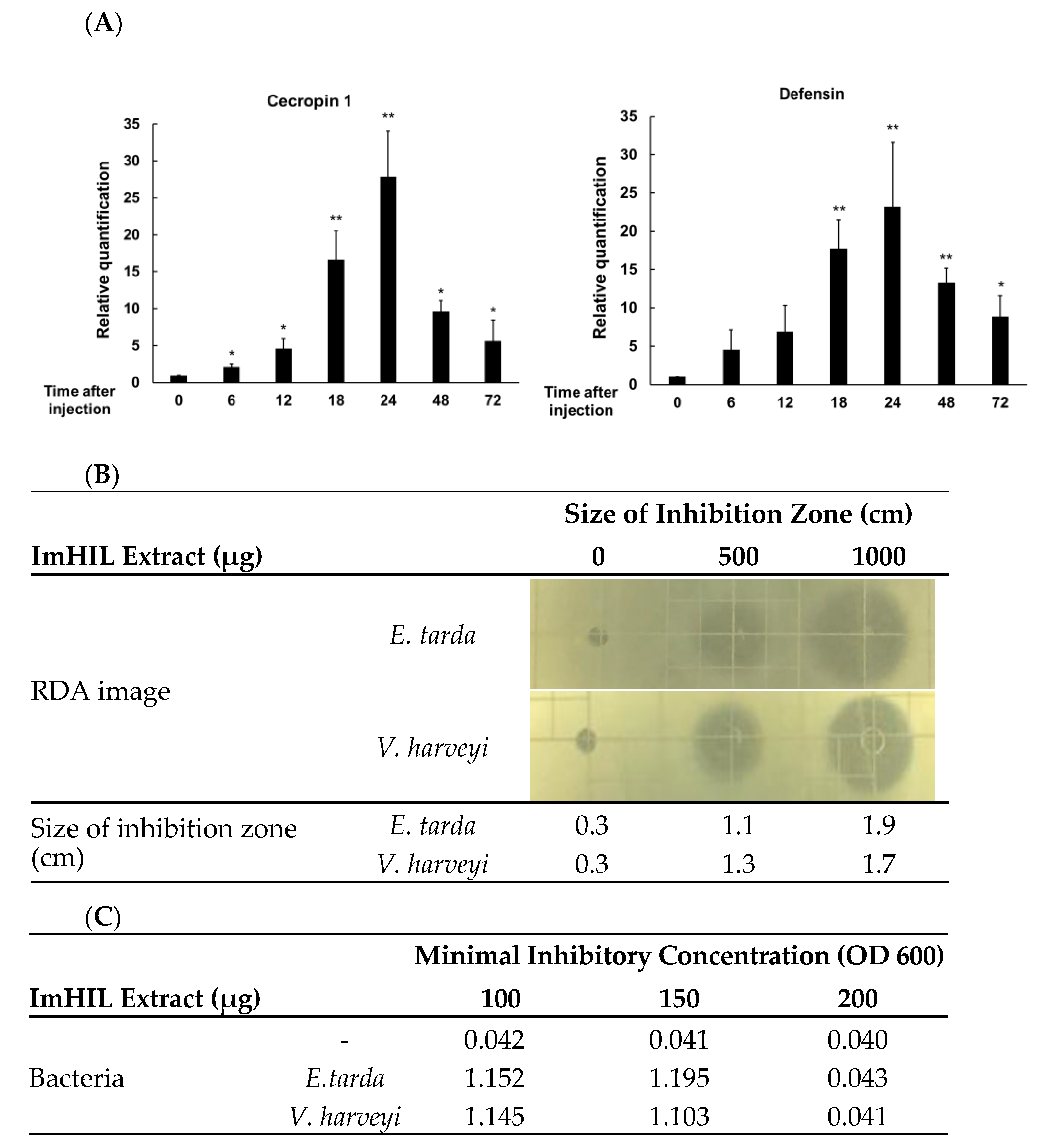
3.1. Establishment of Conditions for Inducing the Maximum Amount of AMPs in ImHIL
3.2. Ingredient of ImHIL-Containing Feed
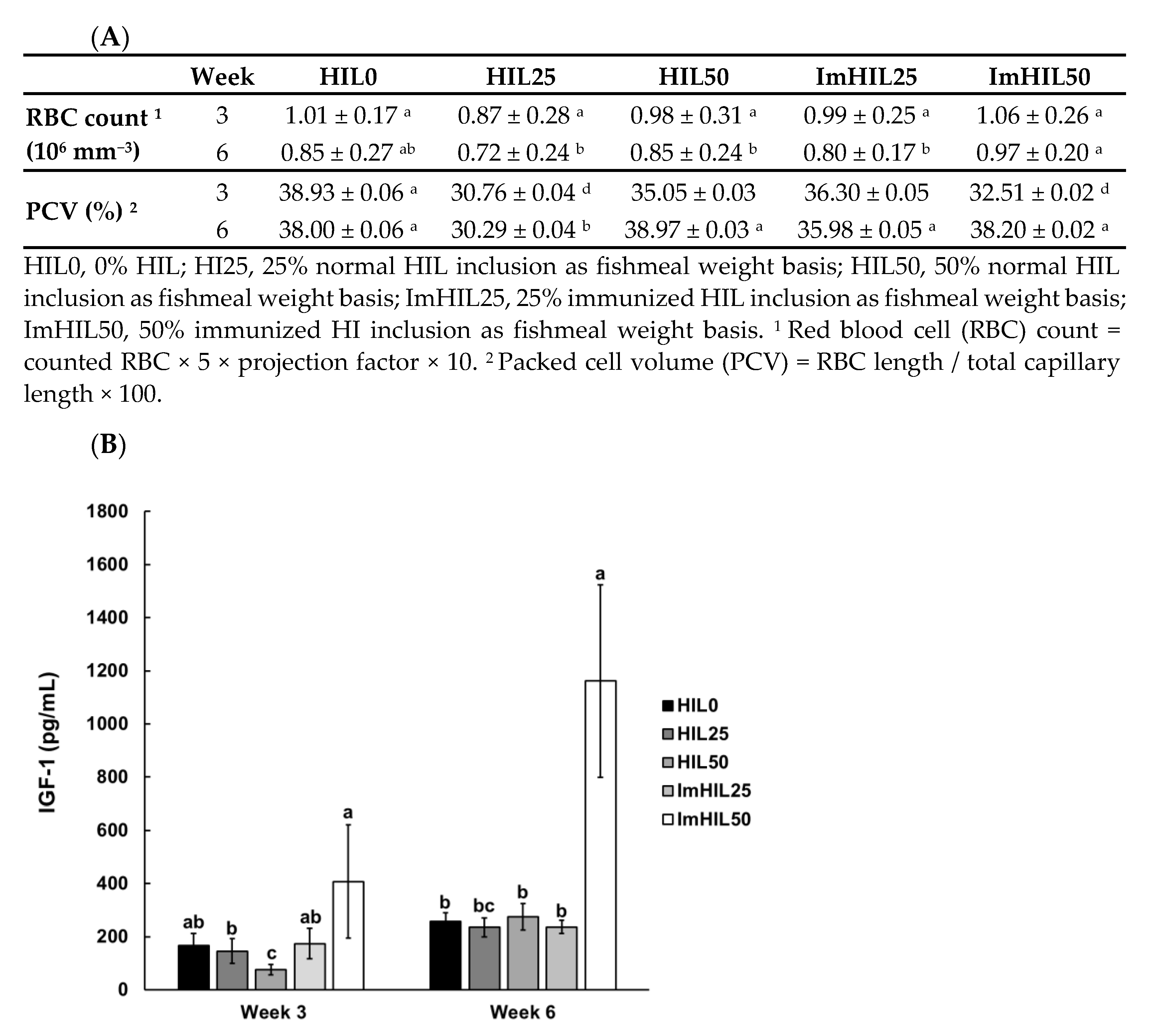
3.3. Effect of ImHIL Intake on the Somatic Index of O. mykiss
3.4. Effect of ImHIL Intake on the Hematological Index of O. mykiss
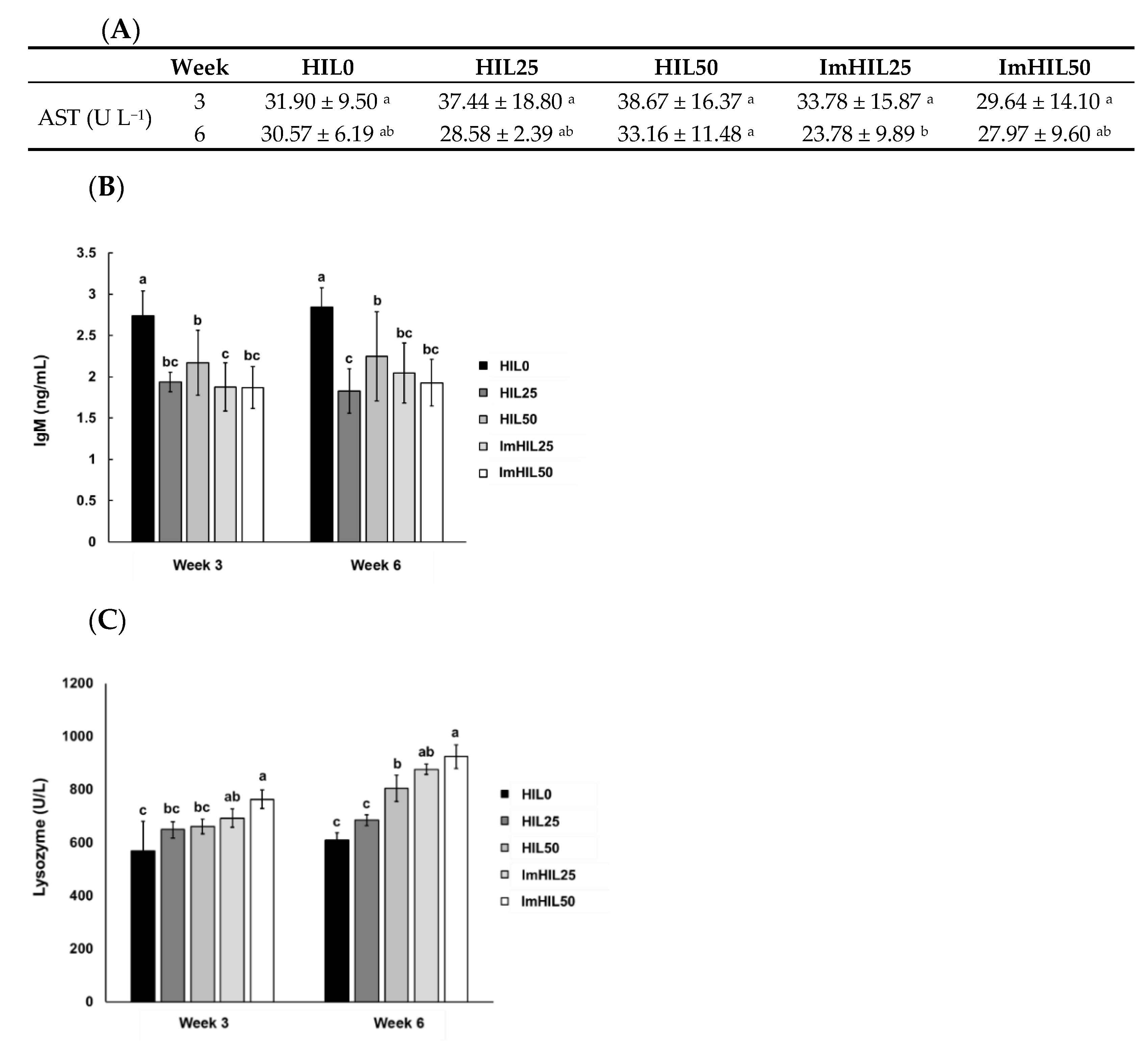
3.5. Effects of ImHIL Intake on Immunity Enhancement of O. mykiss
3.5.1. Analysis of AST in the Blood of O. mykiss Fed with ImHIL
3.5.2. Analysis of lysozyme Activity in the Blood of O. mykiss Fed with ImHIL
3.5.3. Analysis of IgM in the Blood of O. mykiss Fed with ImHIL
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sheeshka, J.; Murkin, E. Nutritional aspects of fish compared with other protein sources. Comm. Toxicol. 2002, 8, 375–397. [Google Scholar] [CrossRef]
- Stenberg, O.K.; Holen, E.; Piemontese, L.; Liland, N.S.; Lock, E.J.; Espe, M.; Belghit, I. Effect of dietary replacement of fish meal with insect meal on In vitro bacterial and viral induced gene response in atlantic salmon (Salmo salar) head kidney leukocytes. Fish Shellfish Immunol. 2019, 91, 223–232. [Google Scholar] [CrossRef] [PubMed]
- Daniel, D. A Review on replacing fish meal in aqua feeds using plant protein sources. Int. J. Fish. Aquat. 2018, 6, 164–179. [Google Scholar]
- Olsen, R.L.; Hasan, M.R. A limited supply of fish meal: Impact on future increases in global aquaculture production. Trends Food Sci. Technol. 2012, 27, 120–128. [Google Scholar] [CrossRef]
- Grau, T.; Vilcinskas, A.; Joop, G. Sustainable farming of the mealworm Tenebrio molitor for the production of food and feed. Z. Naturforsch. C J. Biosci. 2017, 72, 337–349. [Google Scholar] [CrossRef]
- Cashion, T.; Le Manach, F.L.; Zeller, D.; Pauly, D. Most fish destined for fish meal production are food-grade fish. Fish Fish. 2017, 18, 837–844. [Google Scholar] [CrossRef]
- Nogales-Mérida, S.; Gobbi, P.; Józefiak, D.; Mazurkiewicz, J.; Dudek, K.; Rawski, M.; Kierończyk, B.; Józefiak, A. Insect meals in fish nutrition. Rev. Aquacult. 2019, 11, 1080–1103. [Google Scholar] [CrossRef]
- Barragan-Fonseca, K.B.; Dicke, M.; van Loon, J.J.A. Nutritional value of the black soldier fly (Hermetia illucens L.) and its suitability as animal feed—A review. J. Insects Food Feed 2017, 3, 105–120. [Google Scholar] [CrossRef]
- Kim, T.K.; Yong, H.I.; Kim, Y.B.; Kim, H.W.; Choi, Y.S. Edible insects as a protein source: A review of public perception, processing technology, and research trends. Korean J. Food Sci. Food Sci. Anim. Resour. 2019, 39, 521–540. [Google Scholar] [CrossRef] [Green Version]
- Kroeckel, S.; Harjes, A.-G.E.; Roth, I.; Katz, H.; Wuertz, S.; Susenbeth, A.; Schulz, C. When a turbot catches a fly: Evaluation of a pre-pupae meal of the black soldier fly (Hermetia illucens) as fish meal substitute—Growth performance and chitin degradation in juvenile turbot (Psetta maxima). Aquaculture 2012, 364–365, 345–352. [Google Scholar] [CrossRef]
- Xiao, X.; Jin, P.; Zheng, L.; Cai, M.; Yu, Z.; Yu, J.; Zhang, J. Effects of black soldier fly (Hermetia illucens) larvae meal protein as a fish meal replacement on the growth and immune index of yellow catfish (Pelteobagrus fulvidraco). Aquac. Res. 2018, 49, 1569–1577. [Google Scholar] [CrossRef]
- Taufek, N.M.; Aspani, F.; Muin, H.; Raji, A.A.; Razak, S.A.; Alias, Z. The effect of dietary cricket meal (Gryllus bimaculatus) on growth performance, antioxidant enzyme activities, and haematological response of african catfish (Clarias Gariepinus). Fish Physiol. Biochem. 2016, 42, 1143–1155. [Google Scholar] [CrossRef] [PubMed]
- Widjastuti, T.; Wiradimadja, R.; Rusmana, D. The effect of substitution of fishmeal fish meal by black soldier fly (Hermetia illucens) maggot meal in the diet on production performance of quail (Coturnix japonica). Ani. Sci. 2014, 57, 125–129. [Google Scholar]
- Wang, Y.S.; Shelomi, M. Review of black soldier fly (Hermetia illucens) as animal feed and human food. Foods 2017, 6, 91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Devic, E.; Leschen, W.; Murray, F.; Little, D.C. Growth performance, feed utilization and body composition of advanced nursing nile tilapia (Oreochromis niloticus) fed diets containing black soldier fly (Hermetia illucens) larvae meal. Aquacult. Nutr. 2018, 24, 416–423. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Ji, H.; Yu, H.; Zhou, J. Influence of dietary black soldier fly (Hermetia illucens Linnaeus) pulp on growth performance, antioxidant capacity and intestinal health of juvenile mirror carp (Cyprinus carpio var. specularis). Aquacult. Nutr. 2020, 26, 432–443. [Google Scholar] [CrossRef]
- Borgogno, M.; Dinnella, C.; Iaconisi, V.; Fusi, R.; Scarpaleggia, C.; Schiavone, A.; Monteleone, E.; Gasco, L.; Parisi, G. Inclusion of Hermatia illucens larvae meal on rainbow trout (Oncorhynchus mykiss) feed: Effect sensory profile according to static and dynamic evaluations. J. Sci. Food Agric. 2017, 97, 3402–3411. [Google Scholar] [CrossRef]
- Dumas, A.; Raggi, T.; Barkhouse, J.; Lewis, E.; Weltzien, E. The oil fraction and partially defatted meal of black soldier fly larvae (Hermetia illucens) affect differently growth performance, feed efficiency, nutrient deposition, blood glucose and lipid digestibility of rainbow trout (Oncorhynchus mykiss). Aquaculture 2018, 492, 24–34. [Google Scholar] [CrossRef]
- Caimi, C.; Biasato, I.; Chemello, G.; Oddon, S.B.; Lussiana, C.; Malfatto, V.M.; Capucchio, M.T.; Colombino, E.; Schiavone, A.; Gai, F.; et al. Dietary inclusion of a partially defatted black soldier fly (Hermetia illucuns) larva meal in low fishmeal-based diets for rainbow trout (Onchrynchus mykiss). J. Anim. Sci. Technol. 2021, 12, 50. [Google Scholar]
- Stejskal, V.; Tran, H.Q.; Prokesova, M.; Gebauer, T.; Giang, P.T.; Gai, F.; Gasco, L. Partially defatted Hermetia illucens larva meal in diet of Eurasian perch (Perch fluviatilis) juveniles. Animals 2020, 10, 1876. [Google Scholar] [CrossRef]
- Abdel-Latif, H.M.R.; Abdel-Tawwab, M.; Khalil, R.H.; Metwally, A.A.; Shakweer, M.S.; Ghetas, H.A.; Khallaf, M.A. Black soldier fly (Hermetia illucens) larvae meal in diets of european seabass: Effects on antioxidantie capacity, non-specific immunity, transcriptomic responses, and resistance to the challenge with Vibrio alginolyticus. Fish Shellfish Immunol. 2021, 111, 111–118. [Google Scholar] [CrossRef]
- Yi, H.Y.; Chowdhury, M.; Huang, Y.D.; Yu, X.Q. Insect antimicrobial peptides and their applications. Appl. Microbiol. Biotechnol. 2014, 98, 5807–5822. [Google Scholar] [CrossRef] [Green Version]
- Shelomi, M.; Jacobs, C.; Vilcinskas, A.; Vogel, H. The unique antimicrobial peptide repertoire of stick insects. Dev. Comp. Immunol. 2020, 103, 103471. [Google Scholar] [CrossRef]
- Park, S.I.; Kim, J.W.; Yoe, S.M. Purification and characterization of a novel antibacterial peptide from black soldier fly (Hermetia illucens) larvae. Dev. Comp. Immunol. 2015, 52, 98–106. [Google Scholar] [CrossRef]
- Elhag, O.; Zhou, D.; Song, Q.; Soomro, A.A.; Cai, M.; Zheng, L.; Yu, Z.; Zhang, J. Screening, expression, purification and functional characterization of novel antimicrobial peptide genes from Hermetia illucens (L.). PLoS ONE 2017, 12, e0169582. [Google Scholar] [CrossRef]
- Bondari, K.; Sheppard, D.C. Soldier fly larvae as feed in commercial fish production. Aquaculture 1981, 24, 103–109. [Google Scholar] [CrossRef]
- Daneshmand, A.; Kermanshahi, H.; Sekhavati, M.H.; Javadmanesh, A.; Ahmadian, M. Antimicrobial peptide, cLF36, affects performance and intestinal morphology, microflora, junctional proteins, and immune cells in broilers challenged with E. coli. Sci. Rep. 2019, 9, 14176. [Google Scholar] [CrossRef]
- Mo, W.Y.; Lun, C.H.I.; Choi, W.M.; Man, Y.B.; Wong, M.H. Enhancing growth and non-specific immunity of grass carp and nile tilapia by incorporating chinese herbs (Astragalus membranaceus and Lycium barbarum) into food waste based pellets. Environ. Pollut. 2016, 219, 475–482. [Google Scholar] [CrossRef]
- Schiavone, A.; Cullere, A.; De Marco, M.; Meneguz, M.; Biasato, I.; Bergagna, A.; Dezzutto, D.; Gai, F.; Dabbou, S.; Gasco, L.; et al. Partial or total replacement of soybean oil by black soldier fly larvae (Hermetia illucens L.) fat in broiler diets: Effect on growth performances, feed-choice, blood traits, carcass characteristics and meat quality. Ital. J. Anim. Sci. 2016, 16, 93–100. [Google Scholar] [CrossRef] [Green Version]
- Vogel, H.; Müller, A.; Heckel, D.G.; Gutzeit, H.; Vilcinskas, A. nutritional immunology: Diversification and diet-dependent expression of antimicrobial peptides in the black soldier fly Hermetia illucens. Dev. Comp. Immunol. 2018, 78, 141–148. [Google Scholar] [CrossRef]
- Belforti, M.; Gai, F.; Lussiana, C.; Renna, M.; Malfatto, V.; Rotolo, L.; De Marco, M.; Dabbou, S.; Schiavone, A.; Zoccarato, I.; et al. Tenebrio molitor meal in rainbow trout (Oncorhynchus mykiss) diets: Effects on animal performance, nutrient digestibility and chemical composition of fillets. Ital. J. Anim. Sci. 2015, 14, 670–676. [Google Scholar] [CrossRef] [Green Version]
- AOAC. Official Methods of Analysis of AOAC International, 16th ed.; Association of Official Analytical Chemists: Gaithersburg, MA, USA, 2000. [Google Scholar]
- Ido, A.; Hashizume, A.; Ohta, T.; Takahashi, T.; Miura, C.; Miura, T. Replacement of fish meal by defatted yellow mealworm (Tenebrio molitor) larvae in diet improves growth performance and disease resistance in red seabream (Pargus major). Animals 2019, 9, 100. [Google Scholar] [CrossRef] [Green Version]
- Melenchón, F.; Larrán, A.M.; Mercado, E.; Hidalgo, M.C.; Cardenetem, G.; Barroso, F.G.; Fabrikov, D.; Lourenco, H.M.; Pessoa, M.F.; Tomás-Almenar, C. potential use of black soldier fly (Hermetia illucens) and mealworm (Tenebrio molitor) insect meals in diets for rainbow trout (Onchohynchus mykiss). Aquac. Nutr. 2020, 27, 491–505. [Google Scholar] [CrossRef]
- Lee, K.S.; Yun, E.Y.; Goo, T.W. Antimicrobial Activity of an extract of Hermetia illucens larvae immunized with Lactobacillus casei against Salmonella species. Insects 2020, 11, 704. [Google Scholar] [CrossRef]
- Lee, K.S.; Yun, E.Y.; Goo, T.W. Evaluation of the antimicrobial activity of an extract of Lactobacillus casei-infected Hermetia illucens larvae produced using an automatic injection system. Animals 2020, 10, 2121. [Google Scholar] [CrossRef]
- Azevedo, P.A.; Cho, C.Y.; Leeson, S.; Bureau, D.P. Effect of feeding level and water temperature on growth, nutrient and energy utilization and waste outputs of rainbow trout (Oncorhynchus mykiss). Aquat. Living Resour. 1998, 11, 227–238. [Google Scholar] [CrossRef]
- William, S.H. Smolt transformation: Evolution, behavior, and physiology. J. Fish. Res. Board Can. 1976, 33, 1233–1252. [Google Scholar]
- Bureau, D.P.; Hua, K.; Cho, C.Y. Effect of feeding level on growth and nutrient deposition in rainbow trout (Oncorhynchus mykiss Walbaum) growing from 150 to 600 g. Aquac. Res. 2006, 37, 1090–1098. [Google Scholar] [CrossRef]
- Stadtlander, T.; Stamer, A.; Buser, A.; Wohlfahrt, J.; Leiber, F.; Sandrock, C. Hermetia illucens meal as fish meal replacement for rainbow trout on farm. J. Insects Food Feed 2017, 3, 165–175. [Google Scholar] [CrossRef] [Green Version]
- Maita, M. Fish health assessment. In Dietary Supplements for the Health and Quality of Cultured Fish; CABI Publishing: Wallingford, UK, 2007; pp. 10–34. [Google Scholar]
- Adams, S.M.; McLean, R.B. Estimation of largemouth bass, Micropterus salmoides Lacépède, growth using the liver somatic index and physiological variables. J. Fish Biol. 1985, 26, 111–126. [Google Scholar] [CrossRef]
- Barnham, C.; Baxter, A. Condition Factor, K, for Salmonid Fish; Fisheries Notes; Department of Primary Industries: Melbourne, VIC, Australia, 2003; pp. 1–3.
- Frasani, M.N.; Hoseinnifa, S.H.; Rashidian, G.; Farsani, H.G.; Ashouri, G.; Doan, H.V. Dietary effects of Coriandrum sativum extract on growth performance, phtsiological and innate immune responses and resistance of rainbow trout (Onchorhychus mykiss) against Yersinia recheri. Fish Shellfish Immunol. 2019, 91, 233–240. [Google Scholar]
- Yonar, M.E.; Yonar, S.M.; İspir, Ü.; Ural, M.Ş. Effect of curcumin on haematological valures, immunity, antioxidnat status and resistance of rainbow trout (Oncorrhynchus mykiss) against Aeromonas Salmonicida subsp. Achromogenes. Fish Shellfish Immunol. 2019, 89, 83–90. [Google Scholar] [CrossRef]
- McCarthy, D.H.; Stevenson, J.P.; Roberts, M.S. Some blood parameters of the rainbow trout (Salmo gairdneri Richardson). II. the shasta variety. J. Fish Biol. 1975, 7, 215–219. [Google Scholar] [CrossRef]
- Adel, M.; Amiri, A.A.; Zorriehzahre, J.; Nematolahi, A.; Esteban, M.Á. Effects of dietary peppermint (Mentha piperita) on growth performance, chemical body composition and hematological and immnune parameters of fry caspian white fish (Rutilus frisii kutem). Fish Shellfish Immunol. 2015, 45, 841–847. [Google Scholar] [CrossRef]
- Shen, Y.; Wang, D.; Zhao, J.; Chen, X. Fish red blood cells express immune genes and responses. Aquqcul. Fish. 2018, 3, 14–21. [Google Scholar] [CrossRef]
- Lindjay, G.J.H.; Walton, M.J.; Adoeon, J.W.; Fletcher, T.C.; Cho, C.Y.; Cowey, C.B. The growth of rainbow trout (Salmo gaidneri) given diets contaning chitin and its relationship to chitinolytic enzyme and chitin digestibilty. Aquaculture 1984, 37, 315–334. [Google Scholar] [CrossRef]
- Moriyama, S.; Ayson, F.G.; Kawauchi, H. Growth regulation by insulin like growth factor-1 in fish. Biosci. Biotechnol. Biochem. 2000, 64, 1553–1562. [Google Scholar] [CrossRef]
- Bruslé, J.; Anadon, G.G. Fish Morphology, 1st ed.; Taylor & Francis Group: London, UK, 1996; p. 17. [Google Scholar]
- Castro, R.; Abós, B.; Pignatelli, J.; Jørgensen, L.G.; Granza, A.G.; Buchmann, K.; Tafalla, C. Early immune response in rainbow trout liver upon viral hemorrhagic septicemia virus (VHSV) infection. PLoS ONE 2014, 9, e111084. [Google Scholar] [CrossRef]
- Huang, X.J.; Choi, Y.K.; Im, H.S.; Yarimaga, O.; Yoon, E.; Kim, H.S. Aspartate aminotransferase (AST/GOT) and alanine aminotransferase (ALT/GPT) detection techniques. Sensors 2006, 6, 756–782. [Google Scholar] [CrossRef] [Green Version]
- Trullàs, C.; Fontanillas, R.; Tres, A.; Barroeta, A.C.; Sala, R. Acid and re-esterified rapeseed oils as alternative vegetable oils for rainbow trout diets: Effects on lipid digestibility and growth. Aquaculture 2016, 451, 186–194. [Google Scholar] [CrossRef]
- Rey Vázquez, G.R.; Guerrero, G.A. characterization of blood cells and hematological parameters in Cichlasoma dimerus (Teleostei, Perciformes). Tissue Cell 2007, 39, 151–160. [Google Scholar] [CrossRef]
- Kim, J.D.; Kim, Y.C.; Kim, K.S.; Woo, S.H.; Park, S.W. The effect of omae Prunus mume extract on the immune response and growth rate of Japanese eel Anguilla japonica. Fish Pathol. 2009, 22, 367–374. [Google Scholar]
- Saurabh, S.; Sahoo, P.K. Lysozyme: An important defense molecule of fish innate immune system. Aquac. Res. 2008, 39, 223–239. [Google Scholar] [CrossRef]
- Tippayadara, N.; Dawood, M.A.O.; Krutmuang, P.; Hoseinifar, S.H.; Doan, H.V.; Paolucci, M. Replacement of fish meal by black soldier fly (Hermetia illucens) larvae meal: Effects on grwoth haematology, and skin mucus immunity of nile tilapia, Oreochromis niloticus. Animals 2021, 11, 193. [Google Scholar] [CrossRef]
- Ingram, G.A. Substances involved in the natural-resistance of fish to infection—A review. J. Fish Biol. 1980, 16, 23–60. [Google Scholar] [CrossRef]
- Nagae, M.; Fuda, H.; Ura, K.; Kawamura, H.; Adachi, S.; Hara, A.; Yamauchi, K. The effect of cortisol administration on blood plasma immunoglobulin M (IgM) concentrations in masu salmon (Oncorhynchus-Masou). Fish Physiol. Biochem. 1994, 13, 41–48. [Google Scholar] [CrossRef]
- Giri, S.S.; Sen, S.S.; Chi, C.; Kim, H.J.; Yun, S.; Park, S.C.; Sukumaran, V. Effects of guava leaves on the growth performance and cytokine gene expression of Labeo rohita and its susceptibility to Aeromonas hydrophila infection. Fish Shellfish Immunol. 2015, 46, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Giri, S.S.; Sukumaran, V.; Sen, S.S.; Jena, P.K. Effects of dietary supplementation of potential probiotic Bacillus subtilis VSG1 singularly or in combination with Lactobacillus plantarum VSG3 or/and Pseudomonas aeruginosa VSG2 on the growth, immunity and disease resistance of Labeo rohita. Aquac. Nutr. 2013, 20, 163–171. [Google Scholar] [CrossRef]
- Mashoof, S.; Criscitiello, M.F. Fish immunoglobulins. Biology 2016, 5, 45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Candiotto, A.; Bo, T.; Fenoglio, S. Biological and ecological data on an established rainbow trout (Oncorhynchus mykiss) population in an italian stream. Fundam. Appl. Limnol. 2011, 179, 64–76. [Google Scholar] [CrossRef]
- Madrazo, A.L.; Camposm, M.R.S. Review of antimicrobial peptides as promoters of food safety: Limitations and possibilities within the food industry. J. Food Saf. 2020, 40, e12854. [Google Scholar] [CrossRef]
- Naylor, R.L.; Goldburg, R.J.; Primavera, J.H.; Kautsky, N.; Beveridge, M.C.M.; Clay, J.; Folke, C.; Lubchenco, J.; Mooney, H.; Troell, M. Effect of aquaculture on world fish supplies. Nature 2000, 405, 1017–1024. [Google Scholar] [CrossRef] [PubMed] [Green Version]

{kind=link}
{kind=link}
{kind=link}
| Name | Sequence | |
|---|---|---|
| HiCec1 (Accession: JX997953) | Forward | 5’-TTGGTCAACGAGTTCGTGATGC-3’ |
| Reverse | 5’-TCCTTGTTGTGGTGGTCCACCT-3’ | |
| HiDef1 (Accession: KF805347) | Forward | 5’-AGGTGGTGGAGCAGCATTAC-3’ |
| Reverse | 5’-ACGACGTCCCAAAGCAATAC-3’ | |
| Act5C (Accession: P10987) | Forward | 5′-AAGGACTCGTACGTGGGTG-3′ |
| Reverse | 5′-CATCTTCTCACGGTTGGC-3′ | |
| HIL0 | HIL25 | HIL50 | ImHIL25 | ImHIL50 | |
|---|---|---|---|---|---|
| Ingredients, g kg−1 | |||||
| Fish meal | 650 | 470 | 285 | 480 | 310 |
| Fish oil | 100 | 80 | 63 | 100 | 105 |
| Noninduced HI | - | 250 | 500 | - | - |
| Immune induced HI | - | - | - | 250 | 500 |
| Wheat meal | 200 | 150 | 102 | 120 | 35 |
| Gelatinized starch (D500) | 50 | 50 | 50 | 50 | 50 |
| Mineral mixture | 10 | 10 | 10 | 10 | 10 |
| Vitamin Mixture | 10 | 10 | 10 | 10 | 10 |
| Proximate composition | |||||
| DM, g kg−1 | 943.5 | 940 | 937.4 | 936.9 | 938.3 |
| CP, g kg−1 | 455.9 | 449.3 | 444.2 | 449.5 | 447.0 |
| EE, g kg−1 | 153.1 | 157.9 | 177.9 | 158.3 | 177.0 |
| Carbohydrate, g kg−1 | 184.4 | 189.6 | 145.0 | 155.6 | 147.8 |
| Ash, g kg−1 | 132.4 | 130.7 | 140.7 | 126.6 | 147.8 |
| Gross energy, cal g−1 | 4900.0 | 4899.0 | 4919.5 | 4808.0 | 4981.5 |
| Week | HIL0 | HIL25 | HIL50 | ImHIL25 | ImHIL50 | |
|---|---|---|---|---|---|---|
| IBW (g) | 0 | 170.90 ±12.83 a | 178.44 ± 12.21 a | 176.45 ± 0.10 a | 177.54 ± 0.32 a | 167.71 ± 12.80 a |
| IL (cm) | 0 | 24.55 ± 0.68 a | 24.94 ± 0.14 a | 24.67 ± 0.21 a | 24.76 ±0.05 a | 24.05 ± 0.61 a |
| SR (%) | 3 | 100 a | 100 a | 100 a | 100 a | 100 a |
| 6 | 100 a | 96.7 a | 100 a | 100 a | 100 a | |
| BW (g) | 3 | 198.45 ± 0.27 ab | 206.54 ± 7.35 a | 202.05 ± 8.12 ab | 202.16 ± 4.63 ab | 189.21 ± 3.70 b |
| 6 | 226.98 ± 2.25 a | 244.01 ± 10.19 a | 235.97 ± 24.12 a | 237.05 ± 33.68 a | 222.03 ± 17.11 a | |
| WG 1 (%) | 3 | 16.12 ± 0.94 a | 15.75 ± 6.20 b | 14.51 ± 0.99 b | 13.87 ± 1.54 b | 12.82 ± 2.52 c |
| 6 | 32.85 ± 0.86 a | 37.10 ± 7.93 a | 33.73 ± 5.68 a | 33.52 ± 6.10 a | 32.51 ± 3.69 a | |
| SGR 2 (%/day) | 6 | 0.676 ± 0.002 c | 0.745 ± 0.001 a | 0.692 ± 0.003 b | 0.688 ± 0.001 b | 0.668 ± 0.001 c |
| FCR 3 (%) | 3 | 76.68 ± 10.21 a | 76.60 ± 24.18 a | 71.14 ± 5.05 a | 68.40 ± 7.72 b | 59.15 ± 6.75 b |
| 6 | 44.49 ± 1.44 b | 59.36 ± 13.08 a | 52.99 ± 13.08 a | 54.51 ± 17.07 a | 51.60 ± 7.90 a | |
| HIS 4 | 3 | 0.83 ± 0.06 b | 0.91 ± 0.13 a | 0.83 ± 0.01 b | 0.85 ± 0.22 b | 0.77 ± 0.11 c |
| 6 | 0.86 ± 0.12 b | 0.95 ± 0.22 a | 0.86 ± 0.06 b | 0.98 ± 0.30 a | 0.89 ± 0.09 b | |
| HGR 5 (%) | 3.61 ± 0.08 b | 4.40 ± 0.14 b | 3.61 ± 0.07 b | 15.29 ± 0.21 a | 15.58 ± 0.08 a | |
| Obesity | 3 | 1.14 ± 0.01 b | 1.18 ± 0.03 a | 1.18 ± 0.01 a | 1.19 ± 0.03 a | 1.19 ± 0.02 a |
| 6 | 1.18 ± 0.02 a | 1.15 ± 0.08 b | 1.17 ± 0.01 a | 1.15 ± 0.01 b | 1.18 ± 0.01 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hwang, D.; Lim, C.-H.; Lee, S.H.; Goo, T.-W.; Yun, E.-Y. Effect of Feed Containing Hermetia illucens Larvae Immunized by Lactobacillus plantarum Injection on the Growth and Immunity of Rainbow Trout (Oncorhynchus mykiss). Insects 2021, 12, 801. https://doi.org/10.3390/insects12090801
Hwang D, Lim C-H, Lee SH, Goo T-W, Yun E-Y. Effect of Feed Containing Hermetia illucens Larvae Immunized by Lactobacillus plantarum Injection on the Growth and Immunity of Rainbow Trout (Oncorhynchus mykiss). Insects. 2021; 12(9):801. https://doi.org/10.3390/insects12090801
Chicago/Turabian StyleHwang, Dooseon, Chae-Hwan Lim, Seung Hun Lee, Tae-Won Goo, and Eun-Young Yun. 2021. "Effect of Feed Containing Hermetia illucens Larvae Immunized by Lactobacillus plantarum Injection on the Growth and Immunity of Rainbow Trout (Oncorhynchus mykiss)" Insects 12, no. 9: 801. https://doi.org/10.3390/insects12090801
APA StyleHwang, D., Lim, C.-H., Lee, S. H., Goo, T.-W., & Yun, E.-Y. (2021). Effect of Feed Containing Hermetia illucens Larvae Immunized by Lactobacillus plantarum Injection on the Growth and Immunity of Rainbow Trout (Oncorhynchus mykiss). Insects, 12(9), 801. https://doi.org/10.3390/insects12090801