
Antennal Transcriptome Analysis and Identification of Candidate Chemosensory Genes of the Harlequin Ladybird Beetle, Harmonia axyridis (Pallas) (Coleoptera: Coccinellidae)

 , ,
, ,  , ,
, ,  and
and 
Abstract
Simple Summary
Abstract
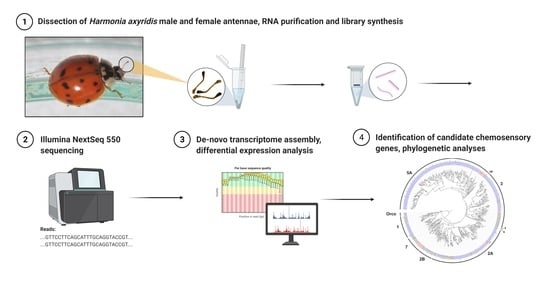
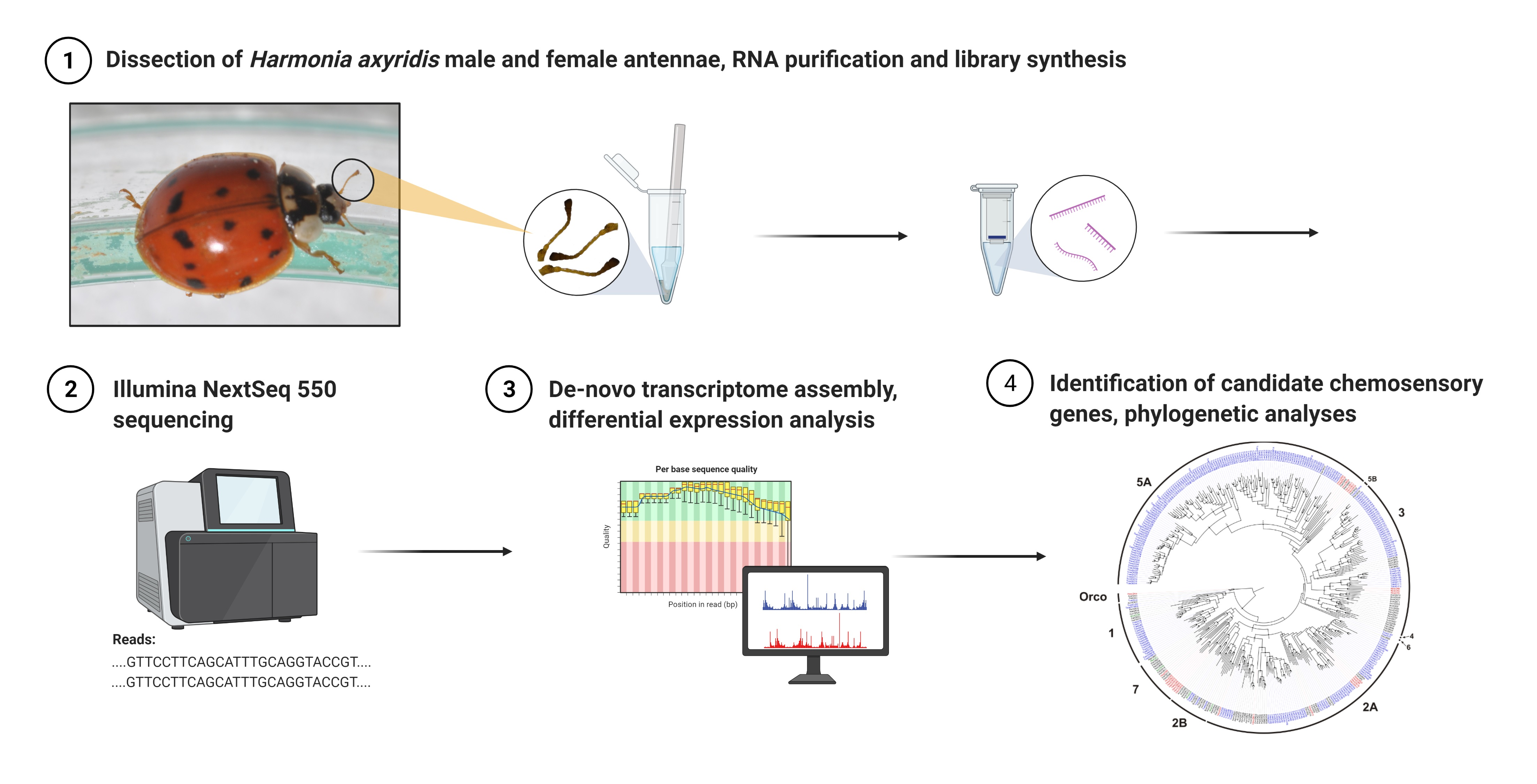{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Insect Rearing and Antenna Collection
2.2. RNA Purification, cDNA Library Preparation and Sequencing
2.3. Assembly and Functional Annotation
2.4. Identification of Differentially Expressed Transcripts
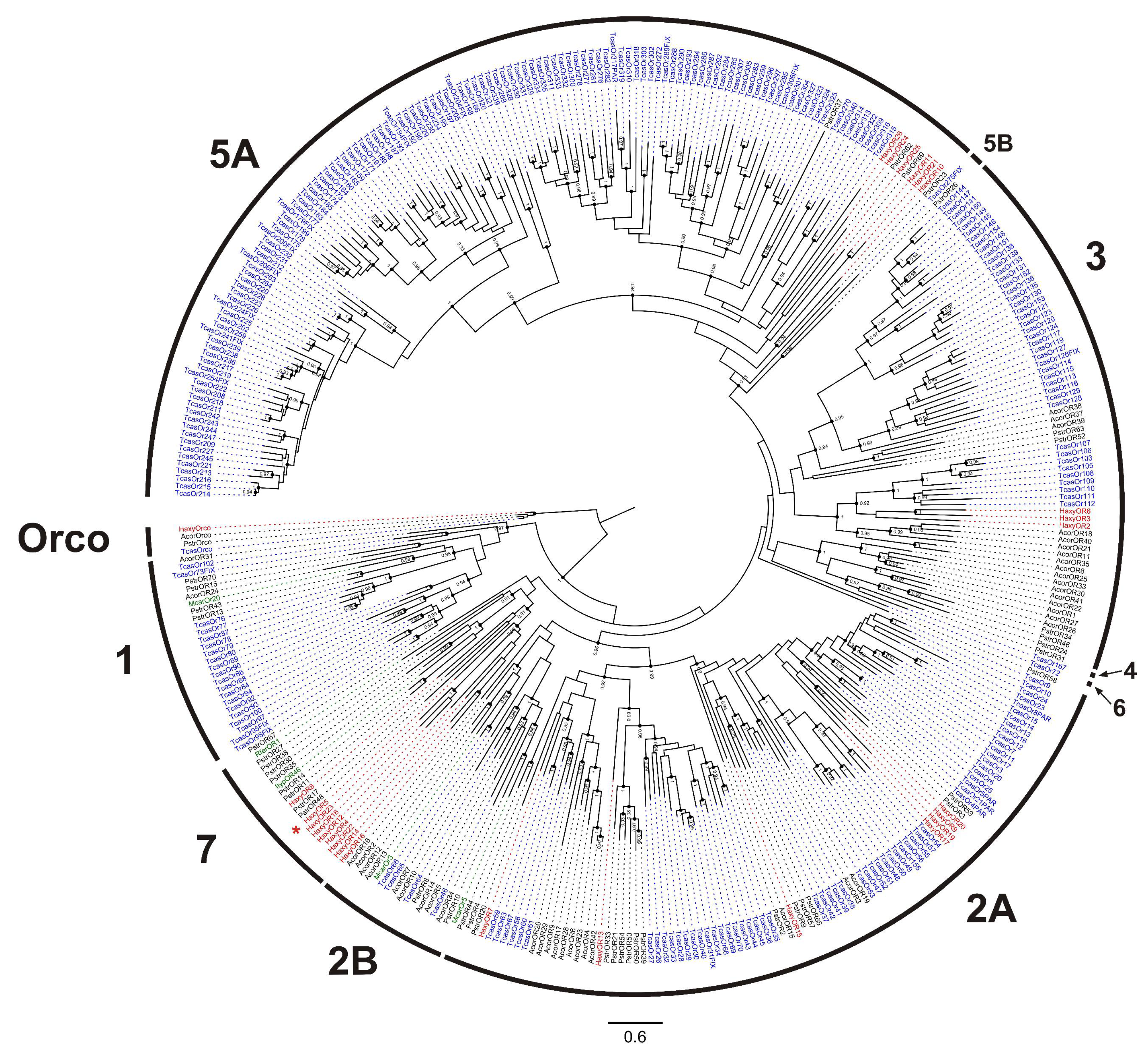
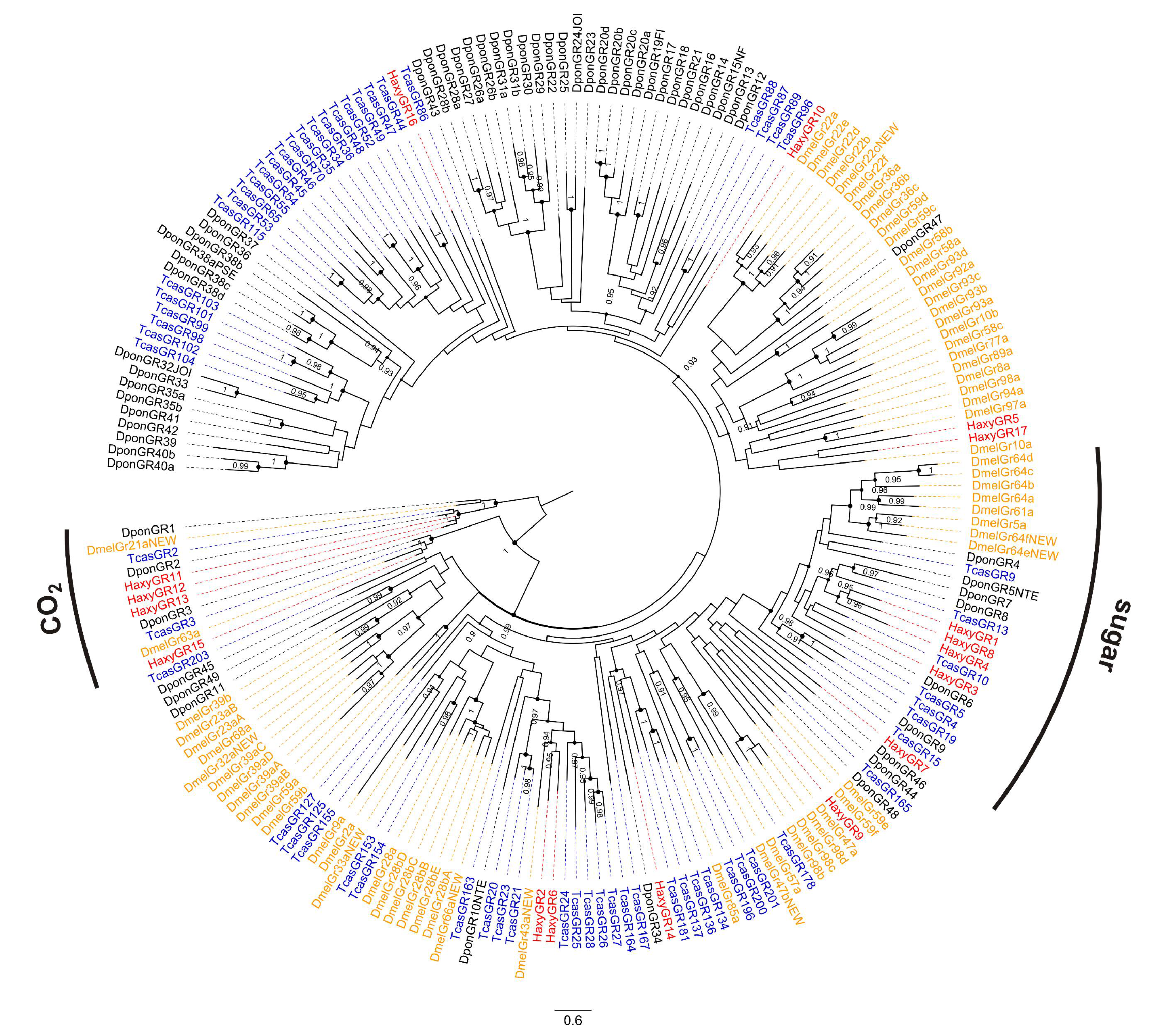
2.5. Identification of Chemosensory Genes and Phylogenetic Analyses
3. Results
3.1. Transcriptome Sequencing, Assembly, and Identification of Chemosensory Genes
3.2. Odorant Receptors (ORs)
3.3. Gustatory Receptors (GRs)
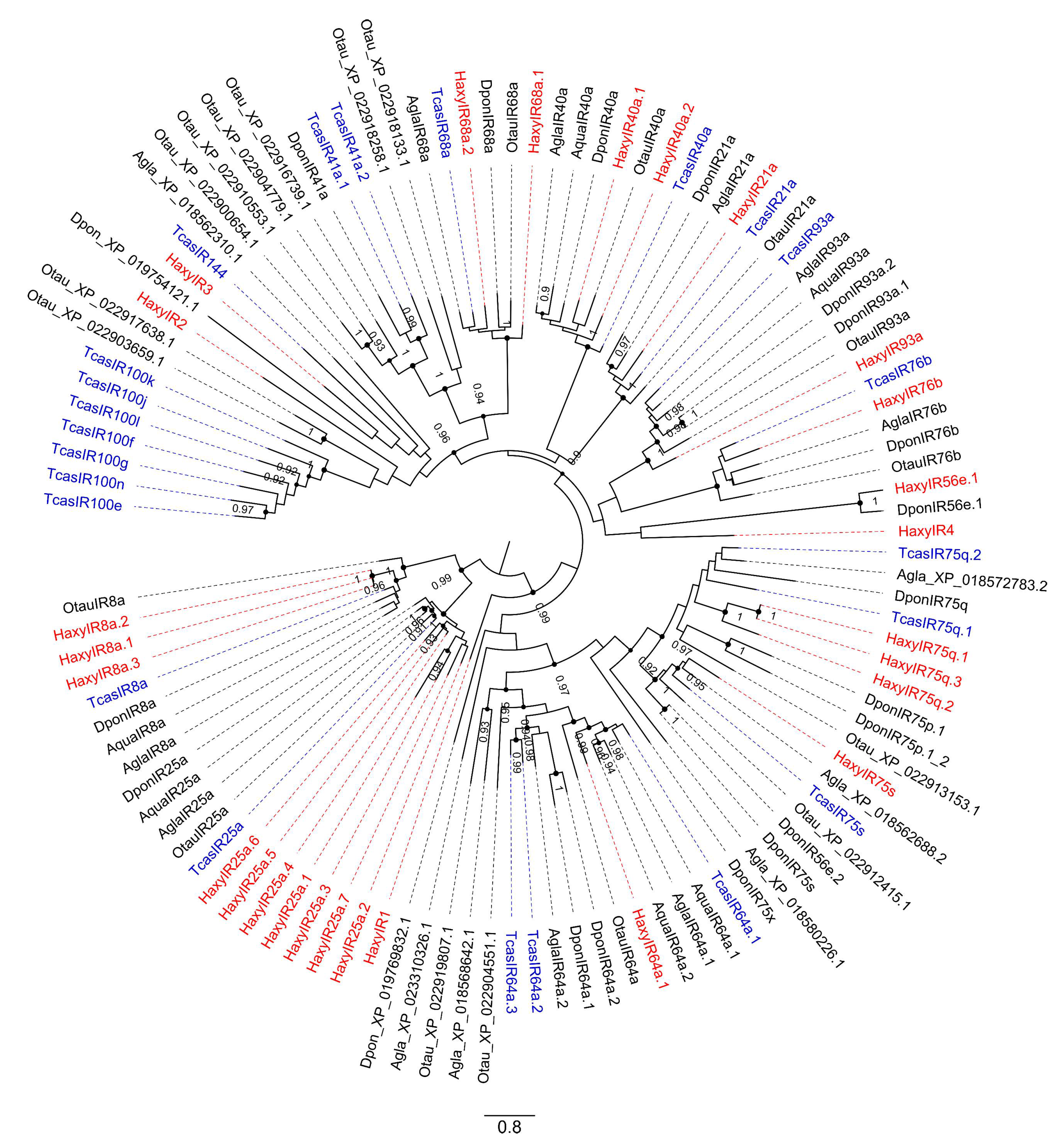
3.4. Ionotropic Receptors (IRs)
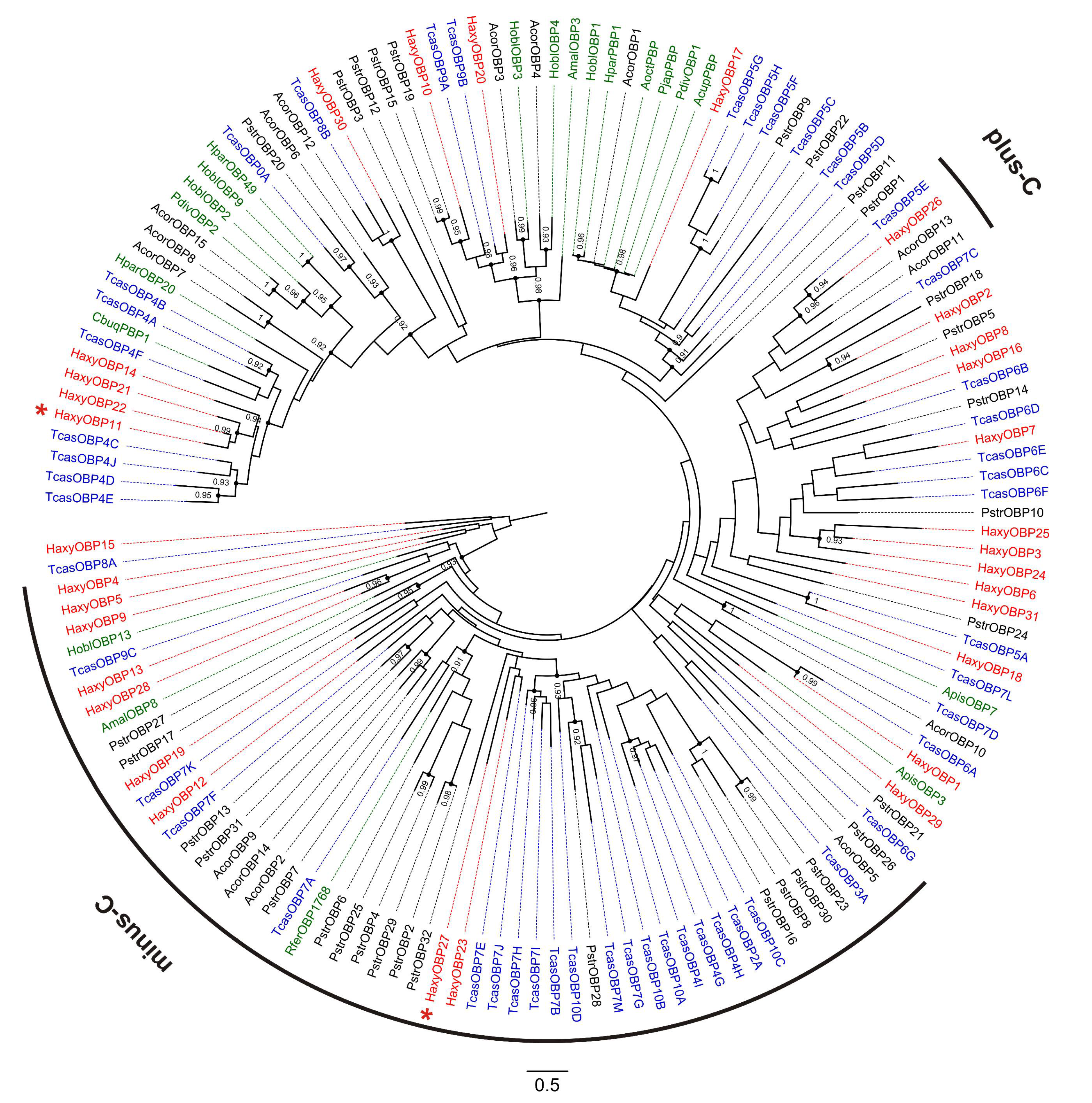
3.5. Odorant-Binding Proteins (OBPs)
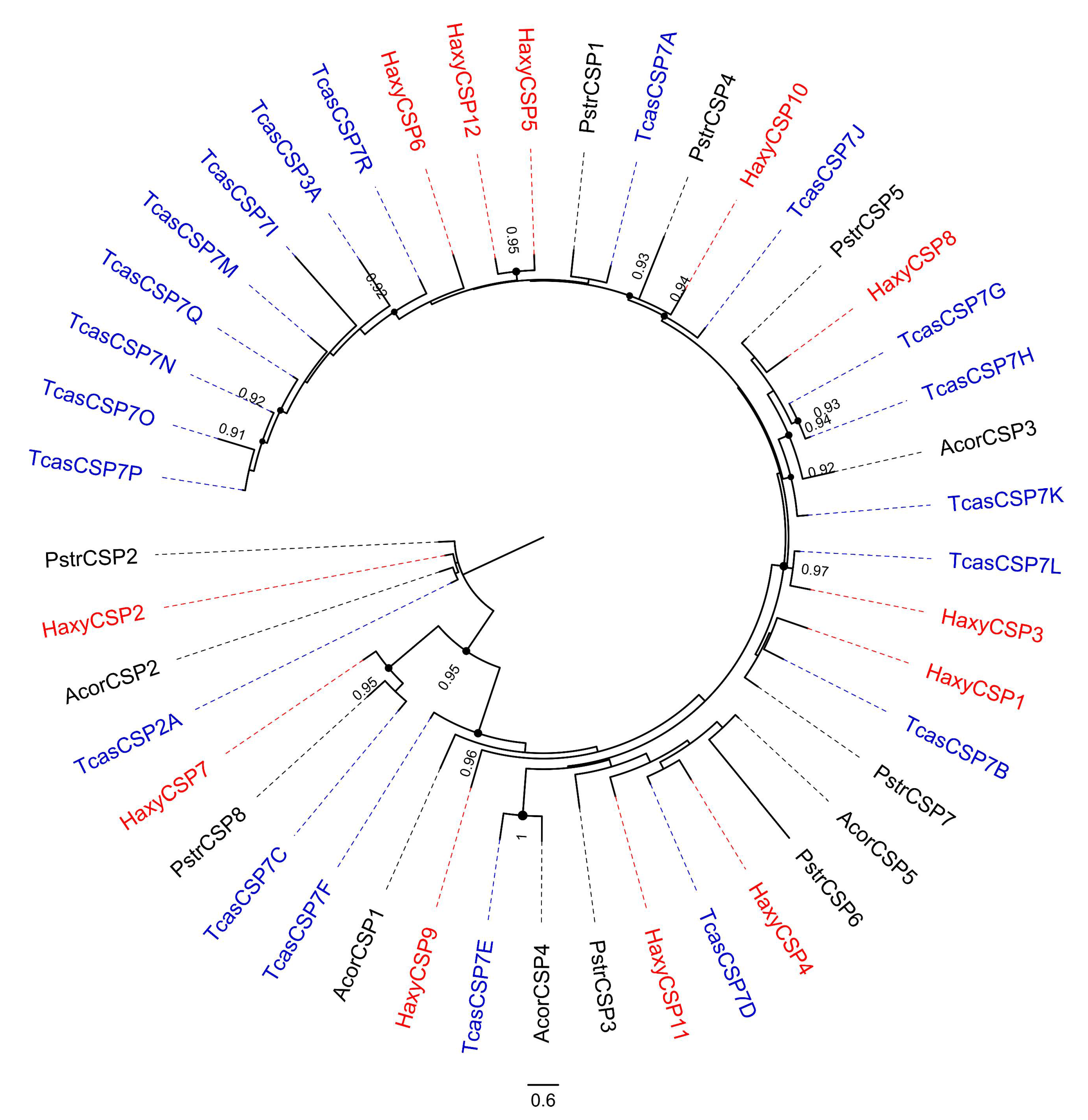
3.6. Chemosensory Proteins (CSPs)
3.7. Sensory Neuron Membrane Proteins (SNMPs)
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Pettersson, J. Coccinellids and Semiochemicals. In Ecology and Behaviour of the Ladybird Beetles (Coccinellidae); Hodek, I., van Emden, H.F., Honek, A., Eds.; John Wiley & Sons: Chichester, UK, 2012; pp. 444–464. [Google Scholar]
- Hamilton, R.M.; Dogan, E.B.; Schaalje, G.B.; Booth, G.M. Olfactory response of the lady beetle Hippodamia convergens (Coleoptera: Coccinellidae) to prey related odors, including a scanning electron microscopy study of the antennal sensilla. Environ. Entomol. 1999, 28, 812–822. [Google Scholar] [CrossRef]
- Haverkamp, A.; Hansson, B.S.; Knaden, M. Combinatorial codes and labeled lines: How insects use olfactory cues to find and judge food, mates, and oviposition sites in complex environments. Front. Physiol. 2018, 9, 46. [Google Scholar] [CrossRef]
- Leal, W.S. Odorant reception in insects: Roles of receptors, binding proteins, and degrading enzymes. Annu. Rev. Entomol. 2013, 58, 373–391. [Google Scholar] [CrossRef] [PubMed]
- Andersson, M.N.; Löfstedt, C.; Newcomb, R.D. Insect olfaction and the evolution of receptor tuning. Front. Ecol. Evol. 2015, 53. [Google Scholar] [CrossRef]
- Jones, W.D.; Cayirlioglu, P.; Grunwald Kadow, I.; Vosshall, L.B. Two chemosensory receptors together mediate carbon dioxide detection in Drosophila. Nature 2007, 445, 86–90. [Google Scholar] [CrossRef]
- Montagné, N.; De Fouchier, A.; Newcomb, R.D.; Jacquin-Joly, E. Advances in the identification and characterization of olfactory receptors in insects. Prog. Mol. Biol. Transl. Sci. 2015, 130, 55–80. [Google Scholar] [CrossRef] [PubMed]
- Robertson, H.M. Molecular evolution of the major arthropod chemoreceptor gene families. Annu. Rev. Entomol. 2019, 64, 227–242. [Google Scholar] [CrossRef]
- Mitchell, R.F.; Schneider, T.M.; Schwartz, A.M.; Andersson, M.N.; McKenna, D.D. The diversity and evolution of odorant receptors in beetles (Coleoptera). Insect Mol. Biol. 2020, 29, 77–91. [Google Scholar] [CrossRef]
- He, P.; Durand, N.; Dong, S.L. Editorial: Insect olfactory proteins (from gene identification to functional characterization). Front. Physiol. 2019, 10, 1313. [Google Scholar] [CrossRef] [PubMed]
- De Santis, F.; François, M.C.; Merlin, C.; Pelletier, J.; Maïbèche-Coisné, M.; Conti, E.; Jacquin-Joly, E. Molecular cloning and in Situ expression patterns of two new pheromone-binding proteins from the corn stemborer Sesamia nonagrioides. J. Chem. Ecol. 2006, 32, 1703–1717. [Google Scholar] [CrossRef] [PubMed]
- De Bruyne, M.; Foster, K.; Carlson, J.R. Odor coding in the Drosophila antenna. Neuron 2001, 30, 537–552. [Google Scholar] [CrossRef]
- Robertson, H.M.; Warr, C.G.; Carlson, J.R. Molecular evolution of the insect chemoreceptor gene superfamily in Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 2003, 100, 14537–14542. [Google Scholar] [CrossRef]
- Li, Z.Q.; Zhang, S.; Ma, Y.; Luo, J.Y.; Wang, C.Y.; Lv, L.M.; Dong, S.L.; Cui, J.J. First transcriptome and digital gene expression analysis in Neuroptera with an emphasis on chemoreception genes in Chrysopa pallens (Rambur). PLoS ONE 2013, 8, e67151. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.Q.; Zhang, S.; Luo, J.Y.; Wang, S.B.; Wang, C.Y.; Lv, L.M.; Dong, S.L.; Cui, J.J. Identification and expression pattern of candidate olfactory genes in Chrysoperla sinica by antennal transcriptome analysis. Comp. Biochem. Physiol. Part D Genom. Proteom. 2015, 15, 28–38. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.Y.; Zhu, M.F.; Jiang, Y.D.; Zhou, W.W.; Liu, S.; Heong, K.L.; Cheng, J.; Zhu, Z.R. Identification of candidate odorant-binding protein and chemosensory protein genes in Cyrtorhinus lividipennis (Hemiptera: Miridae), a key predator of the rice planthoppers in Asia. Environ. Entomol. 2017, 46, 654–662. [Google Scholar] [CrossRef]
- Wang, G.Y.; Zhu, J.L.; Zhou, W.W.; Liu, S.; Khairul, Q.M.; Ansari, N.A.; Zhu, Z.R. Identification and expression analysis of putative chemoreception genes from Cyrtorhinus lividipennis (Hemiptera: Miridae) antennal transcriptome. Sci. Rep. 2018, 8, 12981. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Liu, Y.; Wang, G.R. Chemosensory genes in the antennal transcriptome of two syrphid species, Episyrphus balteatus and Eupeodes corollae (Diptera: Syrphidae). BMC Genom. 2017, 18, 586. [Google Scholar] [CrossRef]
- Rondoni, G.; Borges, I.; Collatz, J.; Conti, E.; Costamagna, A.C.; Dumont, F.; Evans, E.W.; Grez, A.A.; Howe, A.G.; Lucas, E.; et al. Exotic ladybirds for biological control of herbivorous insects—a review. Entomol. Exp. Appl. 2021, 169, 6–27. [Google Scholar] [CrossRef]
- Michaud, J.P. Coccinellids in biological control. In Ecology and Behaviour of the Ladybird Beetles (Coccinellidae); Hodek, I., van Emden, H.F., Honek, A., Eds.; John Wiley & Sons: Chichester, UK, 2012; pp. 488–519. [Google Scholar]
- Obrycki, J.J.; Harwood, J.D.; Kring, T.J.; O’Neil, R.J. Aphidophagy by Coccinellidae: Application of biological control in agroecosystems. Biol. Control 2009, 51, 244–254. [Google Scholar] [CrossRef]
- Hodek, I.; van Emden, H.F.; Honěk, A. Ecology and Behaviour of the Ladybird Beetles (Coccinellidae); John Wiley & Sons: Chichester, UK, 2012. [Google Scholar]
- Rondoni, G.; Ielo, F.; Ricci, C.; Conti, E. Intraguild predation responses in two aphidophagous coccinellids identify differences among juvenile stages and aphid densities. Insects 2014, 5, 974–983. [Google Scholar] [CrossRef]
- Roy, H.E.; Brown, P.M.J.; Adriaens, T.; Berkvens, N.; Borges, I.; Clusella-Trullas, S.; De Clercq, P.; Comont, R.F.; Eschen, R.; Estoup, A.; et al. The harlequin ladybird, Harmonia axyridis: Global perspectives on invasion history and ecology. Biol. Invasions 2016, 18, 997–1044. [Google Scholar] [CrossRef]
- Koch, R.L.; Costamagna, A.C. Reaping benefits from an invasive species: Role of Harmonia axyridis in natural biological control of Aphis glycines in North America. BioControl 2017. [Google Scholar] [CrossRef]
- Gagnon, A.-È.; Doyon, J.; Heimpel, G.E.; Brodeur, J. Prey DNA detection success following digestion by intraguild predators: Influence of prey and predator species. Mol. Ecol. Resour. 2011, 11, 1022–1032. [Google Scholar] [CrossRef] [PubMed]
- Hautier, L.; San Martin, G.; Callier, P.; de Biseau, J.-C.; Grégoire, J.-C. Alkaloids provide evidence of intraguild predation on native coccinellids by Harmonia axyridis in the field. Biol. Invasions 2011, 13, 1805–1814. [Google Scholar] [CrossRef]
- Rondoni, G.; Onofri, A.; Ricci, C. Differential susceptibility in a specialised aphidophagous ladybird, Platynaspis luteorubra (Coleoptera: Coccinellidae), facing intraguild predation by exotic and native generalist predators. Biocontrol Sci. Technol. 2012, 22, 1334–1350. [Google Scholar] [CrossRef]
- Howe, A.G.; Ravn, H.P.; Pipper, C.B.; Aebi, A. Potential for exploitative competition, not intraguild predation, between invasive harlequin ladybirds and flowerbugs in urban parks. Biol. Invasions 2016, 18, 517–532. [Google Scholar] [CrossRef]
- Li, H.; Li, B.; Lövei, G.L.; Kring, T.J.; Obrycki, J.J. Interactions among native and non-native predatory Coccinellidae influence biological control and biodiversity. Ann. Entomol. Soc. Am. 2020, 1–18. [Google Scholar] [CrossRef]
- Rondoni, G.; Ielo, F.; Ricci, C.; Conti, E. Behavioural and physiological responses to prey-related cues reflect higher competitiveness of invasive vs. native ladybirds. Sci. Rep. 2017, 7, 3716. [Google Scholar] [CrossRef] [PubMed]
- Leroy, P.D.; Heuskin, S.; Sabri, A.; Verheggen, F.J.; Farmakidis, J.; Lognay, G.; Thonart, P.; Wathelet, J.-P.; Brostaux, Y.; Haubruge, E. Honeydew volatile emission acts as a kairomonal message for the Asian lady beetle Harmonia axyridis (Coleoptera: Coccinellidae). Insect Sci. 2012, 19, 498–506. [Google Scholar] [CrossRef]
- Sevarika, M.; Rondoni, G.; Conti, E.; Romani, R. Antennal sensory organs and glands of the harlequin ladybird, Harmonia axyridis. Entomol. Exp. Appl. 2021, 169, 111–124. [Google Scholar] [CrossRef]
- Liu, G.; Arnaud, P.; Offmann, B.; Picimbon, J.-F. Pheromone, Natural Odor and Odorant Reception Suppressing Agent (ORSA) for Insect Control. In Olfactory Concepts of Insect Control—Alternative to insecticides; Picimbon, J.F., Ed.; Springer: Cham, Switzerland, 2019. [Google Scholar]
- Jia, X.J.; Wang, H.X.; Yan, Z.G.; Zhang, M.Z.; Wei, C.H.; Qin, X.C.; Ji, W.R.; Falabella, P.; Du, Y.L. Antennal transcriptome and differential expression of olfactory genes in the yellow peach moth, Conogethes punctiferalis (Lepidoptera: Crambidae). Sci. Rep. 2016, 6, 29067. [Google Scholar] [CrossRef]
- Afgan, E.; Baker, D.; Batut, B.; Van Den Beek, M.; Bouvier, D.; Ech, M.; Chilton, J.; Clements, D.; Coraor, N.; Grüning, B.A.; et al. The Galaxy platform for accessible, reproducible and collaborative biomedical analyses: 2018 update. Nucleic Acids Res. 2018, 46, 537–544. [Google Scholar] [CrossRef]
- Andrews, S. FastQC—A Quality Control Tool for High Throughput Sequence Data. 2010. Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 16 June 2018).
- Blankenberg, D.; Gordon, A.; Von Kuster, G.; Coraor, N.; Taylor, J.; Nekrutenko, A.; Team, G. Manipulation of FASTQ data with galaxy. Bioinformatics 2010, 26, 1783–1785. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Schmieder, R.; Lim, Y.W.; Edwards, R. Identification and removal of ribosomal RNA sequences from metatranscriptomes. Bioinformatics 2012, 28, 433–435. [Google Scholar] [CrossRef]
- Haas, B.J.; Papanicolaou, A.; Yassour, M.; Grabherr, M.; Blood, P.D.; Bowden, J.; Couger, M.B.; Eccles, D.; Li, B.; Lieber, M.; et al. De novo transcript sequence reconstruction from RNA-seq using the Trinity platform for reference generation and analysis. Nat. Protoc. 2013, 8, 1494–1512. [Google Scholar] [CrossRef]
- Fu, L.; Niu, B.; Zhu, Z.; Wu, S.; Li, W. CD-HIT: Accelerated for clustering the next-generation sequencing data. Bioinformatics 2012, 28, 3150–3152. [Google Scholar] [CrossRef]
- Conesa, A.; Götz, S. Blast2GO: A comprehensive suite for functional analysis in plant genomics. Int. J. Plant Genom. 2008, 2008, 619832. [Google Scholar] [CrossRef] [PubMed]
- Gurevich, A.; Saveliev, V.; Vyahhi, N.; Tesler, G. QUAST: Quality assessment tool for genome assemblies. Bioinformatics 2013, 29, 1072–1075. [Google Scholar] [CrossRef] [PubMed]
- Simão, F.A.; Waterhouse, R.M.; Ioannidis, P.; Kriventseva, E.V.; Zdobnov, E.M. BUSCO: Assessing genome assembly and annotation completeness with single-copy orthologs. Bioinformatics 2015, 31, 3210–3212. [Google Scholar] [CrossRef] [PubMed]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011. [Google Scholar] [CrossRef]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. EdgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, D.; Price, D.C.; Chan, C.X.; Qiu, H.; Rose, N.; Ball, S.; Weber, A.P.M.; Cecilia Arias, M.; Henrissat, B.; Coutinho, P.M.; et al. Genome of the red alga Porphyridium purpureum. Nat. Commun. 2013, 4, 1941. [Google Scholar] [CrossRef] [PubMed]
- Masuko, K.; Fuse, N.; Komaba, K.; Katsuyama, T.; Nakajima, R.; Furuhashi, H.; Kurata, S. Winged eye induces transdetermination of Drosophila imaginal disc by acting in concert with a histone methyltransferase, Su(var)3-9. Cell Rep. 2018, 22, 206–217. [Google Scholar] [CrossRef]
- Zhao, W.; Lu, L.; Yang, P.; Cui, N.; Kang, L.; Cui, F. Organ-specific transcriptome response of the small brown planthopper toward rice stripe virus. Insect Biochem. Mol. Biol. 2016, 70, 60–72. [Google Scholar] [CrossRef]
- Robinson, M.D.; Oshlack, A. A scaling normalization method for differential expression analysis of RNA-seq data. Genome Biol. 2010, 11, R25. [Google Scholar] [CrossRef]
- Krogh, A.; Larsson, B.; Von Heijne, G.; Sonnhammer, E.L.L. Predicting transmembrane protein topology with a hidden Markov model: Application to complete genomes. J. Mol. Biol. 2001, 305, 567–580. [Google Scholar] [CrossRef] [PubMed]
- Bendtsen, J.D.; Nielsen, H.; Von Heijne, G.; Brunak, S. Improved prediction of signal peptides: SignalP 3.0. J. Mol. Biol. 2004, 340, 783–795. [Google Scholar] [CrossRef]
- Paula, D.P.; Togawa, R.C.; Costa, M.M.C.; Grynberg, P.; Martins, N.F.; Andow, D.A. Identification and expression profile of odorant-binding proteins in Halyomorpha halys (Hemiptera: Pentatomidae). Insect Mol. Biol. 2016, 25, 580–594. [Google Scholar] [CrossRef]
- Zhang, S.Q.; Che, L.H.; Li, Y.; Dan, L.; Pang, H.; Ślipiński, A.; Zhang, P. Evolutionary history of Coleoptera revealed by extensive sampling of genes and species. Nat. Commun. 2018, 9, 205. [Google Scholar] [CrossRef]
- Dippel, S.; Oberhofer, G.; Kahnt, J.; Gerischer, L.; Opitz, L.; Schachtner, J.; Stanke, M.; Schütz, S.; Wimmer, E.A.; Angeli, S. Tissue-specific transcriptomics, chromosomal localization, and phylogeny of chemosensory and odorant binding proteins from the red flour beetle Tribolium castaneum reveal subgroup specificities for olfaction or more general functions. BMC Genomics 2014, 15, 1141. [Google Scholar] [CrossRef]
- Richards, S.; Gibbs, R.A.; Weinstock, G.M.; Brown, S.; Denell, R.; Beeman, R.W.; Gibbs, R.; Bucher, G.; Friedrich, M.; Grimmelikhuijzen, C.J.P.; et al. The genome of the model beetle and pest Tribolium castaneum. Nature 2008, 452, 949–955. [Google Scholar] [CrossRef]
- Dippel, S.; Kollmann, M.; Oberhofer, G.; Montino, A.; Knoll, C.; Krala, M.; Rexer, K.H.; Frank, S.; Kumpf, R.; Schachtner, J.; et al. Morphological and transcriptomic analysis of a beetle chemosensory system reveals a gnathal olfactory center. BMC Biol. 2016, 14, 90. [Google Scholar] [CrossRef] [PubMed]
- Hu, P.; Wang, J.; Cui, M.; Tao, J.; Luo, Y. Antennal transcriptome analysis of the Asian longhorned beetle Anoplophora glabripennis. Sci. Rep. 2016, 6, 26652. [Google Scholar] [CrossRef] [PubMed]
- McKenna, D.D.; Scully, E.D.; Pauchet, Y.; Hoover, K.; Kirsch, R.; Geib, S.M.; Mitchell, R.F.; Waterhouse, R.M.; Ahn, S.J.; Arsala, D.; et al. Genome of the Asian longhorned beetle (Anoplophora glabripennis), a globally significant invasive species, reveals key functional and evolutionary innovations at the beetle-plant interface. Genome Biol. 2016, 17, 1–18. [Google Scholar] [CrossRef]
- Andersson, M.N.; Grosse-Wilde, E.; Keeling, C.I.; Bengtsson, J.M.; Yuen, M.M.S.; Li, M.; Hillbur, Y.; Bohlmann, J.; Hansson, B.S.; Schlyter, F. Antennal transcriptome analysis of the chemosensory gene families in the tree killing bark beetles, Ips typographus and Dendroctonus ponderosae (Coleoptera: Curculionidae: Scolytinae). BMC Genom. 2013, 14, 198. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Bin, S.; He, H.; Wang, Z.; Li, M.; Lin, J. Differential expression analysis of chemoreception genes in the striped flea beetle Phyllotreta striolata using a transcriptomic approach. PLoS ONE 2016, 11, e0153067. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Lin, L.; Xie, M.; Zhang, G.; Su, W. De novo sequencing, assembly and characterization of antennal transcriptome of Anomala corpulenta Motschulsky (Coleoptera: Rutelidae). PLoS ONE 2014, 9, e114238. [Google Scholar] [CrossRef]
- Li, X.; Ju, Q.; Jie, W.; Li, F.; Jiang, X.; Hu, J.; Qu, M. Chemosensory gene families in adult antennae of Anomala corpulenta Motschulsky (Coleoptera: Scarabaeidae: Rutelinae). PLoS ONE 2015, 10, e0121504. [Google Scholar] [CrossRef]
- Mitchell, R.F.; Hughes, D.T.; Luetje, C.W.; Millar, J.G.; Soriano-Agatón, F.; Hanks, L.M.; Robertson, H.M. Sequencing and characterizing odorant receptors of the cerambycid beetle Megacyllene caryae. Insect Biochem. Mol. Biol. 2012, 42, 499–505. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wang, S.; Yi, J.; Li, Y.; Liu, J.; Wang, J.; Xi, J. Three host plant volatiles, hexanal, lauric acid, and tetradecane, are detected by an antenna-biased expressed odorant receptor 27 in the dark black chafer Holotrichia parallela. J. Agric. Food Chem. 2020, 68, 7316–7323. [Google Scholar] [CrossRef]
- Antony, B.; Johny, J.; Montagné, N.; Jacquin-Joly, E.; Capoduro, R.; Cali, K.; Persaud, K.; Al-Saleh, M.A.; Pain, A. Pheromone receptor of the globally invasive quarantine pest of the palm tree, the red palm weevil (Rhynchophorus ferrugineus). bioRxiv 2020, 1–34. [Google Scholar] [CrossRef]
- Yuvaraj, J.K.; Roberts, R.E.; Sonntag, Y.; Hou, X.; Grosse-Wilde, E.; Machara, A.; Hansson, B.S.; Johanson, U.; Löfstedt, C.; Andersson, M.N. Putative ligand binding sites of two functionally characterized bark beetle odorant receptors. bioRxiv 2020, 1–19. [Google Scholar] [CrossRef]
- Nikonov, A.A.; Peng, G.; Tsurupa, G.; Leal, W.S. Unisex pheromone detectors and pheromone-binding proteins in scarab beetles. Chem. Senses 2002, 27, 495–504. [Google Scholar] [CrossRef]
- Cui, X.; Liu, D.; Sun, K.; He, Y.; Shi, X. Expression profiles and functional characterization of two odorant-binding proteins from the apple buprestid beetle Agrilus mali (Coleoptera: Buprestidae). J. Econ. Entomol. 2018, 111, 1420–1432. [Google Scholar] [CrossRef]
- Yang, H.; Su, T.; Yang, W.; Yang, C.P.; Chen, Z.M.; Lu, L.; Liu, Y.L.; Tao, Y.Y. Molecular characterization, expression pattern and ligand-binding properties of the pheromone-binding protein gene from Cyrtotrachelus buqueti. Physiol. Entomol. 2017, 42, 369–378. [Google Scholar] [CrossRef]
- Yin, J.; Wang, C.; Fang, C.; Zhang, S.; Cao, Y.; Li, K.; Leal, W.S. Functional characterization of odorant-binding proteins from the scarab beetle Holotrichia oblita based on semiochemical-induced expression alteration and gene silencing. Insect Biochem. Mol. Biol. 2019, 104, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Guan, L.; Zhong, T.; Li, K.; Yin, J.; Cao, Y. Potential cooperations between odorant-binding proteins of the scarab beetle Holotrichia oblita Faldermann (Coleoptera: Scarabaeidae). PLoS ONE 2013, 8, e84795. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Ju, Q.; Li, X.; Guo, X.Q.; Du, L.; Shi, C.R.; Qu, M.J. Two odorant-binding proteins of the dark black chafer (Holotrichia parallela) display preferential binding to biologically active host plant volatiles. Front. Physiol. 2018, 9, 769. [Google Scholar] [CrossRef]
- Wojtasek, H.; Picimbon, J.F.; Soares Leal, W. Identification and cloning of odorant binding proteins from the scarab beetle Phyllopertha diversa. Biochem. Biophys. Res. Commun. 1999, 263, 832–837. [Google Scholar] [CrossRef]
- Wojtasek, H.; Hansson, B.S.; Leal, W.S. Attracted or repelled?—A matter of two neurons, one pheromone binding protein, and a chiral center. Biochem. Biophys. Res. Commun. 1998, 250, 217–222. [Google Scholar] [CrossRef] [PubMed]
- Antony, B.; Johny, J.; Aldosari, S.A. Silencing the odorant binding protein RferOBP1768 reduces the strong preference of Palm Weevil for the major aggregation pheromone compound ferrugineol. Front. Physiol. 2018, 9, 252. [Google Scholar] [CrossRef]
- Zhang, R.; Wang, B.; Grossi, G.; Falabella, P.; Liu, Y.; Yan, S.; Lu, J.; Xi, J.; Wang, G. Molecular basis of alarm pheromone detection in aphids. Curr. Biol. 2017, 27, 55–61. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Guindon, S.; Dufayard, J.F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [PubMed]
- De Fouchier, A.; Walker, W.B.; Montagné, N.; Steiner, C.; Binyameen, M.; Schlyter, F.; Chertemps, T.; Maria, A.; François, M.C.; Monsempes, C.; et al. Functional evolution of Lepidoptera olfactory receptors revealed by deorphanization of a moth repertoire. Nat. Commun. 2017, 8, 15709. [Google Scholar] [CrossRef]
- Leitch, O.; Papanicolaou, A.; Lennard, C.; Kirkbride, K.P.; Anderson, A. Chemosensory genes identified in the antennal transcriptome of the blowfly Calliphora stygia. BMC Genomics 2015, 16, 255. [Google Scholar] [CrossRef]
- Croset, V.; Rytz, R.; Cummins, S.F.; Budd, A.; Brawand, D.; Kaessmann, H.; Gibson, T.J.; Benton, R. Ancient protostome origin of chemosensory ionotropic glutamate receptors and the evolution of insect taste and olfaction. PLoS Genet. 2010, 6, e1001064. [Google Scholar] [CrossRef]
- Slone, J.; Daniels, J.; Amrein, H. Sugar receptors in Drosophila. Curr. Biol. 2007, 17, 1809–1816. [Google Scholar] [CrossRef]
- Takada, T.; Sato, R.; Kikuta, S. A mannitol/sorbitol receptor stimulates dietary intake in Tribolium castaneum. PLoS ONE 2017, 12, e0186420. [Google Scholar] [CrossRef] [PubMed]
- Missbach, C.; Dweck, H.K.M.; Vogel, H.; Vilcinskas, A.; Stensmyr, M.C.; Hansson, B.S.; Grosse-Wilde, E. Evolution of insect olfactory receptors. eLife 2014, 2014, e02115. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, R.F.; Andersson, M.N. Olfactory genomics of the Coleoptera. In Insect Pheromone Biochemistry and Molecular Biology; Blomquist, G.J., Vogt, R.G., Eds.; Elsevier Academic Press: London, UK, 2021; pp. 547–590. [Google Scholar]
- Leroy, P.D.; Schillings, T.; Farmakidis, J.; Heuskin, S.; Lognay, G.; Verheggen, F.J.; Brostaux, Y.; Haubruge, E.; Francis, F. Testing semiochemicals from aphid, plant and conspecific: Attraction of Harmonia axyridis. Insect Sci. 2012, 19, 372–382. [Google Scholar] [CrossRef]
- Verheggen, F.J.; Fagel, Q.; Heuskin, S.; Lognay, G.; Francis, F.; Haubruge, E. Electrophysiological and behavioral responses of the multicolored Asian lady beetle, Harmonia axyridis Pallas, to sesquiterpene semiochemicals. J. Chem. Ecol. 2007, 33, 2148–2155. [Google Scholar] [CrossRef]
- Engsontia, P.; Sanderson, A.P.; Cobb, M.; Walden, K.K.O.; Robertson, H.M.; Brown, S. The red flour beetle’s large nose: An expanded odorant receptor gene family in Tribolium castaneum. Insect Biochem. Mol. Biol. 2008, 38, 387–397. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Deng, W.; Li, M.; Xiao, Y.; Li, J.; Teng, K.; Xiao, Z.; Li, X.; Zhou, Z.; Li, Y. Analysis of chemosensory genes in full and hungry adults of Arma chinensis (Pentatomidae) through antennal transcriptome. Front. Physiol. 2020, 11, 588291. [Google Scholar] [CrossRef] [PubMed]
- Gautier, M.; Yamaguchi, J.; Foucaud, J.; Loiseau, A.; Ausset, A.; Facon, B.; Gschloessl, B.; Lagnel, J.; Loire, E.; Parrinello, H.; et al. The genomic basis of color pattern polymorphism in the harlequin ladybird. Curr. Biol. 2018, 28, 3296–3302. [Google Scholar] [CrossRef]
- Schoville, S.D.; Chen, Y.H.; Andersson, M.N.; Benoit, J.B.; Bhandari, A.; Bowsher, J.H.; Brevik, K.; Cappelle, K.; Chen, M.J.M.; Childers, A.K.; et al. A model species for agricultural pest genomics: The genome of the Colorado potato beetle, Leptinotarsa decemlineata (Coleoptera: Chrysomelidae). Sci. Rep. 2018, 8, 1931. [Google Scholar] [CrossRef]
- Fassotte, B.; Fischer, C.; Durieux, D.; Lognay, G.; Haubruge, E.; Francis, F.; Verheggen, F.J. First evidence of a volatile sex pheromone in lady beetles. PLoS ONE 2014, 9, e115011. [Google Scholar] [CrossRef]
- Verheggen, F.; Cherif, A.; Martin, C. The production of sex pheromone in lady beetles is conditioned by presence of aphids and not by mating status. J. Chem. Ecol. 2020, 46, 590–596. [Google Scholar] [CrossRef]
- Hodek, I.; Evans, E.W. Food relationships. In Ecology and Behaviour of the Ladybird Beetles (Coccinellidae); Hodek, I., van Emden, H.F., Honek, A., Eds.; John Wiley & Sons: Chichester, UK, 2012; pp. 141–274. [Google Scholar]
- Mathews, C.R.; Brown, M.W.; Wäckers, F.L. Comparison of peach cultivars for provision of extrafloral nectar resources to Harmonia axyridis (Coleoptera: Coccinellidae). Environ. Entomol. 2016, 45, 649–657. [Google Scholar] [CrossRef]
- Sloggett, J.J.; Magro, A.; Verheggen, F.J.; Hemptinne, J.L.; Hutchison, W.D.; Riddick, E.W. The chemical ecology of Harmonia axyridis. BioControl 2011, 56, 643–661. [Google Scholar] [CrossRef]
- Růžička, Z. Perception for oviposition-deterring larval tracks in aphidophagous coccinellids Cycloneda limbifer and Ceratomegilla undecimnotata (Coleoptera: Coccinellidae). Eur. J. Entomol. 2003, 100, 345–350. [Google Scholar] [CrossRef]
- Fujii, S.; Yavuz, A.; Slone, J.; Jagge, C.; Song, X.; Amrein, H. Drosophila sugar receptors in sweet taste perception, olfaction, and internal nutrient sensing. Curr. Biol. 2015, 25, 621–627. [Google Scholar] [CrossRef]
- Guerenstein, P.G.; Hildebrand, J.G. Roles and effects of environmental carbon dioxide in insect life. Annu. Rev. Entomol. 2008, 53, 161–178. [Google Scholar] [CrossRef]
- Chen, F.J.; Wu, G.; Parajulee, M.N.; Ge, F. Impact of elevated CO2 on the third trophic level: A predator Harmonia axyridis and a parasitoid Aphidius picipes. Biocontrol Sci. Technol. 2007, 17, 313–324. [Google Scholar] [CrossRef]
- Wang, S.N.; Peng, Y.; Lu, Z.Y.; Dhiloo, K.H.; Zheng, Y.; Shan, S.; Li, R.J.; Zhang, Y.J.; Guo, Y.Y. Cloning and expression profile of ionotropic receptors in the parasitoid wasp Microplitis mediator (Hymenoptera: Braconidae). J. Insect Physiol. 2016, 90, 27–35. [Google Scholar] [CrossRef]
- Xia, D.; Shen, C.; Zhang, Y.; Tang, Q.; Wang, J. Identification and expression patterns of sensory neuron membrane protein genes from Sitophilus zeamais Motschulsky (Coleoptera: Curculionidae). Coleopt. Bull. 2019, 73, 735–745. [Google Scholar] [CrossRef]
- Zhao, Y.J.; Li, G.C.; Zhu, J.Y.; Liu, N.Y. Genome-based analysis reveals a novel SNMP group of the Coleoptera and chemosensory receptors in Rhaphuma horsfieldi. Genomics 2020, 112, 2713–2728. [Google Scholar] [CrossRef] [PubMed]
- Pregitzer, P.; Greschista, M.; Breer, H.; Krieger, J. The sensory neurone membrane protein SNMP1 contributes to the sensitivity of a pheromone detection system. Insect Mol. Biol. 2014, 23, 733–742. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.; Tal, S.H.; Smith, D.P. SNMP is a signaling component required for pheromone sensitivity in Drosophila. Proc. Natl. Acad. Sci. USA 2008, 105, 10996–11001. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rondoni, G.; Roman, A.; Meslin, C.; Montagné, N.; Conti, E.; Jacquin-Joly, E. Antennal Transcriptome Analysis and Identification of Candidate Chemosensory Genes of the Harlequin Ladybird Beetle, Harmonia axyridis (Pallas) (Coleoptera: Coccinellidae). Insects 2021, 12, 209. https://doi.org/10.3390/insects12030209
Rondoni G, Roman A, Meslin C, Montagné N, Conti E, Jacquin-Joly E. Antennal Transcriptome Analysis and Identification of Candidate Chemosensory Genes of the Harlequin Ladybird Beetle, Harmonia axyridis (Pallas) (Coleoptera: Coccinellidae). Insects. 2021; 12(3):209. https://doi.org/10.3390/insects12030209
Chicago/Turabian StyleRondoni, Gabriele, Alessandro Roman, Camille Meslin, Nicolas Montagné, Eric Conti, and Emmanuelle Jacquin-Joly. 2021. "Antennal Transcriptome Analysis and Identification of Candidate Chemosensory Genes of the Harlequin Ladybird Beetle, Harmonia axyridis (Pallas) (Coleoptera: Coccinellidae)" Insects 12, no. 3: 209. https://doi.org/10.3390/insects12030209
APA StyleRondoni, G., Roman, A., Meslin, C., Montagné, N., Conti, E., & Jacquin-Joly, E. (2021). Antennal Transcriptome Analysis and Identification of Candidate Chemosensory Genes of the Harlequin Ladybird Beetle, Harmonia axyridis (Pallas) (Coleoptera: Coccinellidae). Insects, 12(3), 209. https://doi.org/10.3390/insects12030209