KASP Genotyping as a Molecular Tool for Diagnosis of Cassava-Colonizing Bemisia tabaci


Abstract
1. Introduction
2. Materials and Methods
2.1. Whitefly NextRAD Sequences
2.2. Primer Design
2.3. Conventional PCR and KASP Assay
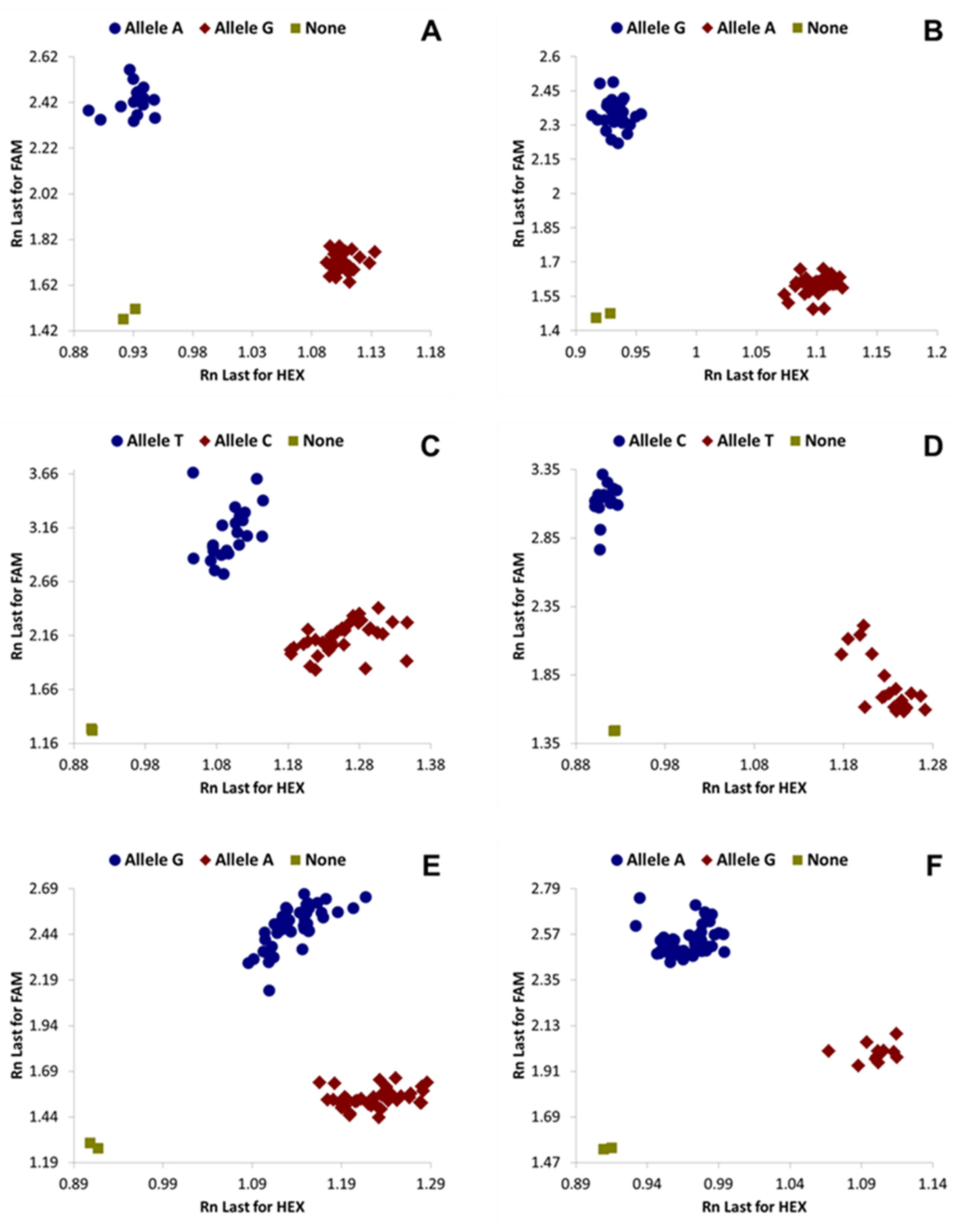
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gilbertson, R.L.; Batuman, O.; Webster, C.G.; Adkins, S. Role of the insect supervectors Bemisia tabaci and Frankliniella occidentalis in the emergence and global spread of plant viruses. Annu. Rev. Virol. 2015, 2, 67–93. [Google Scholar] [CrossRef] [PubMed]
- Storey, H.H.; Nichols, R.F.W. Studies on the mosaic of cassava. Ann. Appl. Biol. 1938, 25, 790–806. [Google Scholar] [CrossRef]
- Maruthi, M.N.; Hillocks, R.J.; Mtunda, K.; Raya, M.D.; Muhanna, M.; Kiozia, H.; Rekha, A.R.; Colvin, J.; Thresh, J.M. Transmission of Cassava brown streak virus by Bemisia tabaci (Gennadius). J. Phytopathol. 2005, 153, 307–312. [Google Scholar] [CrossRef]
- Legg, J.P.; Jeremiah, S.C.; Obiero, H.M.; Maruthi, M.N.; Ndyetabula, I.; Okao-Okuja, G.; Bouwmeester, H.; Bigirimana, S.; Tata-Hangy, W.; Gashaka, G.; et al. Comparing the regional epidemiology of the cassava mosaic and cassava brown streak virus pandemics in Africa. Virus Res. 2011, 159, 161–170. [Google Scholar] [CrossRef] [PubMed]
- De Barro, P.J. The Bemisia species complex: Questions to guide future research. J. Integr. Agric. 2012, 11, 187–196. [Google Scholar] [CrossRef]
- Dinsdale, A.; Cook, L.; Riginos, C.; Buckley, Y.M.; De Barro, P. Refined global analysis of Bemisia tabaci (Hemiptera: Sternorrhyncha: Aleyrodoidea: Aleyrodidae) mitochondrial cytochrome oxidase I to identify species level genetic boundaries. Ann. Entomol. Soc. Am. 2010, 103, 196–208. [Google Scholar] [CrossRef]
- Tay, W.T.; Evans, G.A.; Boykin, L.M.; De Barro, P.J. Will the real Bemisia tabaci please stand up? PLoS ONE 2012, 7, e50550. [Google Scholar] [CrossRef]
- Brown, J.K.; Coats, S.A.; Bedford, I.D.; Markham, P.G.; Bird, J.; Frohlich, D.R. Characterization and distribution of esterase electromorphs in the whitefly, Bemisia tabaci (Genn.) (Homoptera: Aleyrodidae). Biochem. Genet. 1995, 33, 205–214. [Google Scholar]
- Gill, R.J. The morphology of whiteflies. In Whiteflies: Their Bionomics, Pest Status and Management; Gerling, D., Ed.; Intercept: Andover, UK, 1990; pp. 13–46. [Google Scholar]
- Mound, L.A. Host-correlated variation in Bemisia tabaci (Gennadius) (Homoptera: Aleyrodidae). Proc. R. Entomol. Soc. 1963, 38, 171–180. [Google Scholar] [CrossRef]
- Rosell, R.C.; Bedford, I.D.; Frohlich, D.R.; Gill, R.J.; Brown, J.K.; Markham, P.G. Analysis of morphological variation in distinct populations of Bemisia tabaci (Homoptera: Aleyrodidae). Ann. Entomol. Soc. Am. 1997, 90, 575–589. [Google Scholar] [CrossRef]
- Hadjistylli, M.; Brown, J.K.; Roderick, G.K. Tools and recent progress in studying gene flow and population genetics of the Bemisia tabaci sibling species group. In Bemisia: Bionomics and Management of a Global Pest; Stansly, P.A., Naranjo, S.E., Eds.; Springer: Dordrecht, The Netherlands, 2010; pp. 69–103. [Google Scholar]
- Brown, J.K. Phylogenetic biology of the Bemisia tabaci sibling species group. In Bemisia: Bionomics and Management of a Global Pest; Stansly, P.A., Naranjo, S.E., Eds.; Springer: Dordrecht, The Netherlands, 2010; pp. 31–67. [Google Scholar]
- Hadjistylli, M.; Roderick, G.K.; Brown, J.K. Global population structure of a worldwide pest and virus vector: Genetic diversity and population history of the Bemisia tabaci sibling species group. PLoS ONE 2016, 11, e0165105. [Google Scholar] [CrossRef] [PubMed]
- Ballard, J.W.; Whitlock, M.C. The incomplete natural history of mitochondria. Mol. Ecol. 2004, 13, 729–744. [Google Scholar] [CrossRef]
- Zhang, D.X.; Hewitt, G.M. Nuclear integrations: Challenges for mitochondrial DNA markers. Trends Ecol. Evol. 1996, 11, 247–251. [Google Scholar] [CrossRef]
- Tay, W.T.; Elfekih, S.; Court, L.N.; Gordon, K.H.; Delatte, H.; De Barro, P.J. The trouble with MEAM2: Implications of pseudogenes on species delimitation in the globally invasive Bemisia tabaci (Hemiptera: Aleyrodidae) cryptic species complex. Genome Biol. Evol. 2017, 9, 2732–2738. [Google Scholar] [CrossRef] [PubMed][Green Version]
- De Moya, R.S.; Brown, J.K.; Sweet, A.D.; Walden, K.K.; Paredes-Montero, J.R.; Waterhouse, R.M.; Johnson, K.P. Nuclear orthologs derived from whole genome sequencing indicate cryptic diversity in the Bemisia tabaci (Insecta: Aleyrodidae) complex of whiteflies. Diversity 2019, 11, 151. [Google Scholar] [CrossRef]
- Morin, P.A.; Luikart, G.; Wayne, R.K. SNPs in ecology, evolution and conservation. Trends Ecol. Evol. 2004, 19, 208–216. [Google Scholar] [CrossRef]
- Helyar, S.J.; Hemmer-Hansen, J.; Bekkevold, D.; Taylor, M.I.; Ogden, R.; Limborg, M.T.; Cariani, A.; Maes, G.E.; Diopere, E.; Carvalho, G.R.; et al. Application of SNPs for population genetics of nonmodel organisms: New opportunities and challenges. Mol. Ecol. Resour. 2011, 11, 123–136. [Google Scholar] [CrossRef]
- Bourgeois, S.; Senn, H.; Kaden, J.; Taggart, J.B.; Ogden, R.; Jeffery, K.J.; Bunnefeld, N.; Abernethy, K.; McEwing, R. Single-nucleotide polymorphism discovery and panel characterization in the African forest elephant. Ecol. Evol. 2018, 8, 2207–2217. [Google Scholar] [CrossRef]
- Van Tassell, C.P.; Smith, T.P.; Matukumalli, L.K.; Taylor, J.F.; Schnabel, R.D.; Lawley, C.T.; Haudenschild, C.D.; Moore, S.S.; Warren, W.C.; Sonstegard, T.S. SNP discovery and allele frequency estimation by deep sequencing of reduced representation libraries. Nat. Methods 2008, 5, 247–252. [Google Scholar] [CrossRef]
- Russello, M.A.; Waterhouse, M.D.; Etter, P.D.; Johnson, E.A. From promise to practice: Pairing non-invasive sampling with genomics in conservation. PeerJ 2015, 3, e1106. [Google Scholar] [CrossRef]
- Wosula, E.N.; Chen, W.; Fei, Z.; Legg, J.P. Unravelling the genetic diversity among cassava Bemisia tabaci whiteflies using NextRAD sequencing. Genome Biol. Evol. 2017, 9, 2958–2973. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Wosula, E.N.; Hasegawa, D.K.; Casinga, C.; Shirima, R.R.; Fiaboe, K.K.M.; Hanna, R.; Fosto, A.; Goergen, G.; Tamò, M.; et al. Genome of the African cassava whitefly Bemisia tabaci and distribution and genetic diversity of cassava-colonizing whiteflies in Africa. Insect Biochem. Mol. Biol. 2019, 110, 112–120. [Google Scholar] [CrossRef] [PubMed]
- Emerson, K.J.; Conn, J.E.; Bergo, E.S.; Randel, M.A.; Sallum, M.A.M. Brazilian Anopheles darlingi Root (Diptera: Culicidae) clusters by major biogeographical region. PLoS ONE 2015, 10, e0130773. [Google Scholar] [CrossRef] [PubMed]
- Fu, Z.; Epstein, B.; Kelley, J.L.; Zheng, Q.; Bergland, A.O.; Carrillo, C.I.C.; Jensen, A.S.; Dahan, J.; Karasev, A.V.; Snyder, W.E. Using NextRAD sequencing to infer movement of herbivores among host plants. PLoS ONE 2017, 12, e0177742. [Google Scholar] [CrossRef] [PubMed]
- Otim-Nape, G.W.; Bua, A.; Baguma, Y. Accelerating the transfer of improved production technologies: Controlling African cassava mosaic virus disease epidemics in Uganda. Afr. Crop Sci. J. 1994, 2, 479–495. [Google Scholar]
- Burban, C.L.; Fishpool, L.D.; Fauquet, C.; Fargette, D.; Thouvenel, J.C. Host-associated biotypes within West African populations of the whitefly Bemisia tabaci (Genn.), (Hom., Aleyrodidae). J. Appl. Entomol. 1992, 113, 416–423. [Google Scholar] [CrossRef]
- Legg, J.P.; Gibson, R.W.; Otim-Nape, G.W. Genetic polymorphism amongst Ugandan populations of Bemisia tabaci (Gennadius) (Homoptera: Aleyrodidae), vector of African cassava mosaic geminivirus. Trop. Sci. 1991, 34, 73–81. [Google Scholar]
- Legg, J.P. Host-associated strains within Ugandan populations of the whitefly Bemisia tabaci (Genn.), (Hom., Aleyrodidae). J. Appl. Entomol. 1996, 120, 523–527. [Google Scholar] [CrossRef]
- Legg, J.P.; French, R.; Rogan, D.; Okao-Okuja, G.; Brown, J.K. A distinct Bemisia tabaci (Gennadius) (Hemiptera: Sternorrhyncha: Aleyrodidae) genotype cluster is associated with the epidemic of severe cassava mosaic virus disease in Uganda. Mol. Ecol. 2002, 11, 1219–1229. [Google Scholar] [CrossRef]
- Berry, S.D.; Fondong, V.N.; Rey, C.; Rogan, D.; Fauquet, C.M.; Brown, J.K. Molecular evidence for five distinct Bemisia tabaci (Homoptera: Aleyrodidae) geographic haplotypes associated with cassava plants in sub-Saharan Africa. Ann. Entomol. Soc. Am. 2004, 97, 852–859. [Google Scholar] [CrossRef]
- Legg, J.P.; Sseruwagi, P.; Boniface, S.; Okao-Okuja, G.; Shirima, R.; Bigirimana, S.; Gashaka, G.; Herrmann, H.W.; Jeremiah, S.; Obiero, H.; et al. Spatio-temporal patterns of genetic change amongst populations of cassava Bemisia tabaci whiteflies driving virus pandemics in East and Central Africa. Virus Res. 2014, 186, 61–75. [Google Scholar] [CrossRef]
- Ghosh, S.; Bouvaine, S.; Maruthi, M.N. Prevalence and genetic diversity of endosymbiotic bacteria infecting cassava whiteflies in Africa. BMC Microbiol. 2015, 15, 93. [Google Scholar] [CrossRef]
- Semagn, K.; Babu, R.; Hearne, S.; Olsen, M. Single nucleotide polymorphism genotyping using Kompetitive Allele Specific PCR (KASP): Overview of the technology and its application in crop improvement. Mol. Breed. 2014, 33, 1–14. [Google Scholar] [CrossRef]
- Bradbury, P.J.; Zhang, Z.; Kroon, D.E.; Casstevens, T.M.; Ramdoss, Y.; Buckler, E.S. TASSEL: Software for association mapping of complex traits in diverse samples. Bioinformatics 2007, 23, 2633–2635. [Google Scholar] [CrossRef] [PubMed]
- Pelak, K.; Shianna, K.V.; Ge, D.; Maia, J.M.; Zhu, M.; Smith, J.P.; Cirulli, E.T.; Fellay, J.; Dickson, S.P.; Gumbs, C.E.; et al. The characterization of twenty sequenced human genomes. PLoS Genet. 2010, 6, e1001111. [Google Scholar] [CrossRef] [PubMed]
- Gautier, M.; Gharbi, K.; Cezard, T.; Foucaud, J.; Kerdelhué, C.; Pudlo, P.; Cornuet, J.M.; Estoup, A. The effect of RAD allele dropout on the estimation of genetic variation within and between populations. Mol. Ecol. 2013, 22, 3165–3178. [Google Scholar] [CrossRef]
- Senn, H.; Ogden, R.O.; Cezard, T.; Gharbi, K.; Iqbal, Z.; Johnson, E.; Kamps-Hughes, N.; Rosell, F.; McEwing, R. Reference-free SNP discovery for the Eurasian beaver from restriction site-associated DNA paired-end data. Mol. Ecol. 2013, 22, 3141–3150. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
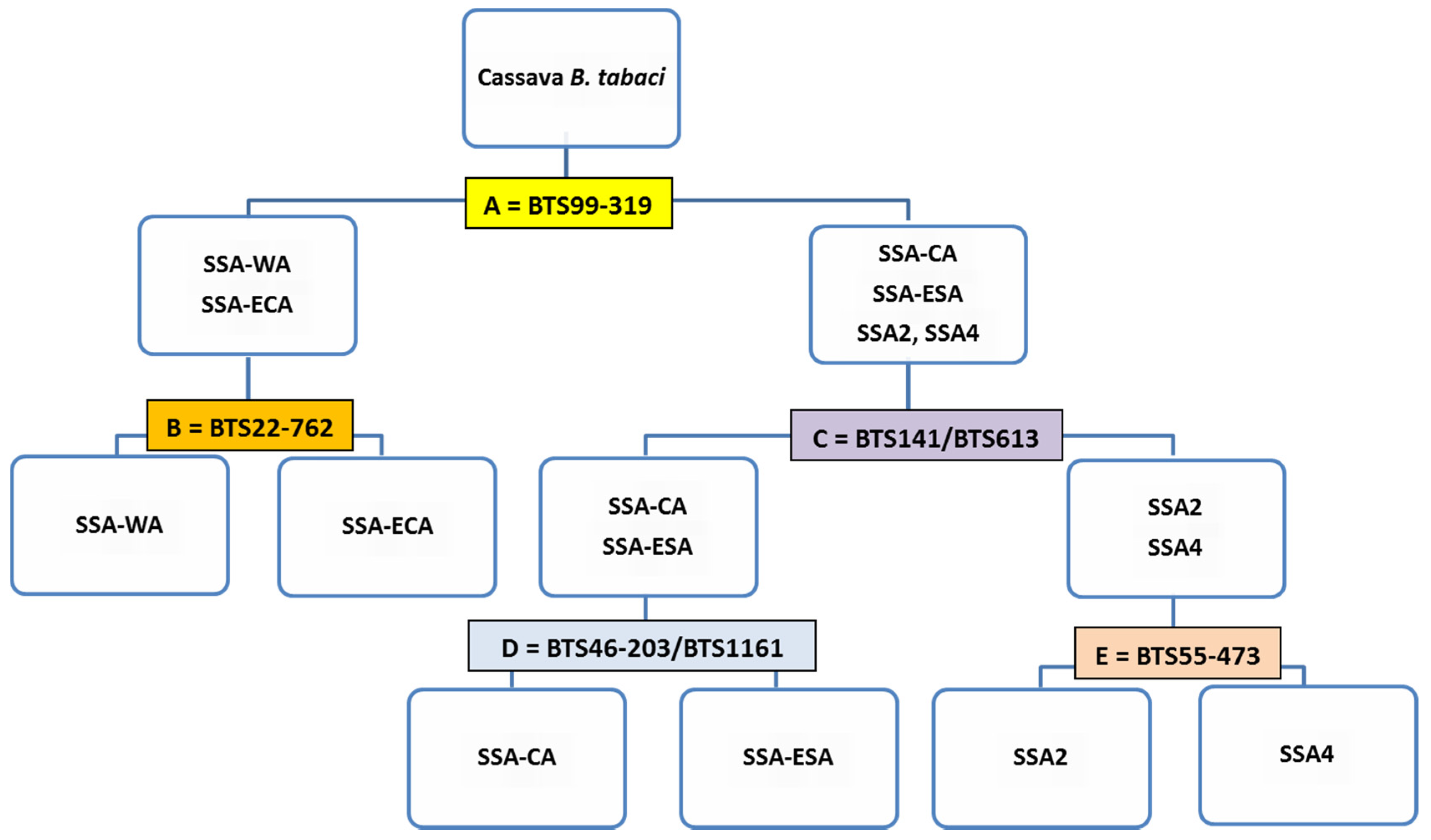
| KASP Primers | Position in Genome | SSA-ECA | SSA-WA | SSA-ESA | SSA-CA | SSA2 | SSA4 |
|---|---|---|---|---|---|---|---|
| BTS99-319 | Intergenic region | A:A | A:A | G:G | G:G | G:G | G:G |
| BTS22-762 | Ssa12858; exon | A:A | G:G | - | - | - | - |
| BTS141 | Intergenic region | C:C | C:C | C:C | C:C | T:T | T:T |
| BTS55-473 | Intergenic region | C:C | C:C | C:C | C:C | T:T | C:C |
| BTS613 | Intergenic region | A:A | A:A | G:G | G:G | A:A | A:A |
| BTS46-203 | Intron of Ssa00724 | - | - | A:A | G:G | - | - |
| BTS1161 | Intron of Ssa00849 | - | - | C:C | A:A | - | - |
| Primer | Sequence | Band (bp) | Accession | Range |
|---|---|---|---|---|
| BTS99-319F | TTTCTGGAGGTATGATGTT | 940 | PGTP01000606.1 | 849,024–849,984 |
| BTS99-319R | GTTGGCTTGTTTTTCTTTG | |||
| BTS22-762F | CAAACGAACACAACCGCAA | 880 | PGTP01001647.1 | 62,324–63,244 |
| BTS22-762R | CAGGGACGTACACAAAATAA | |||
| BTS141F | TCCTCAGCAGTGTCTTTT | 1150 | PGTP01000392.1 | 896,170–897,370 |
| BTS141R | TCTACGTTGTGTTGTCGG | |||
| BTS55-473F | ACCCCACCAAATATCTCAC | 815 | PGTP01143321.1 | 559,651–560,491 |
| BTS55-473R | GGCATTCCAGCAAAATATACA | |||
| BTS613F | CATTCCGCTTTCCATCCTC | 730 | PGTP01000317.1 | 28,760–29,515 |
| BTS613R | CCTTCTCTTGTCGAACAT | |||
| BTS46-203F | CGAGGGCTAAAGAATAATAC | 390 | PGTP01000379.1 | 1,505,366–1,505,786 |
| BTS46-203R | TTCAGAACGAATGAGAAGG | |||
| BTS1161F | TTATTTTCGGTGGTGCGTC | 500 | PGTP01001427.1 | 371,644–372,644 |
| BTS1161R | GATGATGAGGGTAGAGTT |
| ID | Primer AlleleX | Primer AlleleY | Primer Common | Size bp |
|---|---|---|---|---|
| BTS99-319 | CTCAAATTTAAAATACGATTTCAATTACCATT | CTCAAATTTAAAATACGATTTCAATTACCATC | GCTGCATATTATACCGCATGAAAGCTAAA | 40 |
| BTS22-762 | CAGTCAATTAAAAGACGTCTCGCTAA | CAGTCAATTAAAAGACGTCTCGCTAG | GTCGCTGTCTTGTTTTCCCTCCAT | 50 |
| BTS141 | TACTATTTCTAGCAAAGCGAATTTAAATCATA | CTATTTCTAGCAAAGCGAATTTAAATCATG | GGAGTGCTATAAAGCGACCTATATGTAT | 36 |
| BTS55-473 | CCGCACAGGAGACCCAAGTC | ACCGCACAGGAGACCCAAGTT | TAATAAGCCCGACATGCCGCTCTTT | 38 |
| BTS613 | GGTAGAGCGGCGCTTGGTC | ATGGTAGAGCGGCGCTTGGTT | ACTTCGGCTTTGAACTTCCCGCAAA | 30 |
| BTS46-203 | GGTGCATCGTATCGCATCTCTGA | GTGCATCGTATCGCATCTCTGG | CATATAACTACGCGCAACGCAACGTA | 62 |
| BTS1161 | AAGTCTTGCTGCTATGGCTTAGTTC | AAAGTCTTGCTGCTATGGCTTAGTTA | CCCCATGTAGAGCTCCAGGTAAAAT | 60 |
| KASP Primers | NextRAD Identity | KASP Diagnosis | Accuracy | Mismatch |
|---|---|---|---|---|
| (i) BTS99-319 | A:A (SSA-ECA/SSA-WA) (73) | A:A (74) | 99.3% | 0.7% (Malawi) |
| G:G (SSA-ESA/SSA-CA/SSA2/SSA4) (79) | G:G (78) | |||
| (ii) BTS22-762 | G:G (SSA-WA) (41) | G:G (42) | 98.6% | 1.4% (Kenya) |
| A:A (SSA-ECA) (32) | A:A (31) | |||
| (iii) BTS141 | T:T (SSA2/SSA4) (35) | T:T (34) C:T (1) | 99.3% | 0.7% (DRC) |
| C:C (SSA-ECA/SSA-WA/SSA-CA/SSA-ESA) (117) | C:C (117) | |||
| (iv) BTS55-473 | T:T (SSA2) (26) | T:T (26) | 100% | 0% |
| C:C (SSA4) (8) | C:C (8) | 100% | 0% | |
| (v) BTS613 | G:G/G:A (SSA-ESA/SSA-CA) (45) | G:G (45) | 100% | 0% |
| A:A (SSA-ECA/SSA-WA/SSA2/SSA4) (107) | A:A (107) | 100% | 0% | |
| (vi) BTS46-203 | A:A (SSA-ESA) (41) | A:A (39) A:G (2) | 95.1% | 4.9% (Kenya, Tanzania) |
| G:G (SSA-CA) (4) | G:G (2) A:G (2) | 50% | 50% (DRC) | |
| (vii) BTS1161 | A:A (SSA-ESA) (41) | C:C (33) C:A (8) | 80.5% | 19.5% (Malawi, Mozambique) |
| G:G (SSA-CA) (4) | A:A (4) | 100% | 0% |
| NextRAD Identity | |||||||
|---|---|---|---|---|---|---|---|
| SSA-ECA | SSA-WA | SSA-CA | SSA-ESA | SSA2 | SSA4 | ||
| KASP Diagnosis | SSA-ECA | 31 a | |||||
| SSA-WA | 1 b | 41 a | |||||
| SSA-CA | 4 a | ||||||
| SSA-ESA | 41 a | ||||||
| SSA2 | 26 a | ||||||
| SSA4 | 8 a | ||||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wosula, E.N.; Chen, W.; Amour, M.; Fei, Z.; Legg, J.P. KASP Genotyping as a Molecular Tool for Diagnosis of Cassava-Colonizing Bemisia tabaci. Insects 2020, 11, 305. https://doi.org/10.3390/insects11050305
Wosula EN, Chen W, Amour M, Fei Z, Legg JP. KASP Genotyping as a Molecular Tool for Diagnosis of Cassava-Colonizing Bemisia tabaci. Insects. 2020; 11(5):305. https://doi.org/10.3390/insects11050305
Chicago/Turabian StyleWosula, Everlyne N., Wenbo Chen, Massoud Amour, Zhangjun Fei, and James P. Legg. 2020. "KASP Genotyping as a Molecular Tool for Diagnosis of Cassava-Colonizing Bemisia tabaci" Insects 11, no. 5: 305. https://doi.org/10.3390/insects11050305
APA StyleWosula, E. N., Chen, W., Amour, M., Fei, Z., & Legg, J. P. (2020). KASP Genotyping as a Molecular Tool for Diagnosis of Cassava-Colonizing Bemisia tabaci. Insects, 11(5), 305. https://doi.org/10.3390/insects11050305