Behavioral and Electrophysiological Responses of the Fringed Larder Beetle Dermestes frischii to the Smell of a Cadaver at Different Decomposition Stages




{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Rearing
2.2. Rat Decomposition
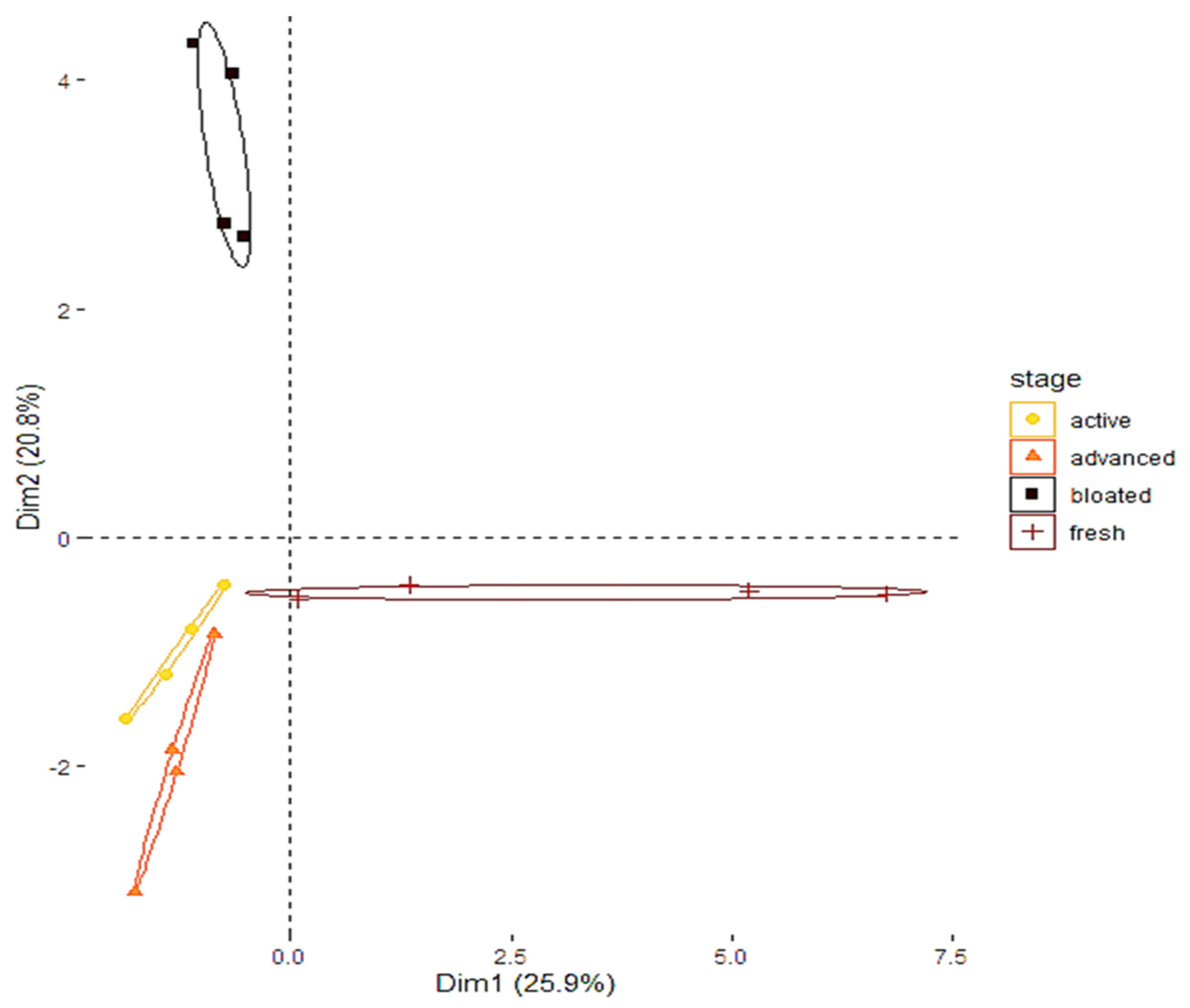
2.3. Odor Collection and Analysis
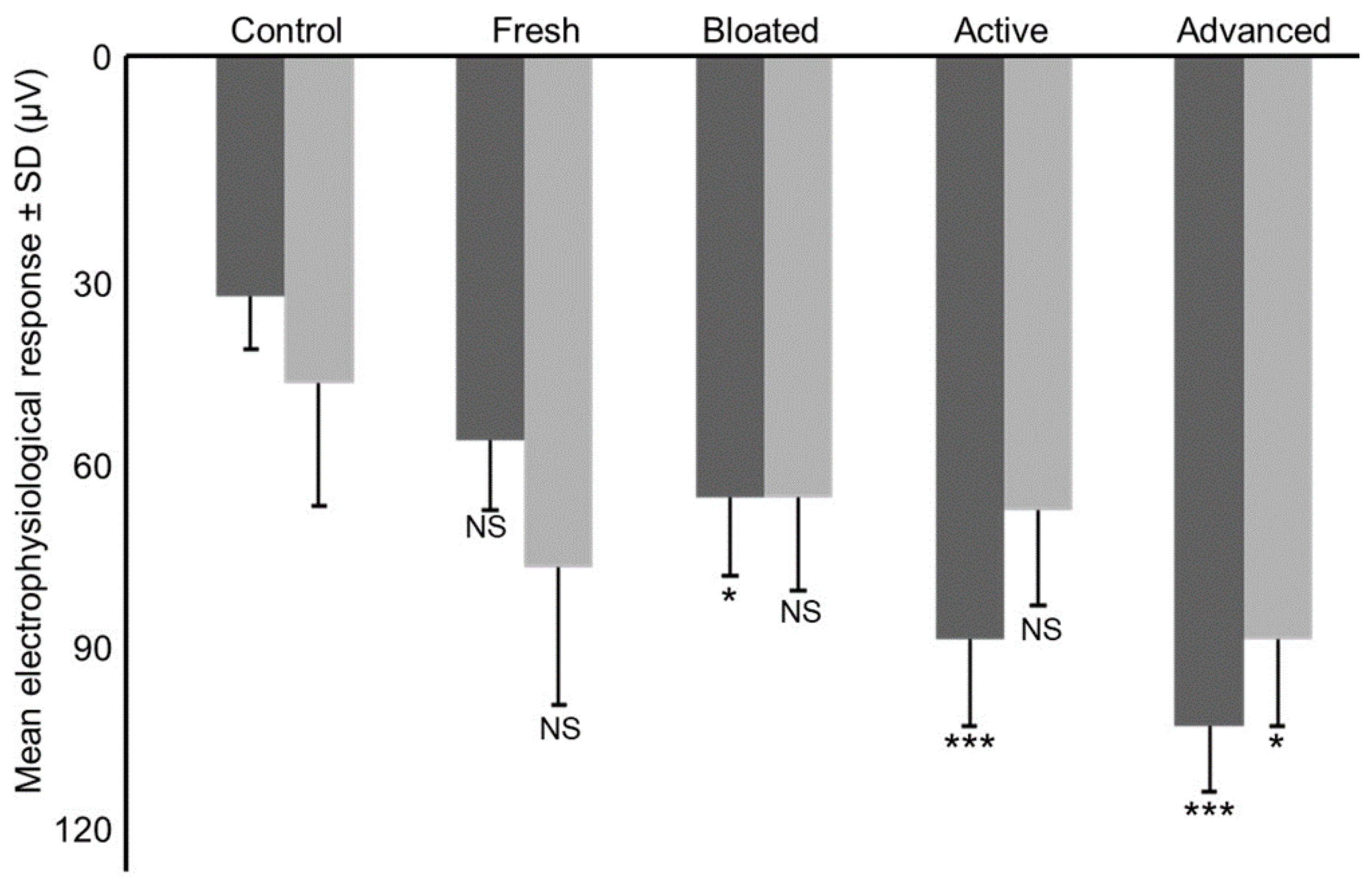
2.4. Electroantennography
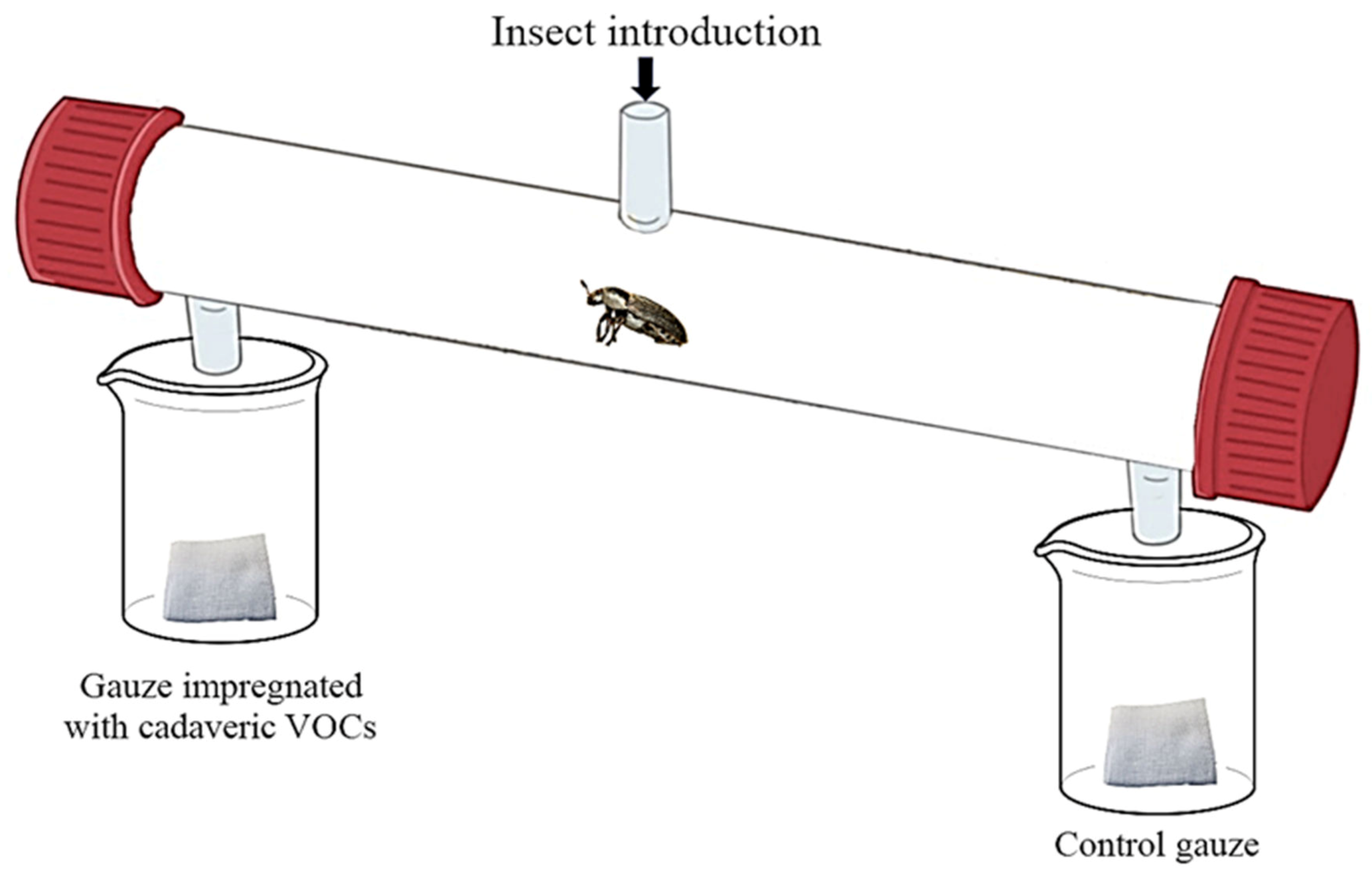
2.5. Behavioral Assays
2.6. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Dent, B.; Forbes, S.; Stuart, B. Review of human decomposition processes in soil. Environ. Geol. 2004, 45, 576–585. [Google Scholar] [CrossRef]
- Statheropoulos, M.; Agapiou, A.; Spiliopoulou, C.; Pallis, G.; Sianos, E. Environmental aspects of VOCs evolved in the early stages of human decomposition. Sci. Total Environ. 2007, 385, 221–227. [Google Scholar] [CrossRef]
- Janaway, R.C.; Percival, S.; Wilson, A. Decomposition of human remains. In Microbiology and Aging: Clinical Manifestations, 1st ed.; Perceval, S.L., Ed.; Springer Nature: Basel, Switzerland, 2009; pp. 313–334. [Google Scholar] [CrossRef]
- Martin, C.; Verheggen, F. Odour profile of human corpses: A review. Forensic Chem. 2018, 10, 27–36. [Google Scholar] [CrossRef]
- Adams, V. Dying: What happens to the body after death. In Dying and Death in Oncology, 1st ed.; Berk, L., Ed.; Springer Nature: Cham, Switzerland, 2017; pp. 23–30. [Google Scholar] [CrossRef]
- Payne, J. A Summer Carrion Study of the Baby Pig Sus Scrofa Linnaeus. Ecology 1965, 46, 592–602. [Google Scholar] [CrossRef]
- Vass, A. Beyond the grave—Understanding human decomposition. Microbiol. Today 2001, 28, 190–193. [Google Scholar]
- Amendt, J.; Krettek, R.; Zehner, R. Forensic entomology. Naturwissenschaftenten 2004, 91, 51–65. [Google Scholar] [CrossRef] [PubMed]
- Grassberger, M.; Frank, C. Initial study of arthropod succession on pig carrion in a central European urban habitat. J. Med. Entomol. 2004, 41, 511–523. [Google Scholar] [CrossRef] [PubMed]
- Dekeirsschieter, J.; Verheggen, F.; Gohy, M.; Hubrecht, F.; Bourguignon, L.; Lognay, G.; Haubruge, E. Cadaveric volatile organic compounds released by decaying pig carcasses (Sus domesticus L.) in different biotopes. Forensic Sci. Int. 2009, 189, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Cernosek, T.; Eckert, K.; Carter, D.; Perrault, K. Volatile organic compound profiling from postmortem microbes using gas chromatography-mass spectrometry. J. Forensic Sci. 2019, 65, 134–143. [Google Scholar] [CrossRef]
- Martin, C.; Vanderplanck, M.; Boullis, A.; Francis, F.; Haubruge, E.; Verheggen, F. Impact of necrophagous insects on the emission of volatile organic compounds released during the decaying process. Entomol. Gen. 2019, 39, 19–31. [Google Scholar] [CrossRef]
- Forbes, S.; Perrault, K.; Stefanuto, P.; Nizio, K.; Focant, J. Comparison of the decomposition VOC profile during winter and summer in a moist, mid-latitude (Cfb) climate. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Forbes, S.; Perrault, K. Decomposition odour profiling in the air and soil surrounding vertebrate carrion. PLoS ONE 2014, 9. [Google Scholar] [CrossRef]
- Verheggen, F.; Perrault, K.; Megido, R.; Dubois, L.; Francis, F.; Haubruge, E.; Forbes, S.L.; Focant, J.-F.; Stefanuto, P.-H. The odor of death: An overview of current knowledge on characterization and applications. Bioscience 2017, 67, 600–613. [Google Scholar] [CrossRef]
- Anderson, G. Insects succession on carrion and its relationship to determining time of death. In Forensic Entomology: The Utility of Arthropods in Legal Investigation, 1st ed.; Castner, E., Byrd, J., Eds.; CRC Press: Boca Raton, FL, USA, 2000; pp. 143–175. [Google Scholar] [CrossRef]
- Brodie, B.; Gries, R.; Martins, A.; Vanlaerhoven, S.; Gries, G. Bimodal cue complex signifies suitable oviposition sites to gravid females of the common green bottle fly. Entomol. Exp. Appl. 2014, 153, 114–127. [Google Scholar] [CrossRef]
- Zhu, J.; Chaudhury, M.; Tangtrakulwanich, K.; Skoda, S. Identification of oviposition attractants of the secondary screwworm, Cochliomyia macellaria (F.) released from rotten chicken liver. J. Chem. Ecol. 2013, 39, 1407–1414. [Google Scholar] [CrossRef]
- Frederickx, C.; Dekeirsschieter, J.; Verheggen, F.; Haubruge, E. Responses of Lucilia sericata Meigen (Diptera: Calliphoridae) to cadaveric volatile organic compounds. J. Forensic Sci. 2012, 57, 386–390. [Google Scholar] [CrossRef] [PubMed]
- Nilssen, A.; Tømmerås, B.; Schmid, R.; Evensen, S. Dimethyl trisulphide is a strong attractant for some calliphorids and a muscid but not for the reindeer oestrids Hypoderma tarandi and Cephenemyia trompe. Entomol. Exp. Appl. 1996, 79, 211–218. [Google Scholar] [CrossRef]
- Dekeirsschieter, J.; Verheggen, F.; Haubruge, E.; Brostaux, Y. Carrion beetles visiting pig carcasses during early spring in urban, forest and agricultural biotopes of Western Europe. J. Insect Sci. 2011, 11, 1–13. [Google Scholar] [CrossRef]
- Charabidzé, D.; Colard, T.; Vincent, B.; Pasquerault, T.; Hedouin, V. Involvement of larder beetles (Coleoptera: Dermestidae) on human cadavers: A review of 81 forensic cases. Int. J. Legal Med. 2014, 128, 1021–1030. [Google Scholar] [CrossRef]
- Magni, P.; Voss, S.; Testi, R.; Borrini, M.; Dadour, I. A biological and procedural review of forensically significant Dermestes species (Coleoptera: Dermestidae). J. Med. Entomol. 2015, 52, 755–769. [Google Scholar] [CrossRef]
- Martin-Vega, D.; Diaz-Aranda, L.; Baz, A.; Cifrian, B. Effect of temperature on the survival and development of three forensically relevant Dermestes species (Coleoptera: Dermestidae). J. Med. Entomol. 2017, 54, 1140–1150. [Google Scholar] [CrossRef]
- Kulshrestha, P.; Satpathy, D. Use of beetles in forensic entomology. Forensic Sci. Int. 2001, 120, 15–17. [Google Scholar] [CrossRef]
- Boeckh, J.; Kaissling, K.E.; Schneider, D. Insect olfactory receptors. Cold Spring Harb. Symp. Quant. Biol. 1965, 30, 263–280. [Google Scholar] [CrossRef]
- Kalinová, B.; Podskalská, H.; Růžička, J.; Hoskovec, M. Irresistible bouquet of death-how are burying beetles (Coleoptera: Silphidae: Nicrophorus) attracted by carcasses. Naturwissenschaften 2009, 96, 889–899. [Google Scholar] [CrossRef]
- Podskalská, H.; Růžička, J.; Hoskovec, M.; Šálek, M. Use of infochemicals to attract carrion beetles into pitfall traps. Entomol. Exp. Appl. 2009, 132, 59–64. [Google Scholar] [CrossRef]
- Dekeirsschieter, J.; Frederickx, C.; Lognay, G.; Brostaux, Y.; Verheggen, F.; Haubruge, E. Electrophysiological and behavioral responses of Thanatophilus sinuatus Fabricius (Coleoptera: Silphidae) to selected cadaveric volatile organic compounds. J. Forensic Sci. 2013, 58, 917–923. [Google Scholar] [CrossRef] [PubMed]
- Von Hoermann, C.; Ruther, J.; Reibe, S.; Madea, B.; Ayasse, M. The importance of carcass volatiles as attractants for the hide beetle Dermestes maculatus (De Geer). Forensic Sci. Int. 2011, 212, 173–179. [Google Scholar] [CrossRef]
- Zanetti, N.; Visciarelli, E.; Centeno, N. Trophic roles of scavenger beetles in relation to decomposition stages and seasons. Rev. Bras. Entomol. 2015, 59, 132–137. [Google Scholar] [CrossRef]
- Kumara, T.; Abu Hassan, A.; Che Salmah, M.; Bhupinder, S. The infestation of Dermestes ater (De Geer) on a human corpse in Malaysia. Trop. Biomed. 2009, 26, 73–79. [Google Scholar] [PubMed]
- Rosenbaum, F.; Devigne, C.; Charabidzé, D. Ecologie des Dermestidae, une famille de Coléoptères nécrophages associée aux cadavres squelettisés. Ann. Soc. Entomol. Fr. 2015, 51, 294–302. [Google Scholar] [CrossRef]
- Martin, C.; Verheggen, F. Behavioural response of Lucilia sericata to a decaying body infested by necrophagous insects. Physiol. Entomol. 2018, 43, 188–195. [Google Scholar] [CrossRef]
- Degreeff, L.; Furton, K. Collection and identification of human remains volatiles by non-contact, dynamic airflow sampling and SPME-GC/MS using various sorbent materials. Anal. Bioanal. Chem. 2011, 401, 1295–1307. [Google Scholar] [CrossRef]
- DeGreeff, L.; Weakley-Jones, B.; Furton, K. Creation of training aids for human remains detection canines utilizing a non-contact, dynamic airflow volatile concentration technique. Forensic Sci. Int. 2012, 217, 32–38. [Google Scholar] [CrossRef] [PubMed]
- Verheggen, F.; Arnaud, L.; Bartram, S.; Gohy, M.; Haubruge, E. Aphid and plant volatiles induce oviposition in an aphidophagous hoverfly. J. Chem. Ecol. 2008, 34, 301–307. [Google Scholar] [CrossRef]
- Verheggen, F.; Fagel, Q.; Heuskin, S.; Lognay, G.; Francis, F.; Haubruge, E. Electrophysiological and behavioral responses of the multicolored Asian lady beetle, Harmonia axyridis pallas, to sesquiterpene semiochemicals. J. Chem. Ecol. 2007, 33, 2148–2155. [Google Scholar] [CrossRef]
- De Backer, L.; Bawin, T.; Schott, M.; Gillard, L.; Markó, I.; Verheggen, F. Betraying its presence: Identification of the chemical signal released by Tuta absoluta-infested tomato plants that guide generalist predators toward their prey. Arthropod Plant Interact. 2016, 11, 111–120. [Google Scholar] [CrossRef]
- Barsics, F.; Latine, R.; Gfeller, A.; Laloux, M.; Lognay, G.; Whatelet, J.; Haubruge, E.; Verheggen, F. Do root-emitted volatile organic compounds attract wireworms? Commun. Agric. Appl. Biol. Sci. 2012, 77, 561–565. [Google Scholar]
- Gfeller, A.; Laloux, M.; Barsics, F.; Kati, D.; Haubruge, E.; Du Jardin, P.; Verheggen, F.; Lognay, G.; Wathelet, J.-P.; Fauconnier, M.-L. Characterization of volatile organic compounds emitted by barley (Hordeum vulgare L.) roots and their attractiveness to wireworms. J. Chem. Ecol. 2013, 39, 1129–1139. [Google Scholar] [CrossRef]
- Kuznetsova, A.; Brockhoff, P.; Christensen, R. lmerTest package: Tests in linear mixed effects models. J. Stat. Softw. 2017, 82, 1–26. [Google Scholar] [CrossRef]
- Campobasso, C.; Di Vella, G.; Introna, F. Factors affecting decomposition and Diptera colonization. Forensic Sci. Int. 2001, 120, 18–27. [Google Scholar] [CrossRef]
- Sharanowski, B.; Walker, E.; Anderson, G. Insect succession and decomposition patterns on shaded and sunlit carrion in Saskatchewan in three different seasons. Forensic Sci. Int. 2008, 179, 219–240. [Google Scholar] [CrossRef]
- Yones, A.; Rasha, A.; Lamia, A.; Saly, Y. Identification of forensically important beetles on exposed human leftover parts in Assiut, Egypt during spring/summer season. Assiut Med. J. 2010, 34, 123–131. [Google Scholar]
- Matuszewski, S.; Szafałowicz, M.; Jarmusz, M. Insects colonising carcasses in open and forest habitats of Central Europe: Search for indicators of corpse relocation. Forensic Sci. Int. 2013, 231, 234–239. [Google Scholar] [CrossRef] [PubMed]
- Dupont, F.; Felix, B.; Daniel, C.; Champlain, D.-L. Biodiversity study of arthropods collected on rat carrion in Yaounde, Cameroon: First study on forensic entomology in Central Africa. Int. J. Biosci. 2012, 2, 1–8. [Google Scholar]
- Kelly, J.; Linde, T.; Van Der Anderson, G. The influence of clothing and wrapping on carcass decomposition and arthropod succession: A winter study in Central South Africa. Can. Soc. Forensic Sci. J. 2008, 41, 135–147. [Google Scholar] [CrossRef]
- Haberer, W.; Schmitt, T.; Peschke, K.; Schreier, P.; Müller, J.K. Ethyl 4-methyl heptanoate: A male-produced pheromone of Nicrophorus vespilloides. J. Chem. Ecol. 2008, 34, 94–98. [Google Scholar] [CrossRef] [PubMed]
- Francke, W.; Dettner, K. Chemical signalling in beetles. Top. Curr. Chem. 2005, 240, 85–166. [Google Scholar] [CrossRef]
- Dekeirsschieter, J.; Stefanuto, P.-H.; Brasseur, C.; Haubruge, E.; Focant, J.-F. Enhanced characterization of the smell of death by comprehensive two-dimensional gas chromatography-time-of-flight mass spectrometry (GCxGC-TOFMS). PLoS ONE 2012, 7. [Google Scholar] [CrossRef]
- Stefanuto, P.-H.; Perrault, K.; Stadler, S.; Pesesse, R.; Leblanc, H.; Forbes, S.; Focant, J.-F. GC × GC-TOFMS and supervised multivariate approaches to study human cadaveric decomposition olfactive signatures. Anal. Bioanal. Chem. 2015, 407, 4767–4778. [Google Scholar] [CrossRef]
- Armstrong, P.; Nizio, K.; Perrault, K.; Forbes, S. Establishing the volatile profile of pig carcasses as analogues for human decomposition during the early postmortem period. Heliyon 2016, 2, 1–24. [Google Scholar] [CrossRef]
- Von Hoermann, C.; Ruther, J.; Ayasse, M. The attraction of virgin female hide beetles (Dermestes maculatus) to cadavers by a combination of decomposition odour and male sex pheromones. Front. Zool. 2012, 9. [Google Scholar] [CrossRef] [PubMed]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martin, C.; Minchilli, D.; Francis, F.; Verheggen, F. Behavioral and Electrophysiological Responses of the Fringed Larder Beetle Dermestes frischii to the Smell of a Cadaver at Different Decomposition Stages. Insects 2020, 11, 238. https://doi.org/10.3390/insects11040238
Martin C, Minchilli D, Francis F, Verheggen F. Behavioral and Electrophysiological Responses of the Fringed Larder Beetle Dermestes frischii to the Smell of a Cadaver at Different Decomposition Stages. Insects. 2020; 11(4):238. https://doi.org/10.3390/insects11040238
Chicago/Turabian StyleMartin, Clément, Damien Minchilli, Frédéric Francis, and François Verheggen. 2020. "Behavioral and Electrophysiological Responses of the Fringed Larder Beetle Dermestes frischii to the Smell of a Cadaver at Different Decomposition Stages" Insects 11, no. 4: 238. https://doi.org/10.3390/insects11040238
APA StyleMartin, C., Minchilli, D., Francis, F., & Verheggen, F. (2020). Behavioral and Electrophysiological Responses of the Fringed Larder Beetle Dermestes frischii to the Smell of a Cadaver at Different Decomposition Stages. Insects, 11(4), 238. https://doi.org/10.3390/insects11040238