Using Nutritional Geometry to Explore How Social Insects Navigate Nutritional Landscapes


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
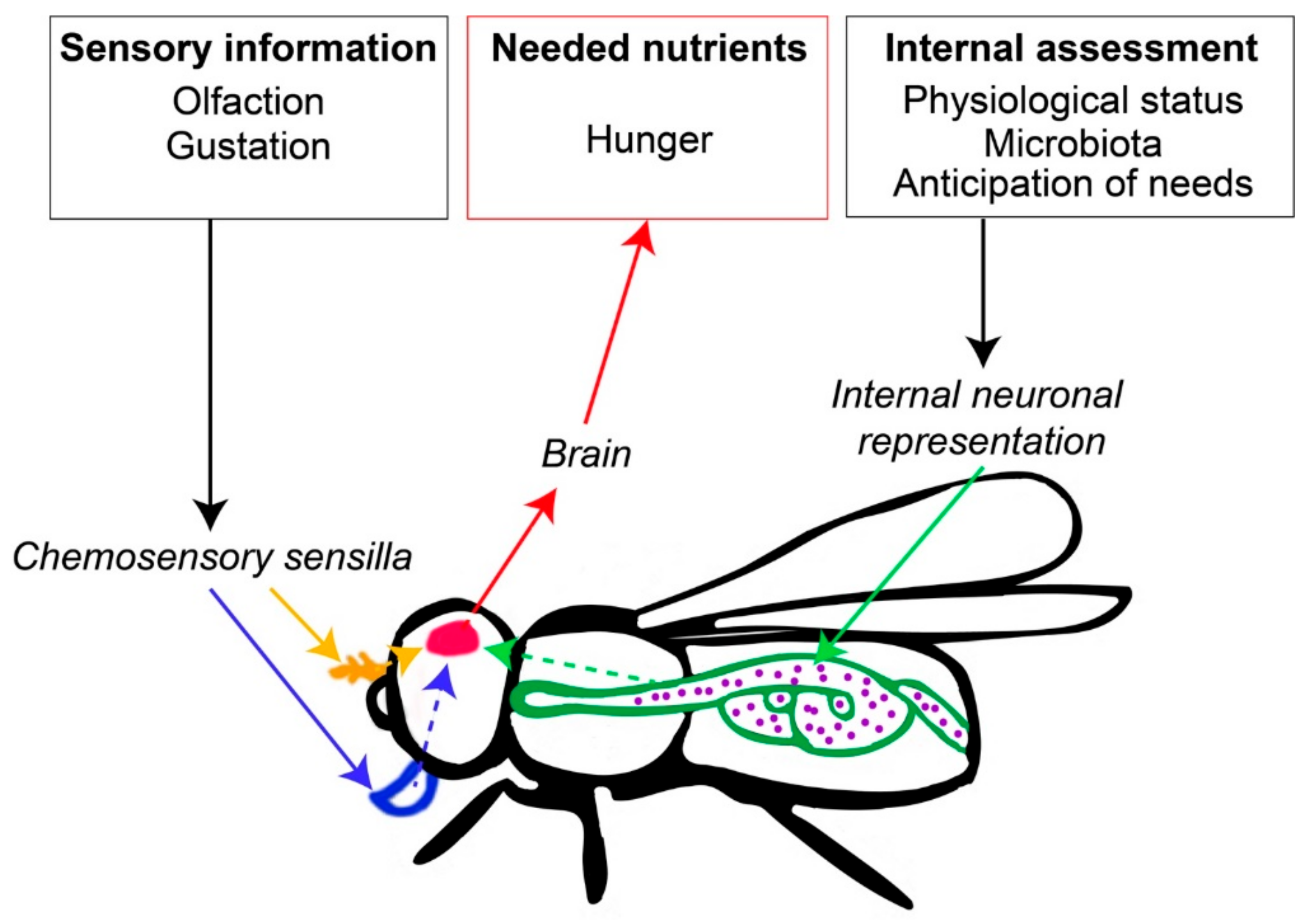
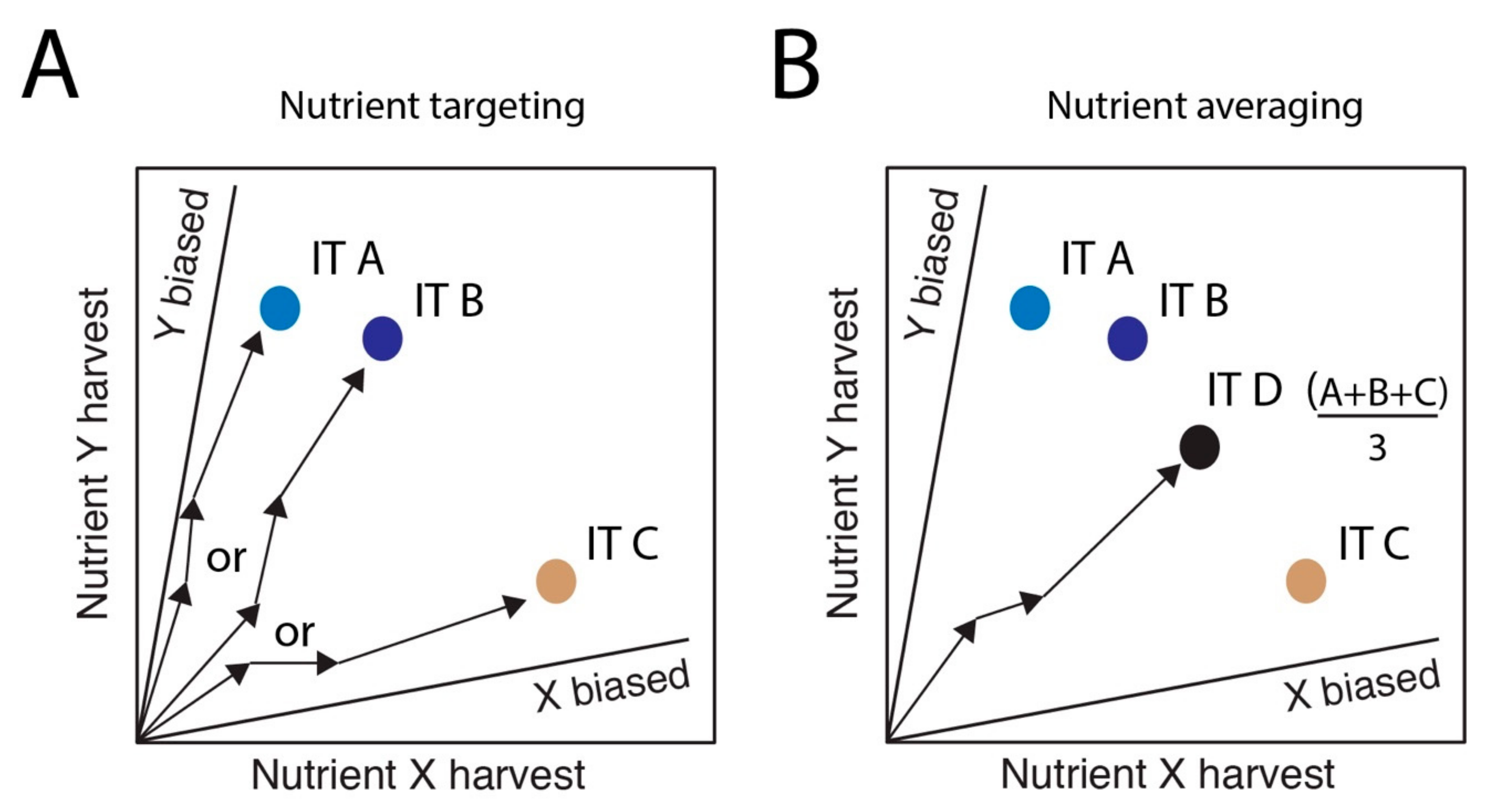
2. Cognitive Mechanisms Governing Nutritional Decisions
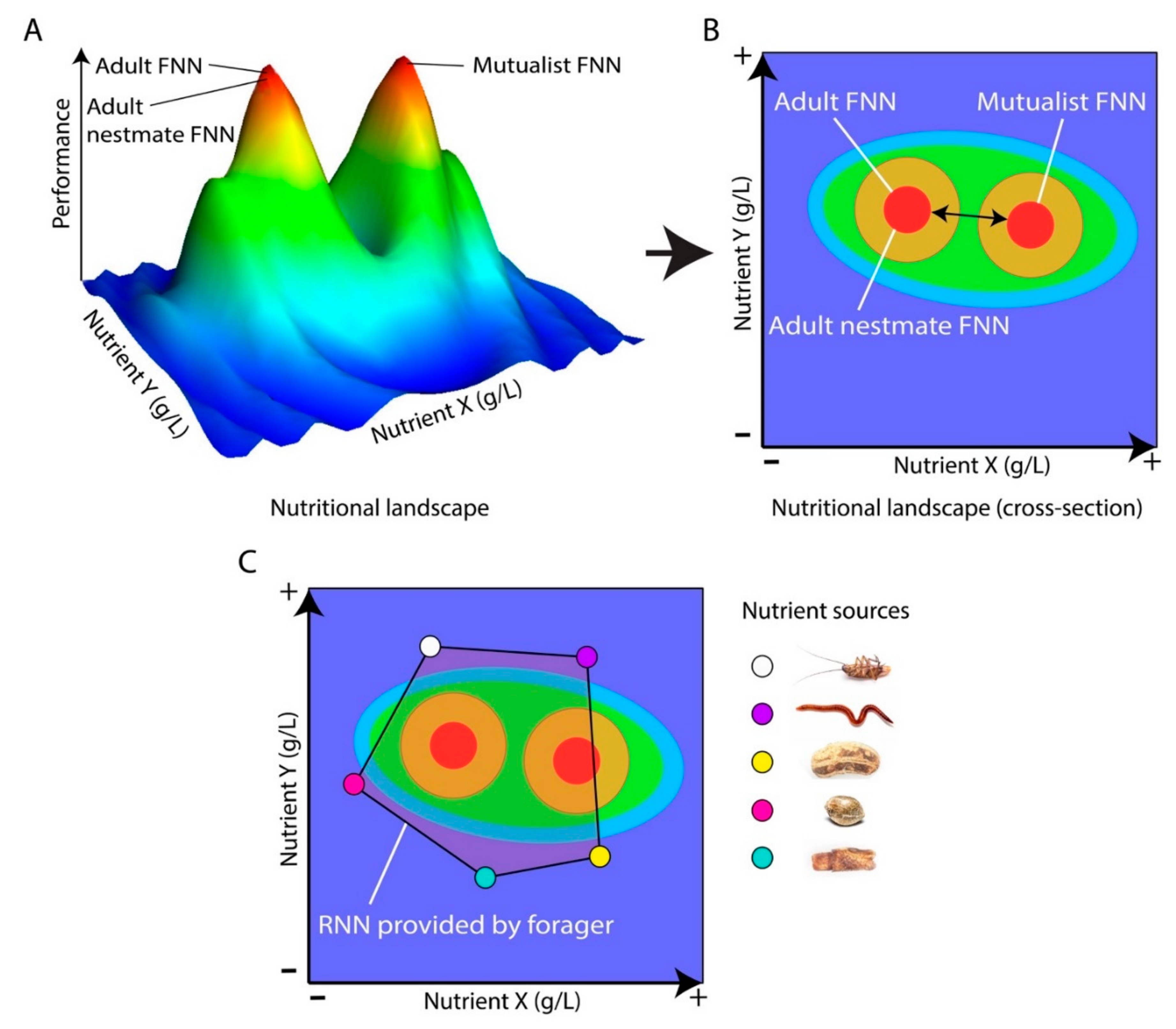
3. Using Ants to Apply NG to Cognition Research
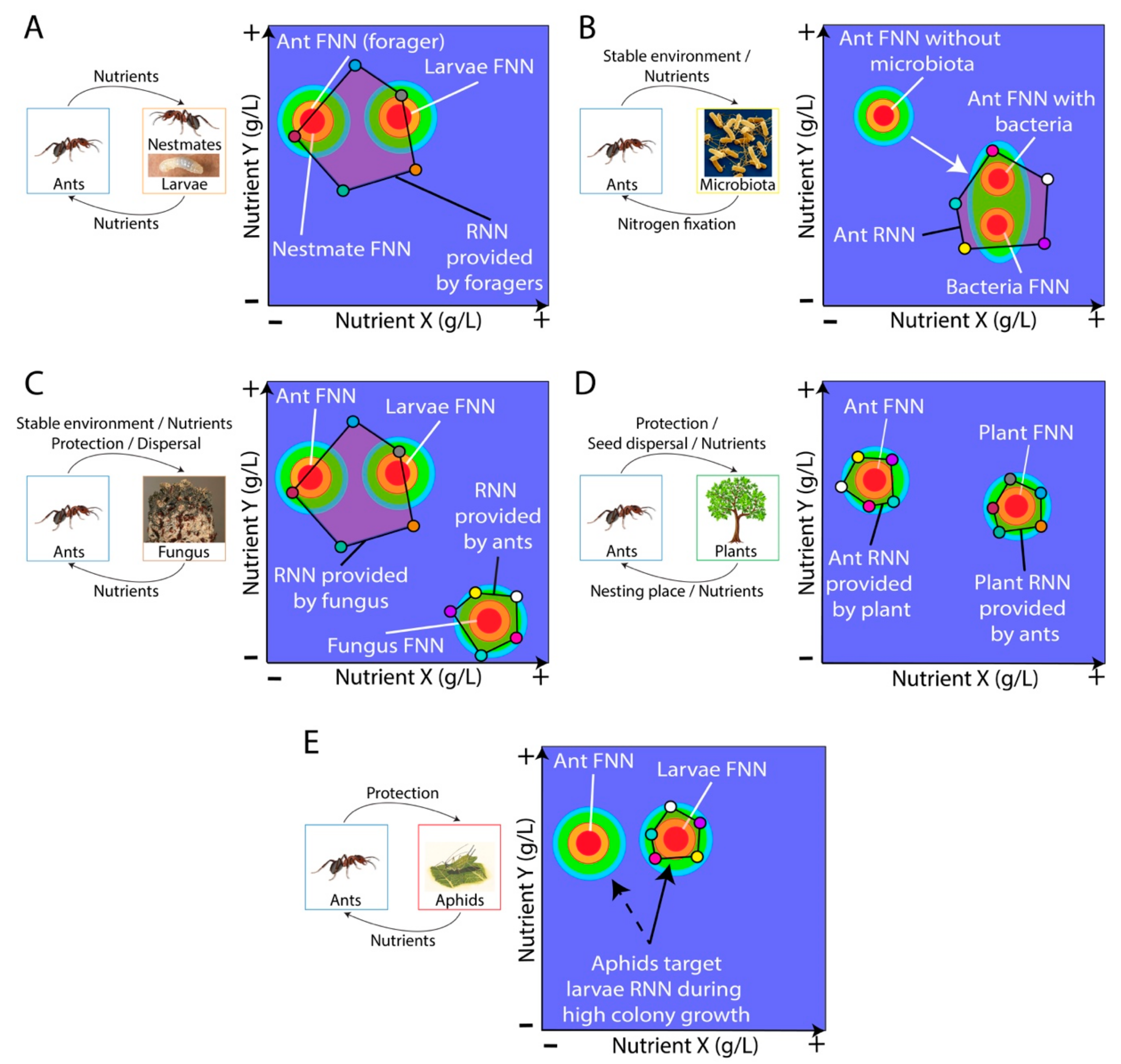
4. Mutualistic Relationships Involving Ants and an Unrelated Partner
4.1. Ant-Bacteria Mutualisms
4.2. Ant-Fungus Mutualisms
4.3. Ant-Plant Mutualisms
4.4. Ant-Insect Mutualisms
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Watanabe, K.; Kanaoka, Y.; Mizutani, S.; Uchiyama, H.; Yajima, S.; Watada, M.; Uemura, T.; Hattori, Y. Interspecies Comparative Analyses Reveal Distinct Carbohydrate-Responsive Systems among Drosophila Species. Cell Rep. 2019, 28, 2594. [Google Scholar] [CrossRef]
- Lee, K.P.; Simpson, S.J.; Clissold, F.J.; Brooks, R.; Ballard, J.W.O.; Taylor, P.W.; Soran, N.; Raubenheimer, D. Lifespan and reproduction in Drosophila: New insights from nutritional geometry. Proc. Natl. Acad. Sci. USA 2008, 105, 2498–2503. [Google Scholar] [CrossRef] [PubMed]
- Toshima, N.; Tanimura, T. Taste preference for amino acids is dependent on internal nutritional state in Drosophila melanogaster. J. Exp. Biol. 2012, 215, 2827–2832. [Google Scholar] [CrossRef] [PubMed]
- Hildebrand, J.G.; Shepherd, G.M. Mechanisms of olfactory discrimination: Converging evidence for common principles across phyla. Annu. Rev. Neurosci. 1997, 20, 595–631. [Google Scholar] [CrossRef] [PubMed]
- Amrein, H.; Thorne, N. Gustatory perception and behavior in Drosophila melanogaster. Curr. Biol. 2005, 15, R673–R684. [Google Scholar] [CrossRef]
- Yarmolinsky, D.A.; Zuker, C.S.; Ryba, N.J.P. Common sense about taste: From mammals to insects. Cell 2009, 139, 234–244. [Google Scholar] [CrossRef]
- Fujita, M.; Tanimura, T. Drosophila Evaluates and Learns the Nutritional Value of Sugars. Curr. Biol. 2011, 21, 751–755. [Google Scholar] [CrossRef]
- Itskov, P.M.; Ribeiro, C. The dilemmas of the gourmet fly: The molecular and neuronal mechanisms of feeding and nutrient decision making in Drosophila. Front. Neurosci. 2013, 7, 12. [Google Scholar] [CrossRef]
- Moreira, J.M.; Itskov, P.M.; Goldschmidt, D.; Baltazar, C.; Steck, K.; Tastekin, I.; Walker, S.J.; Ribeiro, C. optoPAD, a closed-loop optogenetics system to study the circuit basis of feeding behaviors. Elife 2019, 8, e43924. [Google Scholar] [CrossRef]
- Simpson, S.J.; Raubenheimer, D. The Nature of Nutrition: A Unifying Framework from Animal Adaptation to Human Obesity; Princeton University Press: Princeton, NJ, USA, 2012. [Google Scholar]
- Dussutour, A.; Nicolis, S.C.; Despland, E.; Simpson, S.J. Individual differences influence collective behaviour in social caterpillars. Anim. Behav. 2008, 76, 5–16. [Google Scholar] [CrossRef]
- Csata, E.; Dussutour, A. Nutrient regulation in ants (Hymenoptera: Formicidae): A review. Myrmecol. News 2019, 29, 111–124. [Google Scholar] [CrossRef]
- Lihoreau, M.; Buhl, J.; Charleston, M.A.; Sword, G.A.; Raubenheimer, D.; Simpson, S.J. Nutritional ecology beyond the individual: A conceptual framework for integrating nutrition and social interactions. Ecol. Lett. 2015, 18, 273–286. [Google Scholar] [CrossRef] [PubMed]
- Shik, J.Z.; Gomez, E.B.; Kooij, P.W.; Santos, J.C.; Wcislo, W.T.; Boomsma, J.J. Nutrition mediates the expression of cultivar-farmer conflict in a fungus-growing ant. Proc. Natl. Acad. Sci. USA 2016, 113, 10121–10126. [Google Scholar] [CrossRef] [PubMed]
- Behmer, S.T. Insect Herbivore Nutrient Regulation. Annu. Rev. Entomol. 2009, 54, 165–187. [Google Scholar] [CrossRef]
- Farina, W.M.; Grüter, C. Trophallaxis: A mechanism of information transfer. In Food Exploitation by Social Insects: Ecological, Behavioral, and Theoretical Approaches; CRC Press: Boca Raton, FL, USA, 2009. [Google Scholar]
- Nielsen, C.; Agrawal, A.A.; Hajek, A.E. Ants defend aphids against lethal disease. Biol. Lett. 2010, 6, 205–208. [Google Scholar] [CrossRef]
- Blatrix, R.; Mayer, V. Communication in Ant–Plant Symbioses. In Plant Communication from an Ecological Perspective; Baluška, F., Ninkovic, V., Eds.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 127–158. [Google Scholar] [CrossRef]
- Boucias, D.G.; Lietze, V.-U.; Teal, P. Chemical Signals That Mediate Insect-Fungal Interactions. In Biocommunication of Fungi; Witzany, G., Ed.; Springer: Dordrecht, The Netherlands, 2012; pp. 305–336. [Google Scholar] [CrossRef]
- Brandão, C.R.F.; Diniz, J.L.M.; Tomotake, E.M.J.I.S. Thaumatomyrmex strips millipedes for prey: A novel predatory behaviour in ants, and the first case of sympatry in the genus (Hymenoptera: Formicidae). Insectes Sociaux 1991, 38, 335–344. [Google Scholar] [CrossRef]
- Riveros, A.J.; Seid, M.A.; Wcislo, W.T. Evolution of brain size in class-based societies of fungus-growing ants (Attini). Anim. Behav. 2012, 83, 1043–1049. [Google Scholar] [CrossRef]
- Machovsky-Capuska, G.E.; Senior, A.M.; Simpson, S.J.; Raubenheimer, D. The Multidimensional Nutritional Niche. Trends Ecol. Evol. 2016, 31, 355–365. [Google Scholar] [CrossRef]
- Hawley, J.; Simpson, S.J.; Wilder, S.M. Flesh flies regulate the consumption of 3 macronutrients to maximize lifespan and egg production. Behav. Ecol. 2016, 27, 245–251. [Google Scholar] [CrossRef]
- Krabbe, B.A.; Arnan, X.; Lannes, P.; Bergstedt, C.E.; Larsen, R.S.; Pedersen, J.S.; Shik, J.Z. Using nutritional geometry to define the fundamental macronutrient niche of the widespread invasive ant Monomorium pharaonis. PLoS ONE 2019, 14, e0218764. [Google Scholar] [CrossRef]
- Raubenheimer, D. Toward a quantitative nutritional ecology: The right-angled mixture triangle. Ecol. Monogr. 2011, 81, 407–427. [Google Scholar] [CrossRef]
- Loewe, L. A framework for evolutionary systems biology. BMC Syst. Biol. 2009, 3, 27. [Google Scholar] [CrossRef] [PubMed]
- Dussutour, A.; Simpson, S.J. Communal Nutrition in Ants. Curr. Biol. 2009, 19, 740–744. [Google Scholar] [CrossRef] [PubMed]
- Hölldobler, B.; Wilson, E.O. The Superorganism: The beauty, Elegance, and Strangeness of Insect Societies; W.W. Norton: New York, NY, USA, 2009; p. 544. [Google Scholar]
- Martin, S.; Drijfhout, F. A Review of Ant Cuticular Hydrocarbons. J. Chem. Ecol. 2009, 35, 1151–1161. [Google Scholar] [CrossRef]
- Shik, J.Z.; Santos, J.C.; Seal, J.N.; Kay, A.; Mueller, U.G.; Kaspari, M. Metabolism and the Rise of Fungus Cultivation by Ants. Am. Nat. 2014, 184, 364–373. [Google Scholar] [CrossRef]
- Sorensen, A.A.; Vinson, S.B. Quantitative Food Distribution Studies within Laboratory Colonies of the Imported Fire Ant, Solenopsis-Invicta Buren. Insectes Sociaux 1981, 28, 129–160. [Google Scholar] [CrossRef]
- Weeks, R.D.; Wilson, L.T.; Vinson, S.B. Resource partitioning among colonies of polygyne red imported fire ants (Hymenoptera: Formicidae). Environ. Entomol. 2004, 33, 1602–1608. [Google Scholar] [CrossRef]
- Beattie, A.J. The Evolutionary ecology of Ant-Plant Mutualisms; Cambridge University Press: Cambridge, UK, 1985. [Google Scholar]
- Mueller, U.G.; Schultz, T.R.; Currie, C.R.; Adams, R.M.M.; Malloch, D. The origin of the attine ant-fungus mutualism. Q. Rev. Biol. 2001, 76, 169–197. [Google Scholar] [CrossRef]
- Russell, J.A.; Moreau, C.S.; Goldman-Huertas, B.; Fujiwara, M.; Lohman, D.J.; Pierce, N.E. Bacterial gut symbionts are tightly linked with the evolution of herbivory in ants. Proc. Natl. Acad. Sci. USA 2009, 106, 21236–21241. [Google Scholar] [CrossRef]
- Stadler, B.; Dixon, A.F.G. Mutualism: Ants and Their Insect Partners; Cambridge University Press: Cambridge, UK, 2008. [Google Scholar]
- Shanbhag, S.R.; Vazhappilly, A.T.; Sane, A.; D’Silva, N.M.; Tripathi, S. Electrolyte transport pathways induced in the midgut epithelium of Drosophila melanogaster larvae by commensal gut microbiota and pathogens. J. Physiol. Lond. 2017, 595, 523–539. [Google Scholar] [CrossRef]
- Feldhaar, H.; Gross, R. Insects as hosts for mutualistic bacteria. Int. J. Med. Microbiol. 2009, 299, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Russell, J.A.; Sanders, J.G.; Moreau, C.S. Hotspots for symbiosis: Function, evolution, and specificity of ant-microbe associations from trunk to tips of the ant phylogeny (Hymenoptera: Formicidae). Myrmecol. News 2017, 24, 43–69. [Google Scholar]
- Mehdiabadi, N.J.; Schultz, T.R. Natural history and phylogeny of the fungus-farming ants (Hymenoptera: Formicidae: Myrmicinae: Attini). Myrmecol. News 2010, 13, 37–55. [Google Scholar]
- Poulsen, M.; Currie, C.R. On ants, plants and fungi. N. Phytol. 2009, 182, 785–788. [Google Scholar] [CrossRef]
- Rico-Gray, V.; Oliveira, P.S. The Ecology and Evolution of Ant-Plant Interactions; University of Chicago Press: Chicago, IL, USA, 2007; p. 331. [Google Scholar]
- Kay, A.D.; Bruning, A.J.; van Alst, A.; Abrahamson, T.T.; Hughes, W.O.H.; Kaspari, M. A carbohydrate-rich diet increases social immunity in ants. Proc. Biol. Sci 2014, 281, 20132374. [Google Scholar] [CrossRef]
- Shik, J.Z.; Concilio, A.; Kaae, T.; Adams, R.M.M. The farming ant Sericomyrmex amabilis nutritionally manages its fungal symbiont and its social parasite. Ecol. Entomol. 2018, 43, 440–446. [Google Scholar] [CrossRef]
- Feldhaar, H.; Straka, J.; Krischke, M.; Berthold, K.; Stoll, S.; Mueller, M.J.; Gross, R. Nutritional upgrading for omnivorous carpenter ants by the endosymbiont Blochmannia. BMC Biol. 2007, 5, 48. [Google Scholar] [CrossRef]
- Feldhaar, H. Bacterial symbionts as mediators of ecologically important traits of insect hosts. Ecol. Entomol. 2011, 36, 533–543. [Google Scholar] [CrossRef]
- Sapountzis, P.; Zhukova, M.; Hansen, L.H.; Sorensen, S.J.; Schiott, M.; Boomsma, J.J. Acromyrmex Leaf-Cutting Ants Have Simple Gut Microbiota with Nitrogen-Fixing Potential. Appl. Environ. Microb. 2015, 81, 5527–5537. [Google Scholar] [CrossRef]
- Hu, Y.; Sanders, J.G.; Lukasik, P.; D’Amelio, C.L.; Millar, J.S.; Vann, D.R.; Lan, Y.M.; Newton, J.A.; Schotanus, M.; Kronauer, D.J.C.; et al. Herbivorous turtle ants obtain essential nutrients from a conserved nitrogen-recycling gut microbiome. Nat. Commun. 2018, 9, 964. [Google Scholar] [CrossRef]
- Sapountzis, P.; Zhukova, M.; Shik, J.Z.; Schiott, M.; Boomsma, J.J. Reconstructing the functions of endosymbiotic Mollicutes in fungus-growing ants. Elife 2018, 7, e39209. [Google Scholar] [CrossRef] [PubMed]
- Klein, A.; Schrader, L.; Gil, R.; Manzano-Marin, A.; Florez, L.; Wheeler, D.; Werren, J.H.; Latorre, A.; Heinze, J.; Kaltenpoth, M.; et al. A novel intracellular mutualistic bacterium in the invasive ant Cardiocondyla obscurior. ISME J. 2016, 10, 376–388. [Google Scholar] [CrossRef] [PubMed]
- Wong, A.C.N.; Wang, Q.P.; Morimoto, J.; Senior, A.M.; Lihoreau, M.; Neely, G.G.; Simpson, S.J.; Ponton, F. Gut Microbiota Modifies Olfactory-Guided Microbial Preferences and Foraging Decisions in Drosophila. Curr. Biol. 2017, 27, 2397. [Google Scholar] [CrossRef] [PubMed]
- Pasquaretta, C.; Gomez-Moracho, T.; Heeb, P.; Lihoreau, M. Exploring Interactions between the Gut Microbiota and Social Behavior through Nutrition. Genes 2018, 9, 534. [Google Scholar] [CrossRef]
- Holmes, A.J.; Chew, Y.V.; Colakoglu, F.; Cliff, J.B.; Klaassens, E.; Read, M.N.; Solon-Biet, S.M.; McMahon, A.C.; Cogger, V.C.; Ruohonen, K.; et al. Diet-Microbiome Interactions in Health Are Controlled by Intestinal Nitrogen Source Constraints. Cell Metab. 2017, 25, 140–151. [Google Scholar] [CrossRef]
- Defossez, E.; Selosse, M.A.; Dubois, M.P.; Mondolot, L.; Faccio, A.; Djieto-Lordon, C.; McKey, D.; Blatrix, R. Ant-plants and fungi: A new threeway symbiosis. N. Phytol. 2009, 182, 942–949. [Google Scholar] [CrossRef]
- Nepel, M.; Voglmayr, H.; Schonenberger, J.; Mayer, V.E. High Diversity and Low Specificity of Chaetothyrialean Fungi in Carton Galleries in a Neotropical Ant-Plant Association. PLoS ONE 2014, 9, e112756. [Google Scholar] [CrossRef]
- Maschwitz, U.; Hölldobler, B. Der Kartonnestbau bei Lasius fuliginosus Latr. (Hym. Formicidae). Z. Vgl. Physiol. 1970, 66, 176–189. [Google Scholar] [CrossRef]
- Blatrix, R.; Djieto-Lordon, C.; Mondolot, L.; La Fisca, P.; Voglmayr, H.; McKey, D. Plant-ants use symbiotic fungi as a food source: New insight into the nutritional ecology of ant-plant interactions. Proc. R. Soc. B Biol. Sci. 2012, 279, 3940–3947. [Google Scholar] [CrossRef]
- Shik, J.Z.; Rytter, W.; Arnan, X.; Michelsen, A. Disentangling nutritional pathways linking leafcutter ants and their co-evolved fungal symbionts using stable isotopes. Ecology 2018, 99, 1999–2009. [Google Scholar] [CrossRef]
- Nygaard, S.; Hu, H.F.; Li, C.; Schiott, M.; Chen, Z.S.; Yang, Z.K.; Xie, Q.L.; Ma, C.Y.; Deng, Y.; Dikow, R.B.; et al. Reciprocal genomic evolution in the ant-fungus agricultural symbiosis. Nat. Commun. 2016, 7, 12233. [Google Scholar] [CrossRef] [PubMed]
- Mueller, U.G.; Scott, J.J.; Ishak, H.D.; Cooper, M.; Rodrigues, A. Monoculture of Leafcutter Ant Gardens. PLoS ONE 2010, 5, e12668. [Google Scholar] [CrossRef] [PubMed]
- De Fine Licht, H.H.; Boomsma, J.J. Forage collection, substrate preparation, and diet composition in fungus-growing ants. Ecol. Entomol. 2010, 35, 259–269. [Google Scholar] [CrossRef]
- De Fine Licht, H.H.; Boomsma, J.J.; Tunlid, A. Symbiotic adaptations in the fungal cultivar of leaf-cutting ants. Nat. Commun. 2014, 5, 5675. [Google Scholar] [CrossRef]
- Quinlan, R.J.; Cherrett, J.M. Role of Fungus in the Diet of the Leaf-Cutting Ant Atta cephalotes (L.). Ecol. Entomol. 1979, 4, 151–160. [Google Scholar] [CrossRef]
- Martin, M.M.; Macconnell, J.G.; Gale, G.R. The Chemical Basis for the Attine Ant-Fungus Symbiosis. Absence of Antibiotics. Ann. Entomol. Soc. Am. 1969, 62, 386–388. [Google Scholar] [CrossRef]
- Green, P.W.C.; Kooij, P.W. The role of chemical signalling in maintenance of the fungus garden by leaf-cutting ants. Chemoecology 2018, 28, 101–107. [Google Scholar] [CrossRef]
- Moreau, C.S.; Bell, C.D. Testing the Museum Versus Cradle Tropical Biological Diversity Hypothesis: Phylogeny, Diversification, and Ancestral Biogeographic Range Evolution of the Ants. Evolution 2013, 67, 2240–2257. [Google Scholar] [CrossRef]
- Chomicki, G.; Staedler, Y.M.; Schonenberger, J.; Renner, S.S. Partner choice through concealed floral sugar rewards evolved with the specialization of ant-plant mutualisms. N. Phytol. 2016, 211, 1358–1370. [Google Scholar] [CrossRef]
- Koptur, S. Extrafloral nectary-mediated interactions between insects and plants. Insect Plant Interact. 1992, IV, 81–129. [Google Scholar]
- Fiala, B.; Maschwitz, U. Food Bodies and Their Significance for Obligate Ant-Association in the Tree Genus Macaranga (Euphorbiaceae). Bot. J. Linn. Soc. 1992, 110, 61–75. [Google Scholar] [CrossRef]
- Galetto, L.; Bernardello, L. Nectar Secretion Pattern and Removal Effects in Six Argentinean Pitcairnioideae (Bromeliaceae). Bot. Acta 1992, 105, 292–299. [Google Scholar] [CrossRef]
- Heil, M.; Baumann, B.; Kruger, R.; Linsenmair, K.E. Main nutrient compounds in food bodies of Mexican Acacia ant-plants. Chemoecology 2004, 14, 45–52. [Google Scholar] [CrossRef]
- Heil, M.; Hilpert, A.; Fiala, B.; Bin Hashim, R.; Strohm, E.; Zotz, G.; Linsenmair, K.E. Nutrient allocation of Macaranga triloba ant plants to growth, photosynthesis and indirect defence. Funct. Ecol. 2002, 16, 475–483. [Google Scholar] [CrossRef]
- O’Dowd, D.J. Pearl Bodies as Ant Food: An Ecological Role for Some Leaf Emergences of Tropical Plants. Biotropica 1982, 14, 40–49. [Google Scholar] [CrossRef]
- Hatada, A.; Itioka, T.; Yamaoka, R.; Itino, T. Carbon and nitrogen contents of food bodies in three myrmecophytic species of Macaranga: Implications for antiherbivore defense mechanisms. J. Plant Res. 2002, 115, 179–184. [Google Scholar] [CrossRef]
- Sagers, C.L.; Ginger, S.M.; Evans, R.D. Carbon and nitrogen isotopes trace nutrient exchange in an ant-plant mutualism. Oecologia 2000, 123, 582–586. [Google Scholar] [CrossRef]
- Culver, D.C.; Beattie, A.J. Myrmecochory in Viola—Dynamics of Seed-Ant Interactions in Some West-Virginia Species. J. Ecol. 1978, 66, 53–72. [Google Scholar] [CrossRef]
- Youngsteadt, E.; Nojima, S.; Haberlein, C.; Schulz, S.; Schal, C. Seed odor mediates an obligate ant-plant mutualism in Amazonian rainforests. Proc. Natl. Acad. Sci. USA 2008, 105, 4571–4575. [Google Scholar] [CrossRef]
- Bond, W.; Slingsby, P. Collapse of an Ant-Plant Mutalism: The Argentine Ant (Iridomyrmex Humilis) and Myrmecochorous Proteaceae. Ecology 1984, 65, 1031–1037. [Google Scholar] [CrossRef]
- Janzen, D.H. Interaction of the bull’s-horn acacia (Acacia cornigera L.) with an ant inhabitant (Pseudomyrmex ferrugineus F. Smith) in eastern Mexico. Kans. Univ. Sci. Bull. 1967, 47, 315–558. [Google Scholar]
- Kusmenoglu, S.; Rockwood, L.L.; Gretz, M.R. Fatty-Acids and Diacylglycerols from Elaiosomes of Some Ant-Dispersed Seeds. Phytochemistry 1989, 28, 2601–2602. [Google Scholar] [CrossRef]
- Lanza, J.; Schmitt, M.A.; Awad, A.B. Comparative chemistry of elaiosomes of three species of Trillium. J. Chem. Ecol. 1992, 18, 209–221. [Google Scholar] [CrossRef] [PubMed]
- Hughes, L.; Westoby, M.; Jurado, E. Convergence of Elaiosomes and Insect Prey—Evidence from Ant Foraging Behavior and Fatty-Acid Composition. Funct. Ecol. 1994, 8, 358–365. [Google Scholar] [CrossRef]
- Fischer, R.C.; Olzant, S.M.; Wanek, W.; Mayer, V. The fate of Corydalis cava elaiosomes within an ant colony of Myrmica rubra: Elaiosomes are preferentially fed to larvae. Insectes Sociaux 2005, 52, 55–62. [Google Scholar] [CrossRef]
- Chomicki, G.; Renner, S.S. Partner abundance controls mutualism stability and the pace of morphological change over geologic time. Proc. Natl. Acad. Sci. USA 2017, 114, 3951–3956. [Google Scholar] [CrossRef]
- Youngsteadt, E.; Baca, J.A.; Osborne, J.; Schal, C. Species-Specific Seed Dispersal in an Obligate Ant-Plant Mutualism. PLoS ONE 2009, 4, e4335. [Google Scholar] [CrossRef]
- Blüthgen, N.; Schmit-Neuerburg, V.; Engwald, S.; Barthlott, W. Ants as epiphyte gardeners: Comparing the nutrient quality of ant and termite canopy substrates in a Venezuelan lowland rain forest. J. Trop. Ecol. 2001, 17, 887–894. [Google Scholar] [CrossRef]
- Hughes, L.; Westoby, M. Fate of Seeds Adapted for Dispersal by Ants in Australian Sclerophyll Vegetation. Ecology 1992, 73, 1285–1299. [Google Scholar] [CrossRef]
- Orona-Tamayo, D.; Wielsch, N.; Blanco-Labra, A.; Svatos, A.; Farias-Rodriguez, R.; Heil, M. Exclusive rewards in mutualisms: Ant proteases and plant protease inhibitors create a lock-key system to protect Acacia food bodies from exploitation. Mol. Ecol. 2013, 22, 4087–4100. [Google Scholar] [CrossRef]
- Stadler, B.; Dixon, A.F.G. Ecology and evolution of aphid-ant interactions. Annu. Rev. Ecol. Evol. Syst. 2005, 36, 345–372. [Google Scholar] [CrossRef]
- Nash, D.R.; Als, T.D.; Boomsma, J.J. Survival and growth of parasitic Maculinea alcon caterpillars (Lepidoptera, Lycaenidae) in laboratory nests of three Myrmica ant species. Insectes Sociaux 2011, 58, 391–401. [Google Scholar] [CrossRef]
- Nixon, G.E.J. The Association of Ants with Aphids and Coccids; Commonwealth Institute of Entomology: London, UK, 1951; p. 36. [Google Scholar]
- Volkl, W.; Woodring, J.; Fischer, M.; Lorenz, M.W.; Hoffmann, K.H. Ant-aphid mutualisms: The impact of honeydew production and honeydew sugar composition on ant preferences. Oecologia 1999, 118, 483–491. [Google Scholar] [CrossRef] [PubMed]
- Fischer, M.K.; Shingleton, A.W. Host plant and ants influence the honeydew sugar composition of aphids. Funct. Ecol. 2001, 15, 544–550. [Google Scholar] [CrossRef]
- Shik, J.Z.; Silverman, J. Towards a nutritional ecology of invasive establishment: Aphid mutualists provide better fuel for incipient Argentine ant colonies than insect prey. Biol. Invasions 2013, 15, 829–836. [Google Scholar] [CrossRef]
- Helms, K.R.; Vinson, S.B. Plant resources and colony growth in an invasive ant: The importance of honeydew-producing Hemiptera in carbohydrate transfer across trophic levels. Environ. Entomol. 2008, 37, 487–493. [Google Scholar] [CrossRef]
- Kiss, A. Melezitose, Aphids and Ants. Oikos 1981, 37, 382. [Google Scholar] [CrossRef]
- Fischer, M.K.; Volkl, W.; Schopf, R.; Hoffmann, K.H. Age-specific patterns in honeydew production and honeydew composition in the aphid Metopeurum fuscoviride: Implications for ant-attendance. J. Insect Physiol. 2002, 48, 319–326. [Google Scholar] [CrossRef]
- Cloutier, C.; Mackauer, M. The effect of parasitism by Aphidius smithi (Hymenoptera: Aphidiidae) on the food budget of the pea aphid, Acyrthosiphon pisum (Homoptera: Aphididae). Can. J. Zool. 1979, 57, 1605–1611. [Google Scholar] [CrossRef]
- Volkl, W. Aphids or Their Parasitoids—Who Actually Benefits from Ant-Attendance. J. Anim. Ecol. 1992, 61, 273–281. [Google Scholar] [CrossRef]
- Sakata, H. How an Ant Decides to Prey on or to Attend Aphids. Popul. Ecol. 1994, 36, 45–51. [Google Scholar] [CrossRef]
- Becerra, J.X.I.; Venable, D.L. Extrafloral Nectaries—A Defense against Ant-Homoptera Mutualisms. Oikos 1989, 55, 276–280. [Google Scholar] [CrossRef]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Crumière, A.J.J.; Stephenson, C.J.; Nagel, M.; Shik, J.Z. Using Nutritional Geometry to Explore How Social Insects Navigate Nutritional Landscapes. Insects 2020, 11, 53. https://doi.org/10.3390/insects11010053
Crumière AJJ, Stephenson CJ, Nagel M, Shik JZ. Using Nutritional Geometry to Explore How Social Insects Navigate Nutritional Landscapes. Insects. 2020; 11(1):53. https://doi.org/10.3390/insects11010053
Chicago/Turabian StyleCrumière, Antonin J. J., Calum J. Stephenson, Manuel Nagel, and Jonathan Z. Shik. 2020. "Using Nutritional Geometry to Explore How Social Insects Navigate Nutritional Landscapes" Insects 11, no. 1: 53. https://doi.org/10.3390/insects11010053
APA StyleCrumière, A. J. J., Stephenson, C. J., Nagel, M., & Shik, J. Z. (2020). Using Nutritional Geometry to Explore How Social Insects Navigate Nutritional Landscapes. Insects, 11(1), 53. https://doi.org/10.3390/insects11010053