Bionomics and Ecological Services of Megaloptera Larvae (Dobsonflies, Fishflies, Alderflies)




Abstract
1. An Ancient Holometabolous Order
2. Distribution and Diversity
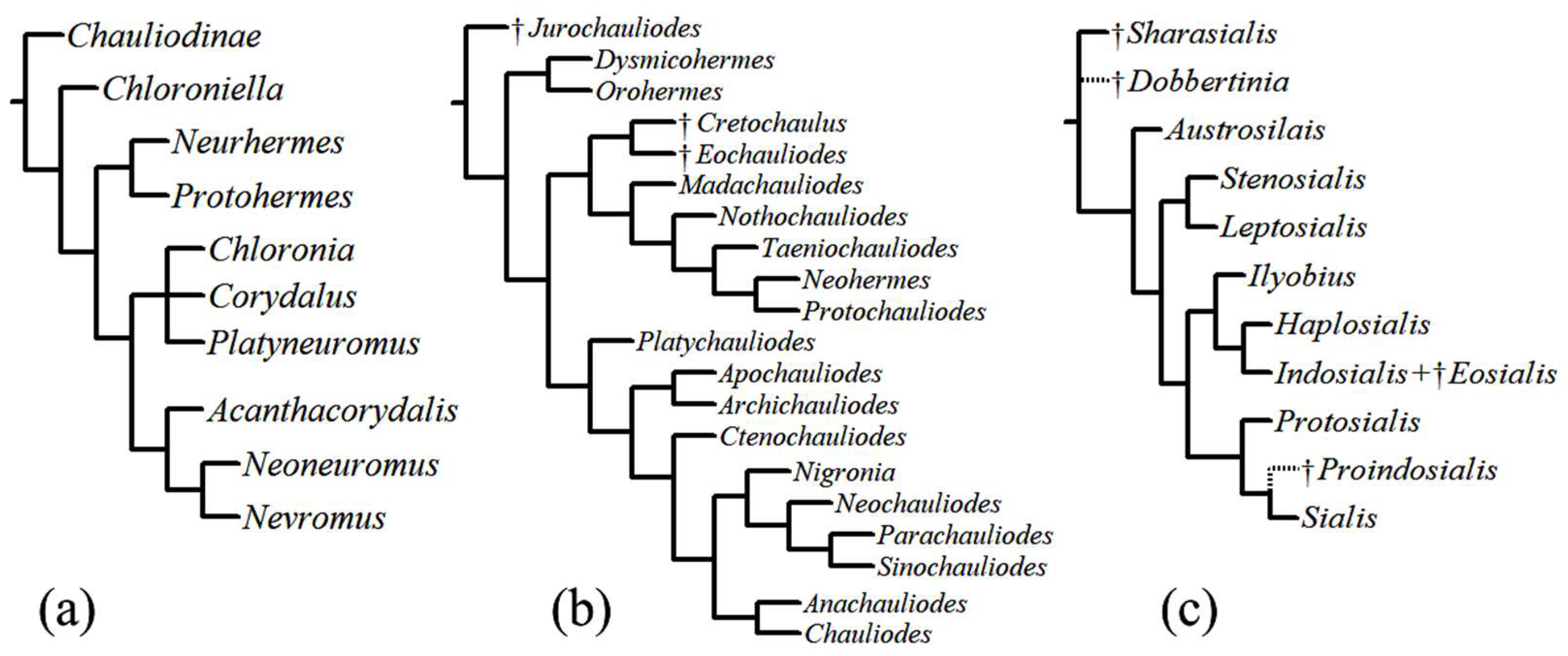
3. Origin and Fossil Record
4. Phylogenetic Relationships
5. Evolution of Ecological Roles
6. Essential Services for Ecosystem Function
Food for Other Organisms and Uses for Humans
7. Anatomical, Physiological, and Behavioral Adaptations for Life in Water
8. Responses to Threats of Water Pollution and Climate Change
9. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ponomarenko, A.G. Superorder Myrmeleontidea Latreille, 1802 (=Neuropteroidea Handlirsch, 1903). In History of Insects, 1st ed.; Rasnitsyn, A.P., Quicke, D.L., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2002; pp. 176–192. ISBN 14-020-0026-X. [Google Scholar]
- Grimaldi, D.; Engel, M.S. Evolution of the Insects; Cambridge University Press: Cambridge, NY, USA, 2005; p. 755. ISBN 0-521-82149-5. [Google Scholar]
- Contreras-Ramos, A. Systematics of the Dobsonfly Genus Corydalus (Megaloptera: Corydalidae). Thomas Say Publications in Entomology: Monographs; Entomological Society of America: Annapolis, MD, USA, 1998; p. 360. ISBN 0-938522-70-1. [Google Scholar]
- Cao, C.; Liu, X. Description of the final-instar larva and pupa of Acanthacorydalis orientalis (McLachlan, 1899) (Megaloptera: Corydalidae) with some life history notes. Zootaxa 2013, 3691, 145–152. [Google Scholar] [CrossRef]
- New, T.R.; Theischinger, G. Megaloptera (Alderflies, Dobsonflies) Handbuch der Zoologie; Walter de Gruyter: Berlin, Germany, 1993; Volume 4, p. 102. [Google Scholar]
- Glorioso, M.J.; Flint, O.S., Jr. A review of the genus Platyneuromus (Insecta: Neuroptera: Corydalidae). Proc. Biol. Soc. Wash. 1984, 97, 601–614. [Google Scholar]
- Ramírez-Ponce, A.; Garfias-Lozano, G.; Contreras-Ramos, A. The nature of allometry in an exaggerated trait: The postocular flange in Platyneuromus Weele (Insecta: Megaloptera). PLoS ONE 2017, 12, e0172388. [Google Scholar] [CrossRef]
- Zhao, C.; Liu, X.; Yang, D. Wing base structural data support the sister relationship of Megaloptera and Neuroptera (Insecta: Neuropterida). PLoS ONE 2014, 9, e114695. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.Y.; Wang, Y.J.; Shih, C.K.; Ren, D.; Yang, D. Early evolution and historical biogeography of fishflies (Megaloptera: Chauliodinae): Implications from a phylogeny combining fossil and extant taxa. PLoS ONE 2012, 7, e40345. [Google Scholar] [CrossRef] [PubMed]
- Winterton, S.L.; Hardy, N.B.; Wiegmann, B.M. On wings of lace: Phylogeny and Bayesian divergence time estimates of Neuropterida (Insecta) based on morphological and molecular data. Syst. Entomol. 2010, 35, 349–378. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, X.Y.; Winterton, S.L.; Yang, D. The first mitochondrial genome for the fishfly subfamily Chauliodinae and implications for the higher phylogeny of Megaloptera. PLoS ONE 2012, 7, e47302. [Google Scholar] [CrossRef]
- Liu, X.Y.; Lü, Y.; Aspök, H.; Yang, D.; Aspök, U. Homology of the genital sclerites of Megaloptera (Insecta: Neuropterida) and their phylogenetic relevance. Syst. Entomol. 2015, 41, 256–286. [Google Scholar] [CrossRef]
- Engel, M.S.; Winterton, S.L.; Breitkreuz, L.C.V. Phylogeny and evolution of Neuropterida: Where have wings of lace taken us? Annu. Rev. Entomol. 2018, 63, 531–551. [Google Scholar] [CrossRef]
- Cover, M.R.; Resh, V.H. Global diversity of dobsonflies, fishflies, and alderflies (Megaloptera; Insecta) and spongillaflies, nevrorthids, and osmylids (Neuroptera; Insecta) in freshwater. Hydrobiologia 2008, 595, 409–417. [Google Scholar] [CrossRef]
- Monserrat, V.J. Los megalópteros de la Península Ibérica (Insecta, Neuropterida, Megaloptera, Sialidae). Graellsia 2014, 70, 9. [Google Scholar] [CrossRef]
- Bowles, D.; Contreras-Ramos, A. First record of the family Sialidae (Megaloptera) from Thailand and description of the female and putative larva of Indosialis bannaensis. Zootaxa 2016, 4114, 485–491. [Google Scholar] [CrossRef]
- Jung, S.W.; Vshivkova, T.S.; Bae, Y.J. DNA-based identification of South Korean Megaloptera larvae with taxonomic notes. Can. Entomol. 2016, 148, 123–139. [Google Scholar] [CrossRef]
- Archangelsky, M.; Pessacq, P.; Berrondo, M. Description of the larva of Ilyobius chilensis (McLachlan) (Megaloptera: Sialidae) and notes on the adult morphology. Zootaxa 2017, 4318, 177–186. [Google Scholar] [CrossRef]
- Misof, B.; Liu, S.; Meusemann, K.; Peters, R.S.; Donath, A.; Mayer, C.; Frandsen, P.B.; Ware, J.; Flouri, T.; Beutel, R.G.; et al. Phylogenomics resolves the timing and pattern of insect evolution. Science 2014, 346, 763–767. [Google Scholar] [CrossRef]
- Winterton, S.L.; Lemmon, A.R.; Gillung, J.P.; Garzon, I.J.; Badano, D.; Bakkes, D.K.; Beitkreuz, L.C.V.; Engel, M.S.; Lemmon, E.M.; Liu, X.Y.; et al. Evolution of lacewings and allied orders using anchored phylogenomics (Neuroptera, Megaloptera, Raphidioptera). Syst. Entomol. 2017, 43, 330–354. [Google Scholar] [CrossRef]
- Shcherbakov, D.E. Permian ancestors of Hymenoptera and Raphidioptera. ZooKeys 2013, 358, 45–67. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liu, X.; Garzón-Orduña, I.J.; Winterton, S.L.; Yan, Y.; Aspöck, U.; Aspöck, H.; Yang, D. Mitochondrial phylogenomics illuminates the evolutionary history of Neuropterida. Cladistics 2016, 33, 617–636. [Google Scholar] [CrossRef]
- Jepson, J.E.; Heads, S.W. Fossil Megaloptera (Insecta: Neuropterida) from the Lower Cretaceous Crato Formation of Brazil. Zootaxa 2016, 4098, 134–144. [Google Scholar] [CrossRef]
- Wichard, W.; Chatterton, C.; Ross, A. Corydasialidae fam. n. (Megaloptera) from Baltic Amber. Insect Syst. Evol. 2005, 36, 279–284. [Google Scholar] [CrossRef]
- Archibald, S.B.; Makarkin, V.N. The second genus and species of the extinct neuropteroid family Corydasialidae, from early Eocene McAbee, British Columbia, Canada: Do they belong to Megaloptera? Zootaxa 2015, 4040, 569–575. [Google Scholar] [CrossRef]
- Beutel, R.G.; Friedrich, F.; Yang, X.; Ge, S. Insect Morphology and Phylogeny, 1st ed.; De Gruyter: Berlin, Germany, 2014; p. 516. ISBN 978-3110262636. [Google Scholar]
- Kristensen, N.P. Phylogeny of insect orders. Annu. Rev. Entomol. 1981, 26, 135–157. [Google Scholar] [CrossRef]
- Beutel, R.G.; Friedrich, F.; Aspöck, U. The larval head of Nevrorthidae and the phylogeny of Neuroptera (Insecta). Zool. J. Linn. Soc. Lond. 2010, 158, 533–562. [Google Scholar] [CrossRef]
- Beutel, R.G.; Friedrich, F.; Hörnschemeyer, T.; Pohl, H.; Hünefeld, F.; Beckmann, F.; Meier, R.; Misof, B.; Whiting, M.F.; Vilhelmsen, L. Morphological and molecular evidence converge upon a robust phylogeny of the megadiverse Holometabola. Cladistics 2011, 27, 341–355. [Google Scholar] [CrossRef]
- Van der Weele, H.W. Megaloptera (Latreille), monographic revision. In Collections Zoologiques du Baron Edm. de Selys Longchamps Fasc. V (Premiere partie), 1st ed.; HathiTrust’s Digital Library: Bruxelles, Belgium, 1910; p. 93. [Google Scholar]
- Contreras-Ramos, A. Is the family Corydalidae (Neuropterida, Megaloptera) a monophylum? Denisia 2004, 13, 135–140. [Google Scholar]
- Contreras-Ramos, A. Phylogenetic review of dobsonflies of the subfamily Corydalinae and the genus Corydalus Latreille (Megaloptera: Corydalidae). Zootaxa 2011, 2862, 1–38. [Google Scholar] [CrossRef]
- Glorioso, M.J. Systematics of the dobsonfly subfamily Corydalinae (Megaloptera: Corydalidae). Syst. Entomol. 1981, 6, 253–290. [Google Scholar] [CrossRef]
- Cardoso-Costa, G.; Azevêdo, C.A.S.; Ferreira, N., Jr. New genus and new species of Chauliodinae (Insecta: Megaloptera: Corydalidae) from Brazil. Zootaxa 2013, 3613, 391–399. [Google Scholar] [CrossRef]
- Cover, M.R.; Bogan, M.T. Minor Insect Orders. In Thorp and Covich’s Freshwater Invertebrates: Ecology and General Biology, 4th ed.; Thorp, J.H., Rogers, D.C., Eds.; Elsevier: London, UK, 2015; Volume 1, pp. 1059–1072. [Google Scholar]
- Liu, X.Y.; Hayashi, F.; Yang, D. Phylogeny of the family Sialidae (Insecta: Megaloptera) inferred from morphological data, with implications for generic classification and historical biogeography. Cladistics 2015, 31, 18–49. [Google Scholar] [CrossRef]
- Flint, O.S., Jr.; Evans, E.D.; Neunzig, H.H. Megaloptera and aquatic Neuroptera. In An Introduction to the Aquatic Insects of North America, 4th ed.; Merritt, R.W., Cummins, K.W., Berg, M.E., Eds.; Kendall Hunt: Dubuque, IA, USA, 2008; pp. 425–437. ISBN 978-0757563218. [Google Scholar]
- Azam, K.M.; Anderson, N.H. Life history and habits of Sialis rotunda and S. californica in Western Oregon. Ann. Entomol. Soc. Am. 1969, 62, 549–558. [Google Scholar] [CrossRef]
- Ardila-Camacho, A.; Contreras-Ramos, A. The Megaloptera (Insecta: Neuropterida) of Colombia. Aquat. Insects 2018, 3, 297–353. [Google Scholar] [CrossRef]
- Hayashi, F.; Nakane, M. Radio tracking and activity monitoring of the dobsonfly larva, Protohermes grandis (Megaloptera: Corydalidae). Oecologia 1989, 78, 468–472. [Google Scholar] [CrossRef]
- Fuller, R.L.; Rand, P.S. Influence of substrate type on vulnerability of prey to predacious aquatic insects. J. N. Am. Benthol. Soc. 1990, 9, 1–8. [Google Scholar] [CrossRef]
- Hayashi, F. Nanocladius (Plecopteracoluthus) asiaticus sp. n. (Diptera: Chironomidae) phoretic on dobsonfly and fishfly larvae (Megaloptera: Corydalidae). Aquat. Insects 1998, 20, 215–229. [Google Scholar] [CrossRef]
- Evans, E.D. A Study of the Megaloptera of the Pacific Coastal Region of the United States. Ph.D. Thesis, Oregon State University, Corvalis, OR, USA, 1972; p. 210. [Google Scholar]
- Fashing, N.J. A novel habitat for larvae of the fishfly Chauliodes pectinicornis (Megaloptera: Corydalidae). Banisteria 1994, 3, 25–26. [Google Scholar]
- Smith, E.L. Biology and structure of the dobsonfly, Neohermes californicus (Walker) (Megaloptera: Corydalidae). Pan Pac. Entomol. 1970, 46, 142–150. [Google Scholar]
- Contreras-Ramos, A. Mating behavior of Platyneuromus (Megaloptera: Corydalidae), with life history notes on dobsonflies from Mexico and Costa Rica. Entomol. News 1999, 110, 125–135. [Google Scholar]
- Geijskes, D.C. Notes on Megaloptera from the Guyanas, S. Am. In Progress in World’s Neuropterology; Gepp, J., Aspöck, H., Hölzel, H., Eds.; Food and Agriculture Organization: Graz, Austria, 1984; pp. 79–84. [Google Scholar]
- Contreras-Ramos, A. Richard W. Merritt, Kenneth W. Cummins y Martin B. Berg (eds.). 2008. An introduction to the aquatic insects of North America, 4a edición. Kendall/Hunt Publishing Company, Dubuque, 1158 p. + CD con clave interactive [book review]. Rev. Mex. Biodivers. 2010, 81, 593–595. [Google Scholar]
- Cover, M.R.; Seo, J.H.; Resh, V.H. Life history, burrowing behavior, and distribution of Neohermes filicornis (Megaloptera: Corydalidae), a long-lived aquatic insect in intermittent streams. West. N. Am. Nat. 2015, 75, 474–490. [Google Scholar] [CrossRef]
- Contreras-Ramos, A. Megaloptera. In Macroinvertebrados Bentónicos Sudamericanos-Sistemática y Biología; Domínguez, E., Fernández, H.R., Eds.; Fundación Miguel Lillo: Tucumán, Argentina, 2009; pp. 233–245. ISBN 978-950-668-015-2. [Google Scholar]
- Contreras-Ramos, A. Notes on some Neotropical Alderflies (Sialidae: Megaloptera). Ann. Entomol. Soc. Am. 2008, 101, 808–814. [Google Scholar] [CrossRef]
- Roldán-Pérez, G. Guía Para el Estudio de los Macroinvertebrados Acuáticos del Departamento de Antioquia, 1st ed.; FEN Colombia, Colciencias: Bogotá, Colombia, 1998; p. 217. ISBN 958-9129-04-8. [Google Scholar]
- Buzzi, Z.J.; Miyazaki, R.D. Entomologia Didática, 3th ed.; Editora da UFPR: Curitiba, Brazil, 1999; p. 306. ISBN 8573350318. [Google Scholar]
- Shepard, W.D. Pigs as corydalid predators. Pan Pac. Entomol. 2004, 80, 66. [Google Scholar]
- Epperson, C.R.; Short, R.A. Annual Production of Corydalus cornutus (Megaloptera) in the Guadalupe River, Texas. Am. Midl. Nat. 1987, 118, 433–438. [Google Scholar] [CrossRef]
- Bowles, D.E. Life history and variability of secondary production estimates for Corydalus cornutus (Megaloptera: Corydalidae) in an Ozark Stream. J. Agric. Entomol. 1990, 7, 61–70. [Google Scholar]
- Kelsey, L.P. The skeleto-muscular mechanism of the dobsonfly. Corydalus cornutus Part I. Head and prothorax. CUAES Monogr. 1954, 34, 1–42. [Google Scholar]
- Beutel, R.G.; Friedrich, F. Comparative study of larval head structures of Megaloptera (Hexapoda). Eur. J. Entomol. 2008, 105, 917–938. [Google Scholar] [CrossRef]
- Yamamoto, K.; Toh, Y. The fine structure of the lateral ocellus of the dobsonfly larva. J. Morph. 1975, 146, 415–430. [Google Scholar] [CrossRef]
- Paulus, H.F. Comparative morphology of the larval eyes of Neuropteroidea. In Recent Research in Neuropterology (Proceedings of the 2nd International Symposium on Neuropterology, 21–23 August); Gepp, J., Aspöck, H., Hölzel, H., Eds.; Food and Agriculture Organization: Graz, Germany, 1986; pp. 157–164. [Google Scholar]
- Cao, C.; Liu, Z.; Chen, S.; Tong, C. The swimming behavior of the aquatic larva of Neoneuromus ignobilis (Megaloptera: Corydalidae: Corydalinae). Acta Ent. Sin. 2012, 55, 133–138. [Google Scholar]
- Takeuchi, Y.; Hoshiba, H. The life histories of three species of Corydalidae (Megaloptera) from Japan. Aquat. Insects 2012, 34, 55–63. [Google Scholar] [CrossRef]
- Contreras-Ramos, A.; Harris, S.C. The immature stages of Platyneuromus (Corydalidae), with a key to the genera of larval Megaloptera of Mexico. J. N. Am. Benthol. Soc. 1998, 17, 489–517. [Google Scholar] [CrossRef]
- Sánchez-Bayo, F.; Wyckhuys, K.A.G. Worldwide decline of the entomofauna: A review of its drivers. Biol. Conserv. 2019, 232, 8–27. [Google Scholar] [CrossRef]
- Barbour, M.T.; Gerritsen, J.; Snyder, B.D.; Stribling, J.B. Rapid Bioassessment Protocols for Use in Streams and Wadeable Rivers: Periphyton, Benthic Macroinvertebrates and Fish, 2nd ed.; EPA 841-B-99-002; U.S. Environmental Protection Agency, Office of Water: Washington, DC, USA, 1999; p. 339.
- Martín-Torrijos, L.; Sandoval-Sierra, J.V.; Muñoz, J.; Diéguez-Uribeondo, J.; Bosch, J.; Guayasamin, J.M. Rainbow trout (Oncorhynchus mykiss) threaten Andean amphibians. Neotrop. Biodiver. 2016, 2, 26–36. [Google Scholar] [CrossRef]
- Arias-Pineda, J.; Pedroza-Martínez, D.R. Presencia del cangrejo rojo Procambarus clarkii (Girard, 1852) en la sabana de Bogotá, Colombia. Bol. Soc. Entomol. Aragon. 2018, 62, 283–286. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genus | Number of Species | Biogeographic Region |
|---|---|---|
| CORYDALIDAE: Corydalinae | ||
| Acanthacorydalis van der Weele, 1907 | 8 | Oriental |
| Chloronia Banks, 1908 | 18 | Nearctic, Neotropical |
| Chloroniella Esben-Petersen, 1924 | 1 | Afrotropical |
| Corydalus Latreille, 1802 | 39 | Nearctic, Neotropical, Oriental ** |
| Neoneuromus van der Weele, 1909 | 14 | Oriental, Palaearctic |
| Neurhermes Navás, 1915 | 7 | Oriental |
| Nevromus Rambur, 1842 | 6 | Oriental |
| Platyneuromus van der Weele, 1909 | 3 | Nearctic, Neotropical |
| Protohermes van der Weele, 1907 | 78 | Oriental, Palaearctic |
| CORYDALIDAE: Chauliodinae | ||
| Anachauliodes Kimmins, 1954 | 1 | Oriental |
| Apochauliodes Theischinger, 1983 | 1 | Australian |
| Archichauliodes van der Weele, 1909 | 21 | Australian, Neotropical |
| Chauliodes Latreille, 1802 | 2 | Nearctic |
| Ctenochauliodes van der Weele, 1909 | 13 | Oriental |
| Dysmicohermes Munroe, 1953 | 2 | Nearctic |
| Madachauliodes Paulian, 1951 | 3 | Afrotropical |
| Neochauliodes van der Weele, 1909 | 47 | Oriental, Palaearctic |
| Neohermes Banks, 1908 | 6 | Nearctic |
| Nigronia Banks, 1908 | 2 | Nearctic |
| Nothochauliodes Flint, 1983 | 1 | Neotropical |
| Orohermes Evans, 1984 | 1 | Nearctic |
| Parachauliodes van der Weele, 1909 | 7 | Oriental, Palaearctic |
| Platychauliodes Esben-Petersen, 1924 | 3 | Afrotropical |
| Protochauliodes van der Weele, 1909 | 17 * | Australian, Nearctic, Neotropical |
| Puri Cardoso-Costa et al., 2013 | 1 | Neotropical |
| Sinochauliodes Liu & Yang, 2006 | 4 | Oriental |
| Taeniochauliodes Esben-Petersen, 1924 | 8 | Afrotropical |
| SIALIDAE: Sialidinae | ||
| Austrosialis Tillyard, 1919 | 2 | Australian |
| Haplosialis Navás, 1927 | 2 | Afrotropical |
| Ilyobius Enderlein, 1910 | 9 | Neotropical, Palaearctic |
| Indosialis Lestage, 1927 | 3 | Oriental, Palaearctic |
| Leptosialis Esben-Petersen, 1920 | 2 | Afrotropical |
| Protosialis van der Weele, 1909 | 3 | Nearctic, Neotropical |
| Sialis Latreille, 1802 | 60 | Nearctic, Oriental, Palaearctic |
| Stenosialis Tillyard, 1919 | 2 | Australian |
| Total: 35 genera | 397 spp. | |
| Genus | Species | Locality-Biogeographic Region | Period (Ma) |
|---|---|---|---|
| CORYDALIDAE: Corydalinae | |||
| † Corydalites Scudder, 1878 | 1 | USA-Nearctic | Cenozoic (66 Ma) |
| Corydalus Latreille, 1802 | 1 | DEU-Palaearctic | Late Jurassic (152 Ma) |
| † Cratocorydalopsis Jepson & Heads, 2016 | 1 | BRA-Neotropical | Early Cretaceous (115 Ma) |
| † Lithocorydalus Jepson & Heads, 2016 | 1 | BRA-Neotropical | Early Cretaceous (115 Ma) |
| CORYDALIDAE: Chauliodinae | |||
| Chauliodes Latreille, 1802 | 2 * | RUS-Palaearctic | Middle Eocene (47.8 Ma) |
| † Chauliosialis Ponomarenko, 1976 | 1 | RUS-Palaearctic | Late Cretaceous (89.8 Ma) |
| † Cretochaulus Ponomarenko, 1976 | 1 | RUS-Palaearctic | Early Cretaceous (139.4 Ma) |
| † Eochauliodes Liu et al., 2012 | 1 | CHN-Oriental | Middle Jurassic (168.3 Ma) |
| † Jurochauliodes Wang & Zhang, 2010 | 1 | CHN-Oriental | Middle Jurassic (168.3 Ma) |
| † CORYDASIALIDAE | |||
| † Corydasialis Wichard et al., 2005 | 1 * | RUS-Palearctic | Late Eocene (33.9–37.8 Ma) |
| † Ypresioneura Archibald & Makark., 2015 | 1 | CAN-Nearctic | Early Eocene (47.8–56 Ma) |
| † NANOSIALIDAE: † Nanosialinae | |||
| † Hymega Shcherbakov, 2013 | 1 | RUS-Palaearctic | Late Permian (259.8 Ma) |
| † Lydasialis Shcherbakov, 2013 | 1 | RUS-Palaearctic | Late Permian (259.8 Ma) |
| † Nanosialis Shcherbakov, 2013 | 2 | RUS-Palaearctic | Late Permian (259.8 Ma) |
| † Raphisialis Shcherbakov, 2013 | 1 | RUS-Palaearctic | Late Permian (259.8 Ma |
| † PARASIALIDAE | |||
| † Parasialis Ponomarenko, 1977 | 4 | RUS, MNG-Palaearctic | Permian (279.3 Ma) |
| † Sojanasialis Ponomarenko, 1977 | 1 | RUS-Palaearctic | Middle Permian (272.3 Ma) |
| SIALIDAE: † Sharasialinae | |||
| † Sharasialis Ponomarenko, 2012 | 1 | MNG-Palaearctic | Late Jurassic (163.5 Ma) |
| SIALIDAE: Sialidinae | |||
| † Dobbertinia Handlirsch, 1920 | 1 | DEU-Palaearctic | Early Jurassic (182.7 Ma) |
| † Eosialis Nel et al., 2002 | 1 | FRA-Palaearctic | Early Eocene (56 Ma) |
| Ilyobius Enderlein, 1910 | 3 * | DOM-Neotropical, RUS-Palearctic | Eocene, Miocene (47.8–5.3 Ma) |
| Indosialis Lestage, 1927 | 1 | TUR-Palaearctic | Oligocene (33.9 Ma) |
| † Proindosialis Nel, 1988 | 1 | FRA-Palaearctic | Late Miocene (7.2 Ma) |
| Sialis Latreille, 1802 | 4 * | FRA, DEU-Palaearctic | Eocene, Miocene, Pliocene (47.8–2.5 Ma) |
| Total: 24 genera | 34 spp. | ||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rivera-Gasperín, S.L.; Ardila-Camacho, A.; Contreras-Ramos, A. Bionomics and Ecological Services of Megaloptera Larvae (Dobsonflies, Fishflies, Alderflies). Insects 2019, 10, 86. https://doi.org/10.3390/insects10040086
Rivera-Gasperín SL, Ardila-Camacho A, Contreras-Ramos A. Bionomics and Ecological Services of Megaloptera Larvae (Dobsonflies, Fishflies, Alderflies). Insects. 2019; 10(4):86. https://doi.org/10.3390/insects10040086
Chicago/Turabian StyleRivera-Gasperín, Sara Lariza, Adrian Ardila-Camacho, and Atilano Contreras-Ramos. 2019. "Bionomics and Ecological Services of Megaloptera Larvae (Dobsonflies, Fishflies, Alderflies)" Insects 10, no. 4: 86. https://doi.org/10.3390/insects10040086
APA StyleRivera-Gasperín, S. L., Ardila-Camacho, A., & Contreras-Ramos, A. (2019). Bionomics and Ecological Services of Megaloptera Larvae (Dobsonflies, Fishflies, Alderflies). Insects, 10(4), 86. https://doi.org/10.3390/insects10040086