Evidence of Leaf Consumption Rate Decrease in Fall Armyworm, Spodoptera frugiperda, Larvae Parasitized by Coccygidium luteum


 , and
, and
Abstract
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Insect Collection and Rearing
2.2. Feeding Tests on S. frugiperda Larvae
2.3. Generation Time of C. luteum
2.4. Statistical Analysis
3. Results
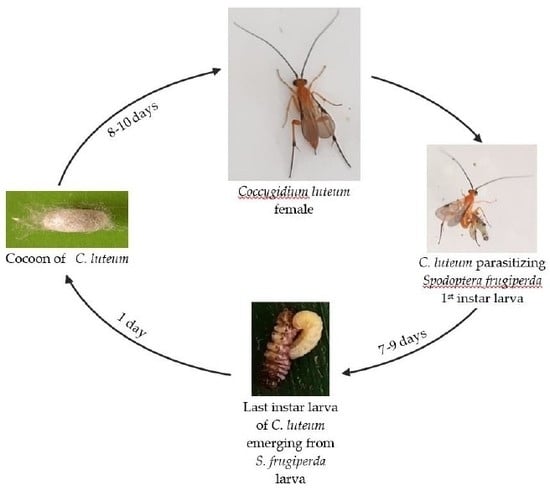
3.1. Generation Time of C. luteum
3.2. Leaf Consumption Rate of Parasitized and Unparasitized S. frugiperda Larvae
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Statistics Research and Information Directorate (SRID). Agriculture in Ghana: Facts and Figures 2015, 25th ed.; Report of MOFA (Ministry of Food and Agriculture); MOFA: Accra, Ghana, 2016; 113p.
- Devi, S. Fall armyworm threatens food security in southern Africa. Lancet 2018, 391, 727. [Google Scholar] [CrossRef]
- Sparks, A.N. A review of the biology of the fall armyworm. Fla. Entomol. 1979, 62, 82–87. [Google Scholar] [CrossRef]
- FAO. Briefing Note on FAO Actions on Fall Armyworm in Africa; FAO: Rome, Italy. Available online: www.fao.org/3/BS183E/bs183e.pdf (accessed on 14 May 2019).
- Wolfe, L.M. Why alien invaders succeed: Support for the escape-from-enemy hypothesis. Am. Nat. 2002, 160, 705–711. [Google Scholar] [CrossRef] [PubMed]
- Colautti, R.; Ricciardi, A.; Grigorovich, I.; MacIsaac, H. Is invasion success explained by the enemy release hypothesis? Ecol. Lett. 2004, 7, 721–733. [Google Scholar] [CrossRef]
- Murùa, G.; Virla, E. Population parameters of Spodoptera frugiperda (Smith) (Lep.: Noctuidae) fed on corn and two predominant grasses in Tucuman (Argentina). Acta Zool. Mex. 2004, 20, 199–210. [Google Scholar]
- Goergen, G.; Kumar, P.L.; Sankung, S.B.; Togola, A.; Tamò, M. First report of outbreaks of the fall armyworm Spodoptera frugiperda (JE Smith) (Lepidoptera, Noctuidae), a new alien invasive pest in West and Central Africa. PLoS ONE 2016, 11, e0165632. [Google Scholar] [CrossRef]
- Day, R.; Abrahams, P.; Bateman, M.; Beale, T.; Clottey, V.; Cock, M.; Colmenarez, Y.; Corniani, N.; Early, R.; Godwin, J.; et al. Fall armyworm: Impacts and implications for Africa. Outlooks Pest Manag. 2017, 28, 196–201. [Google Scholar] [CrossRef]
- Cock, M.J.W.; Beseh, P.K.; Buddie, A.G.; Cafá, G.; Crozier, J. Molecular methods to detect Spodoptera frugiperda in Ghana, and implications for monitoring the spread of invasive species in developing countries. Sci. Rep. 2017, 7, 4103. [Google Scholar] [CrossRef]
- Nagoshi, R.N.; Goergen, G.; Tounou, K.A.; Agboka, K.; Koffi, D.; Meagher, R.L. Analysis of strain distribution, migratory potential, and invasion history of fall armyworm populations in northern Sub-Saharan Africa. Sci. Rep. 2018, 8, 3710. [Google Scholar] [CrossRef]
- Montezano, D.G.; Specht, A.; Sosa-Gómez, D.R.; Roque-Specht, V.F.; Sousa-Silva, J.C.; Paula-Moraes, S.V.; Peterson, J.A.; Hunt, T.E. Host Plants of Spodoptera frugiperda (Lepidoptera: Noctuidae) in the Americas. Afr. Entomol. 2018, 26, 286–300. [Google Scholar] [CrossRef]
- Baudron, F.; Zaman-Allah, M.A.; Chaipa, I.; Chari, N.; Chinwada, P. Understanding the factors influencing fall armyworm (Spodoptera frugiperda J.E. Smith) damage in African smallholder maize fields and quantifying its impact on yield. A case study in Eastern Zimbabwe. Crop Prot. 2019, 120, 141–150. [Google Scholar] [CrossRef]
- Early, R.; González-Moreno, P.; Murphy, S.T.; Day, R. Forecasting the global extent of invasion of the cereal pest Spodoptera frugiperda, the fall armyworm. NeoBiota 2018, 40, 25–50. [Google Scholar] [CrossRef]
- Feldmann, F.; Rieckmann, U.; Winter, S. The spread of the fall armyworm Spodoptera frugiperda in Africa—What should be done next? J. Plant Dis. Prot. 2019, 126, 97–101. [Google Scholar] [CrossRef]
- Bateman, M.L.; Day, R.K.; Luke, B.; Edgington, S.; Kuhlmann, U.; Cock, M.J.W. Assessment of potential biopesticide options for managing fall armyworm (Spodoptera frugiperda) in Africa. J. Appl. Entomol. 2018, 142, 805–819. [Google Scholar] [CrossRef]
- Kumela, T.; Simiyu, J.; Sisay, B.; Likhayo, P.; Mendesil, E.; Gohole, L.; Tefera, T. Farmers’ knowledge, perceptions, and management practices of the new invasive pest, fall armyworm (Spodoptera frugiperda) in Ethiopia and Kenya. Int. J. Pest Manag. 2018. [Google Scholar] [CrossRef]
- Sisay, B.; Simiyu, J.; Malusi, P.; Likhayo, P.; Mendesil, E.; Elibariki, N.; Wakgari, M.; Ayalew, G.; Tefera, T. First report of the fall armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae), natural enemies from Africa. J. Appl. Entomol. 2018, 142, 800–804. [Google Scholar] [CrossRef]
- Yu, S.J. Insecticide Resistance in the Fall Armyworm, Spodoptera frugiperda (J. E. Smith). Pestic. Biochem. Phys. 1991, 39, 84–91. [Google Scholar] [CrossRef]
- Farias, J.R.; Andow, D.A.; Horikoshi, R.J.; Sorgatto, R.J.; Fresia, P.; dos Santos, A.C.; Omoto, C. Field-evolved resistance to Cry1F maize by Spodoptera frugiperda (Lepidoptera: Noctuidae) in Brazil. Crop Prot. 2014, 64, 150–158. [Google Scholar] [CrossRef]
- Togola, A.; Meseka, S.; Menkir, A.; Badu-Apraku, B.; Bouka, O.; Tamò, M.; Djouaka, R. Measurement of Pesticide Residues from Chemical Control of the Invasive Spodoptera frugiperda (Lepidoptera: Noctuidae) in a Maize Experimental Field in Mokwa, Nigeria. Int. J. Environ. Res. Public Health 2018, 15, 849. [Google Scholar] [CrossRef]
- Prasanna, B.M.; Huesing, J.E.; Eddy, R.; Peschke, V.M. Fall Armyworm in Africa: A Guide for Integrated Pest Management, 1st ed.; CIMMYT: Mexico City, Mexico, 2018; 109p. [Google Scholar]
- Sanap, Y.; Dawkar, V.V.; Giri, A.P.; Sen, A.; Pandit, R.S. Parasitism by Chelonus blackburni (Hymenoptera) affects food consumption and development of Helicoverpa armigera (Lepidoptera) and cellular architecture of the midgut. J. Asia Pac. Entomol. 2016, 19, 65–70. [Google Scholar] [CrossRef]
- Agboyi, L.K.; Goergen, G.; Beseh, P.; Mensah, S.A.; Clottey, V.A.; Glikpo, R.; Buddie, A.; Cafà, G.; Offord, L.; Rwomushana, I.; et al. Parasitoid complex of fall armyworm, Spodoptera frugiperda in Ghana and Benin. Insects. Draft manuscript.
- Van Noort, S. WaspWeb: Hymenoptera of the Afrotropical Region. 2019. Available online: www.waspweb.org (accessed on 16 June 2019).
- Sharkey, M.J. Cladistics and tribal classification of the Agathidinae (Hymenoptera: Braconidae). J. Nat. Hist. 1992, 26, 425–447. [Google Scholar] [CrossRef]
- Sharkey, M.J. Two new genera of Agathidinae (Hymenoptera: Braconidae) with a key to the genera of the New World. Zootaxa 2006, 1185, 37–51. [Google Scholar] [CrossRef]
- Farahani, S.; Talebi, A.; Rakhshani, E.; Achterberg, C.; Sharkey, M. A contribution to the knowledge of Agathidinae (Hymenoptera: Braconidae) from Iran with description of a new species. Biologia 2014, 69, 228–235. [Google Scholar] [CrossRef]
- Oluwaranti, A.; Fakorede, M.A.B.; Badu-Apraku, B. Grain yield of maize varieties of different maturity groups under marginal rainfall conditions. J. Agric. Sci. 2008, 53, 183–191. [Google Scholar] [CrossRef]
- Hoballah, M.E.; Degen, T.; Bergvinson, D.; Savidan, A.; Tamò, C.; Turlings, T.C. Occurrence and direct control potential of parasitoids and predators of the fall armyworm (Lepidoptera: Noctuidae) on maize in the subtropical lowlands of Mexico. Agric. For. Entomol. 2004, 6, 83–88. [Google Scholar] [CrossRef]
- Morales, J.; Medina, P.; Vinuela, E. The influence of two endoparasitic wasps, Hyposoter didymator and Chelonus inanitus, on the growth and food consumption of their host larva Spodoptera littoralis. BioControl 2007, 52, 145–160. [Google Scholar] [CrossRef]
- Schmidt, O.; Theopold, U.; Strand, M. Innate immunity and its evasion and suppression by hymenopteran endoparasitoids. BioEssays 2001, 23, 344–351. [Google Scholar] [CrossRef]
- Godfray, H.C.J. Parasitoids. Encycl. Biodivers. 2007, 1–13. [Google Scholar] [CrossRef]
- Yamanaka, A.; Hayakawa, Y.; Noda, H.; Nakashima, N.; Watanabe, H. Characterization of polydnavirus-encoded mRNA in parasitized armyworm larvae. Insect Biochem. Mol. Biol. 1996, 26, 529–536. [Google Scholar] [CrossRef]
- Asgari, S.; Zareie, R.; Zhang, G.; Schmidt, O. Isolation and characterization of a novel venom protein from an endoparasitoid, Cotesia rubecula (Hym: Braconidae). Arch. Insect Biochem. 2003, 53, 92–100. [Google Scholar] [CrossRef]
- Rivers, D. Venoms of Endoparasitic Wasps. In Encyclopedia of Entomology; Capinera, J.L., Ed.; Springer: Dordrecht, The Netherlands, 2008; pp. 4092–4093. [Google Scholar]
- Fang, Q.; Wang, F.; Gatehouse, J.A.; Gatehouse, A.M.R.; Chen, X.; Hu, C.; Ye, G. Venom of parasitoid, Pteromalus puparum, suppresses host, Pieris rapae, immune promotion by decreasing Host C-Type Lectin Gene Expression. PLoS ONE 2011, 6, e26888. [Google Scholar] [CrossRef] [PubMed]
- Noguchi, H.; Hayakawa, Y. Mechanism of parasitism-induced elevation of dopamine levels in host insect larvae. Insect Biochem. Mol. Biol. 1996, 269, 659–665. [Google Scholar] [CrossRef]
- Beckage, N.E.; Riddiford, L.M. Effects of parasitism by Apanteles congregatus on the endocrine physiology of the tobacco hornworm Manduca sexta. Gen. Comp. Endocrinol. 1982, 47, 308–322. [Google Scholar] [CrossRef]
- Beckage, N.E.; Riddiford, L.M. Developmental interactions between the tobacco hornworm Manduca sexta and its braconid parasite Apanteles congregates L. Entomol. Exp. Appl. 1978, 23, 139–151. [Google Scholar] [CrossRef]
- Grieshop, M.J.; Flinn, P.W.; Nechols, J.R. Biological control of Indianmeal moth (Lepidoptera: Pyralidae) on finished stored products using egg and larval parasitoids. J. Econ. Entomol. 2006, 99, 1080–1084. [Google Scholar] [CrossRef]
- Ba, M.N.; Baoua, I.B.; Kaboré, A.; Amadou, L.; Oumarou, N.; Dabire-Binso, C.; Sanon, A. Augmentative on-farm delivery methods for the parasitoid Habrobracon hebetor Say (Hymenoptera: Braconidae) to control the millet head miner Heliocheilus albipunctella (de Joannis) (Lepidoptera: Noctuidae) in Burkina Faso and Niger. BioControl 2014, 59, 689–696. [Google Scholar] [CrossRef]
- Kenis, M.; du Plessis, H.; Van den Berg, J.; Ba, M.N.; Goergen, G.; Kwadjo, K.E.; Baoua, I.; Tefera, T.; Buddie, A.; Cafà, G.; et al. Telenomus Remus, a candidate parasitoid for the biological control of Spodoptera Frugiperda in Africa, is already present on the continent. Insects 2019, 10, 92. [Google Scholar] [CrossRef]
- Sisay, B.; Simiyu, J.; Mendesil, E.; Likhayo, P.; Ayalew, G.; Mohamed, S.; Subramanian, S.; Tefera, T. Fall Armyworm, Spodoptera frugiperda Infestations in East Africa: Assessment of Damage and Parasitism. Insects 2019, 10, 195. [Google Scholar] [CrossRef]
- Agboyi, L.K.; Djade, K.M.; Ahadji-Dabla, K.M.; Ketoh, G.K.; Nuto, Y.; Glitho, I.A. Vegetable production in Togo and potential impact of pesticides use practices on the environment. Int. J. Biol. Chem. Sci. 2015, 9, 723–736. [Google Scholar] [CrossRef]


© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Agboyi, L.K.; Mensah, S.A.; Clottey, V.A.; Beseh, P.; Glikpo, R.; Rwomushana, I.; Day, R.; Kenis, M. Evidence of Leaf Consumption Rate Decrease in Fall Armyworm, Spodoptera frugiperda, Larvae Parasitized by Coccygidium luteum. Insects 2019, 10, 410. https://doi.org/10.3390/insects10110410
Agboyi LK, Mensah SA, Clottey VA, Beseh P, Glikpo R, Rwomushana I, Day R, Kenis M. Evidence of Leaf Consumption Rate Decrease in Fall Armyworm, Spodoptera frugiperda, Larvae Parasitized by Coccygidium luteum. Insects. 2019; 10(11):410. https://doi.org/10.3390/insects10110410
Chicago/Turabian StyleAgboyi, Lakpo Koku, Samuel Adjei Mensah, Victor Attuquaye Clottey, Patrick Beseh, Raymond Glikpo, Ivan Rwomushana, Roger Day, and Marc Kenis. 2019. "Evidence of Leaf Consumption Rate Decrease in Fall Armyworm, Spodoptera frugiperda, Larvae Parasitized by Coccygidium luteum" Insects 10, no. 11: 410. https://doi.org/10.3390/insects10110410
APA StyleAgboyi, L. K., Mensah, S. A., Clottey, V. A., Beseh, P., Glikpo, R., Rwomushana, I., Day, R., & Kenis, M. (2019). Evidence of Leaf Consumption Rate Decrease in Fall Armyworm, Spodoptera frugiperda, Larvae Parasitized by Coccygidium luteum. Insects, 10(11), 410. https://doi.org/10.3390/insects10110410