The Competitive Mating of Irradiated Brown Marmorated Stink Bugs, Halyomorpha halys, for the Sterile Insect Technique

 ,
,  ,
, 
 ,
,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Experiment 1. Male Dose Response
2.2. Experiment 2. Overflooding
2.3. Experiment 3. Overflooding and Competition
2.4. Statistical Methods
3. Results
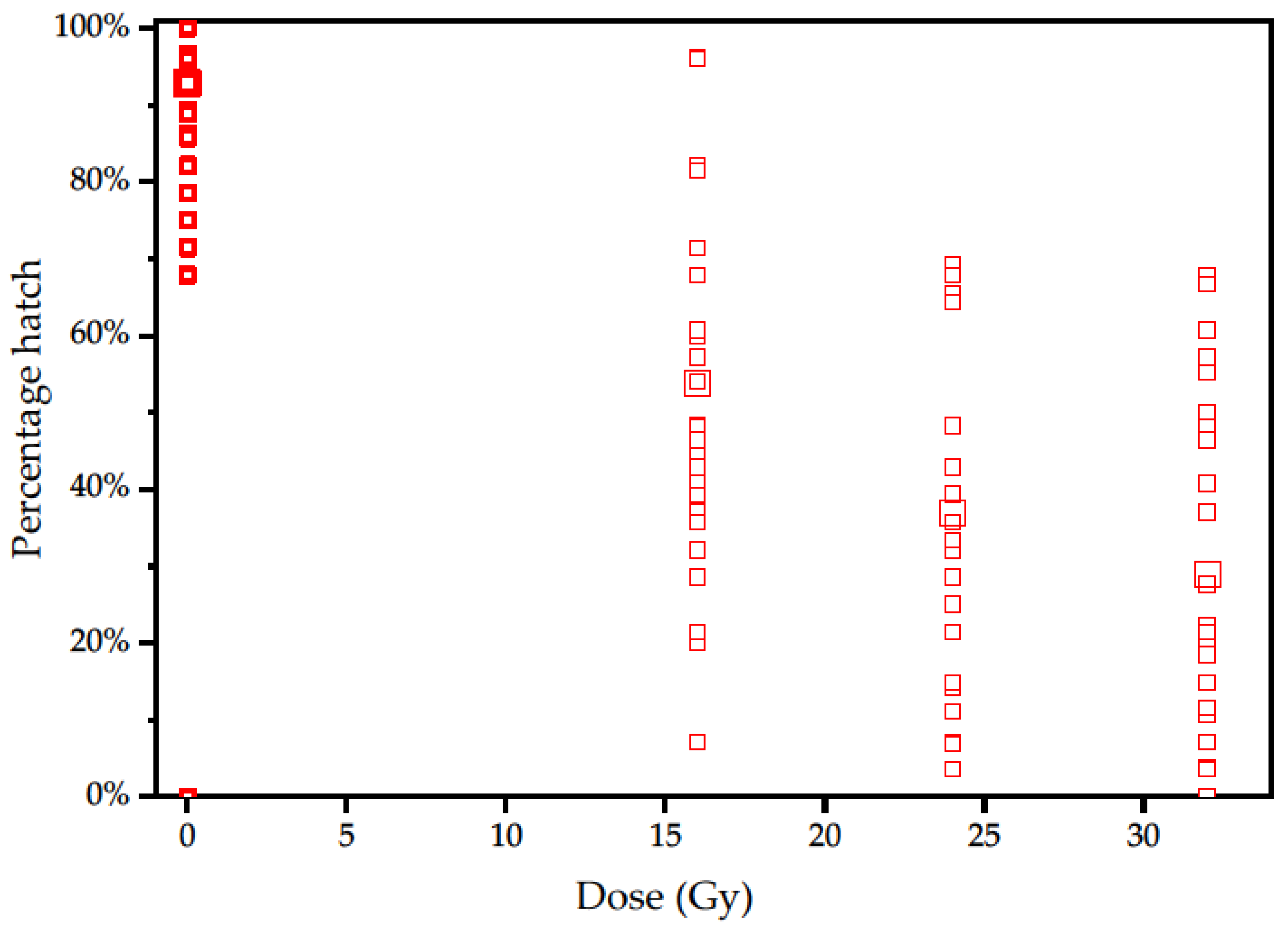
3.1. Experiment 1
3.2. Experiment 2. Overflooding with Older and Virgin Males
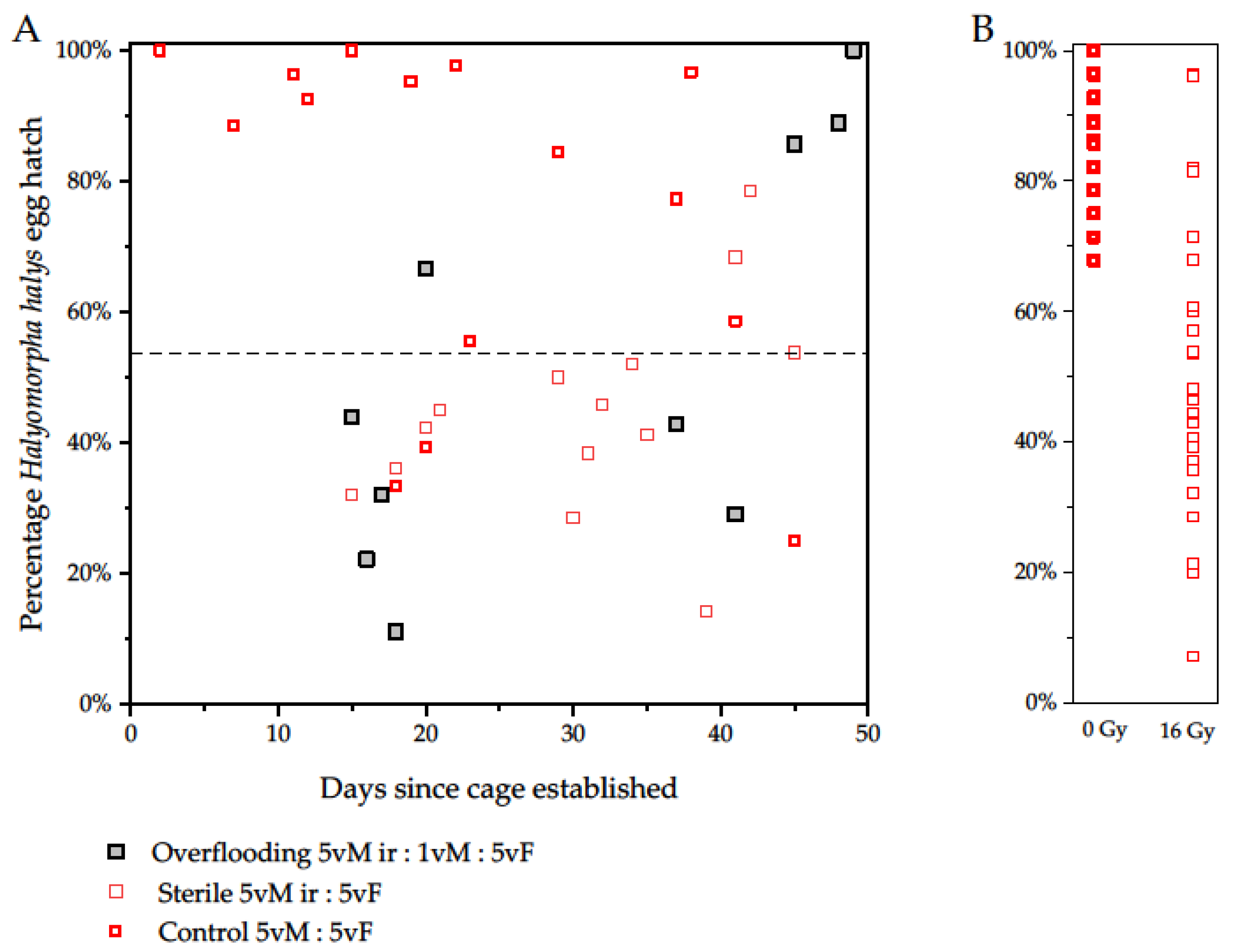
3.3. Experiment 3. Overflooding and Egg Sterility over Time
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kriticos, D.J.; Kean, J.M.; Phillips, C.B.; Senay, S.D.; Acosta, H.; Haye, T. The potential global distribution of the brown marmorated stink bug, Halyomorpha halys, a critical threat to plant biosecurity. J. Pest Sci. 2017, 90, 1033–1043. [Google Scholar] [CrossRef]
- Haye, T.; Gariepy, T.; Hoelmer, K.; Rossi, J.-P.; Streito, J.-C.; Tassus, X.; Desneux, N. Range expansion of the invasive brown marmorated stinkbug, Halyomorpha halys: An increasing threat to field, fruit and vegetable crops worldwide. J. Pest Sci. 2015, 88, 665–673. [Google Scholar] [CrossRef]
- Lee, D.H.; Short, B.D.; Joseph, S.V.; Bergh, J.C.; Leskey, T.C. Review of the biology, ecology, and management of Halyomorpha halys (Hemiptera: Pentatomidae) in China, Japan, and the Republic of Korea. Environ. Entomol. 2013, 42, 627–641. [Google Scholar] [CrossRef] [PubMed]
- Bergmann, E.; Bernhard, K.M.; Bernon, G.; Bickerton, M.; Gill, S.; Gonzales, C.; Hamilton, G.C.; Hedstrom, C.; Kamminga, K.; Koplinka-Loehr, C.; et al. Host Plants of the Brown Marmorated Stink Bug in the United States. Available online: http://www.stopbmsb.org/where-is-bmsb/host-plants/#host_plants_table (accessed on 15 July 2016).
- Malek, R.; Tattoni, C.; Ciolli, M.; Corradini, S.; Andreis, D.; Ibrahim, A.; Mazzoni, V.; Eriksson, A.; Anfora, G. Coupling traditional monitoring and citizen science to disentangle the invasion of Halyomorpha halys. ISPRS Int. J. Geo-Inf. 2018, 7, 171. [Google Scholar] [CrossRef]
- Lee, D.H.; Wright, S.E.; Leskey, T.C. Impact of insecticide residue exposure on the invasive pest, Halyomorpha halys (Hemiptera: Pentatomidae): Analysis of adult mobility. J. Econ. Entomol. 2013, 106, 150–158. [Google Scholar] [CrossRef] [PubMed]
- Knight, K.M.M.; Gurr, G.M. Review of Nezara viridula (L.) management strategies and potential for IPM in field crops with emphasis on Australia. Crop Prot. 2007, 26, 1–10. [Google Scholar] [CrossRef]
- Žunič, A.; Čokl, A.; Serša, G. Effects of 5-Gy irradiation on fertility and mating behaviour of Nezara viridula (Heteroptera: Pentatomidae). Radiol. Oncol. 2002, 36, 231–237. [Google Scholar]
- Mazzoni, V.; Polajnar, J.; Baldini, M.; Rossi Stacconi, M.V.; Anfora, G.; Guidetti, R.; Maistrello, L. Use of substrate-borne vibrational signals to attract the Brown Marmorated Stink Bug, Halyomorpha halys. J. Pest Sci. 2017, 90, 1219–1229. [Google Scholar] [CrossRef]
- Čokl, A.A.; Millar, J.G. Manipulation of insect signaling for monitoring and control of pest insects. In Biorational Control of Arthropod Pests: Application and Resistance Management; Ishaaya, I., Horowitz, A.R., Eds.; Springer Netherlands: Dordrecht, The Netherlands, 2009; pp. 279–316. [Google Scholar]
- Khrimian, A.; Zhang, A.; Weber, D.C.; Ho, H.-Y.; Aldrich, J.R.; Vermillion, K.E.; Siegler, M.A.; Shirali, S.; Guzman, F.; Leskey, T.C. Discovery of the aggregation pheromone of the brown marmorated stink bug (Halyomorpha halys) through the creation of stereoisomeric libraries of 1-bisabolen-3-ols. J. Nat. Prod. 2014, 77, 1708–1717. [Google Scholar] [CrossRef]
- Weber, D.C.; Leskey, T.C.; Walsh, G.C.; Khrimian, A. Synergy of aggregation pheromone with methyl (E,E,Z)-2,4,6-decatrienoate in attraction of Halyomorpha halys (Hemiptera: Pentatomidae). J. Econ. Entomol. 2014, 107, 1061–1068. [Google Scholar] [CrossRef]
- Klassen, W.; Curtis, C.F. History of the sterile insect technique. In Sterile Insect Technique: Principles and Practice in Area-Wide Integrated Pest Management; Dyck, V.A., Hendrichs, J., Robinson, A.S., Eds.; Springer: Dordrecht, The Netherlands, 2005; pp. 3–36. [Google Scholar]
- Knipling, E.F. Possibilities of insect control or eradication through the use of sexually sterile males. J. Econ. Entomol. 1955, 48, 459–462. [Google Scholar] [CrossRef]
- Ameresekere, R.V.W.E.; Georghiou, G.P.; Sevacherian, V. Histopathological studies on irradiated and chemosterilized beet leafhoppers, Circulifer tenellus. Ann. Entomol. Soc. Am. 1971, 64, 1025–1031. [Google Scholar] [CrossRef]
- Dyby, S.D.; Sailer, R.I. Impact of low-level radiation on fertility and fecundity of Nezara viridula (Hemiptera:Pentatomidae). J. Econ. Entomol. 1999, 92, 945–953. [Google Scholar] [CrossRef]
- Mau, R.; Mitchell, W.C.; Anwar, M. Preliminary studies on the effects of gamma irradiation of eggs and adults of the southern green stink bug, Nezara viridula (L.). Proc. Hawaii Entomol. Soc. 1967, 19, 415–417. [Google Scholar]
- Sales, F.M. Effects of gamma radiation on eggs of southern green stink bug, Nezara viridula L. Fitossanidade 1977, 2, 17–18. [Google Scholar]
- Stringer, L.D.; Harland, D.P.; Grant, J.E.; Laban, J.; Suckling, D.M. Effect of 40 Gy irradiation on the ultrastructure, biochemistry, morphology and cytology during spermatogenesis in the southern green stink bug Nezara viridula (Hemiptera: Pentatomidae). bioRxiv 2017, 17, 1991. [Google Scholar]
- LaChance, L.E.; Degrugillie, M. Chromosomal fragments transmitted through three generations in Oncopeltus (Hemiptera). Science 1969, 166, 235–236. [Google Scholar] [CrossRef]
- Kawada, H.; Kitamura, C. The reproductive behavior of the brown marmorated stink bug, Halyomorpha mista UHLER (Heteroptera: Pentatomidae) I. Observation of the mating behavior and multiple copulation. Appl. Entomol. Zool. 1983, 18, 234–242. [Google Scholar] [CrossRef]
- Knipling, E.F. The Basic Principles of Insect Population Suppression and Management; Agricultural Handbook 512; USDA/SEA: Washington, DC, USA, 1979; p. 659.
- Woods, B.; McInnis, D.O.; Steiner, E.; Soopaya, A.; Lindsey, J.; Lacey, I.; Virdi, A.; Fogliani, R. Developing field cage tests to measure mating competitiveness of sterile light brown apple moths (Lepidoptera: Tortricidae) in Western Australia. Fla. Entomol. 2016, 99, 138–145. [Google Scholar] [CrossRef]
- Lance, D.; McInnis, D. Biological basis of the sterile insect technique. In Sterile Insect Technique; Dyck, V.A., Hendrichs, J., Robinson, A.S., Eds.; Springer: Berlin, Germany, 2005; pp. 69–94. [Google Scholar]
- Cáceres, C.; McInnis, D.; Shelly, T.; Jang, E.; Robinson, A.; Hendrichs, J. Quality management systems for fruit fly (Diptera: Tephritidae) sterile insect technique. Fla. Entomol. 2007, 90, 1–9. [Google Scholar] [CrossRef]
- Welsh, T.J.; Stringer, L.D.; Caldwell, R.; Carpenter, J.E.; Suckling, D.M. Irradiation biology of male brown marmorated stink bugs: Is there scope for the sterile insect technique? Int. J. Radiat. Biol. 2017, 93, 1357–1363. [Google Scholar] [CrossRef] [PubMed]
- Baccaro, S.; Cemmi, A.; Sarcina, I.D.; Ferrara, G. Gamma irradiation Calliope facility at ENEA-Casaccia Research Centre (Rome, Italy); Fusion and Technology for Nuclear Safety and Security Department Casaccia Research Centre, Ed.; Italian National Agency For New Technologies, Energy And Sustainable Economic Development, Casaccia Research Centre: Rome, Italy, 2019; Volume RT/2019/4/ENEA, p. 49. [Google Scholar]
- Akers, M.D. Exploring, Analysing and Interpeting Data with Minitab 18, 1st ed.; Book Refinery Limited: Portsmouth, UK, 2018. [Google Scholar]
- Abbott, W.S. A method of computing the effectiveness of an insecticide. J. Econ. Entomol. 1925, 18, 265–267. [Google Scholar] [CrossRef]
- Bakri, A.; Mehta, K.; Lance, D.R. Chapter 3.3. Sterilizing insects with ionizing radiation. In Sterile Insect Technique Principles and Practice in Area-Wide Integrated Pest Management; Dyck, V.A., Hendrichs, J., Robinson, A.S., Eds.; International Atomic Energy Agency: Vienna, Austria, 2005; p. 779. [Google Scholar]
- Vreysen, M.J.B. Monitoring sterile and wild insects in area-wide integrated pest management programmes. In Sterile Insect Technique: Principles and Practice in Area-Wide Integrated Pest Management; Springer: Dordrecht, The Netherlands, 2005; pp. 325–361. [Google Scholar]
- Suckling, D.M.; Conlong, D.E.; Carpenter, J.E.; Bloem, K.A.; Rendon, P.; Vreysen, M.J.B. Global range expansion of pest Lepidoptera requires socially acceptable solutions. Biol. Invasions 2016, 19, 1107–1119. [Google Scholar] [CrossRef]
- Meats, A.W.; Duthie, R.; Clift, A.D.; Dominiak, B.C. Trials on variants of the Sterile Insect Technique (SIT) for suppression of populations of the Queensland fruit fly in small towns neighbouring a quarantine (exclusion) zone. Aust. J. Exp. Agric. 2003, 43, 389–395. [Google Scholar] [CrossRef]
- McInnis, D.O.; Hendrichs, J.; Shelly, T.; Barr, N.; Hoffman, K.; Rodriguez, R.; Lance, D.R.; Bloem, K.; Suckling, D.M.; Enkerlin, W.; et al. Can polyphagous invasive tephritid pest populations escape detection for years under favorable climatic and host conditions? Am. Entomol. 2017, 63, 89–99. [Google Scholar] [CrossRef]
- Steiner, L.F.; Harris, E.J.; Mitchell, W.C.; Fujimoto, M.S.; Christenson, L.D. Melon fly eradication by overflooding with sterile flies. J. Econ. Entomol. 1965, 58, 519–522. [Google Scholar] [CrossRef]
- Suckling, D.M.; Barrington, A.M.; Chhagan, A.; Stephens, A.E.A.; Burnip, G.M.; Charles, J.G.; Wee, S.L. Eradication of the Australian Painted Apple Moth Teia anartoides in New Zealand: Trapping, Inherited Sterility, and Male Competitiveness. In Area-Wide Control of Insect Pests: From Research to Field Implementation; Vreysen, M.J.B., Robinson, A.S., Hendrichs, J., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 603–615. [Google Scholar]
- Walker, M.L.; McLaughlin, W.L.; Puhl, J.M.; Gomes, P. Radiation-field mapping of insect irradiation canisters. Appl. Radiat. Isot. 1997, 48, 117–125. [Google Scholar] [CrossRef]
- Paoli, F.; Dallai, R.; Cristofaro, M.; Arnone, S.; Francardi, V.; Roversi, P.F. Morphology of the male reproductive system, sperm ultrastructure and γ-irradiation of the red palm weevil Rhynchophorus ferrugineus Oliv. (Coleoptera: Dryophthoridae). Tissue Cell 2014, 46, 274–285. [Google Scholar] [CrossRef]
- Soopaya, R.; Stringer, L.D.; Woods, B.; Stephens, A.E.A.; Butler, R.C.; Lacey, I.; Kaur, A.; Suckling, D.M. Radiation biology and inherited sterility of light brown apple moth (Lepidoptera: Tortricidae): Developing a sterile insect release program. J. Econ. Entomol. 2011, 104, 1999–2008. [Google Scholar] [CrossRef]
- Jang, E.B.; McInnis, D.O.; Kurashima, R.; Woods, B.; Suckling, D.M. Irradiation of adult Epiphyas postvittana (Lepidoptera: Tortricidae): Egg sterility in parental and F1 generations. J. Econ. Entomol. 2012, 105, 54–61. [Google Scholar] [CrossRef]
- Balestrino, F.; Medici, A.; Candini, G.; Carrieri, M.; MacCagnani, B.; Calvitti, M.; Maini, S.; Bellini, R. γ ray dosimetry and mating capacity studies in the laboratory on Aedes albopictus males. J. Med. Entomol. 2010, 47, 511, 581–591. [Google Scholar] [CrossRef] [PubMed]
- Horber, E. Eradication of the white grub (Melolontha vulgaris F.) by the sterile male technique. In Radiation and Radioisotopes Applied to Insects of Agricultural Importance, Proceedings of the Symposium on the Use and Application of Radioisotopes and Radiation in the Control of Plant and Animal Insect Pests, Athens, Greece, 22–26 April 1963; IAEA: Vienna, Austria, 1963; pp. 313–332. [Google Scholar]
- Boller, E.F.; Remund, U. Application of SIT on the European Cherry Fruit Fly, Rhagoletis Cerasi l., in Northwest Switzerland; IAEA-PL-582/7; International Atomic Energy Agency (IAEA): Vienna, Austria, 1975. [Google Scholar]
- Suckling, D.M.; Levy, M.C.; Roselli, G.; Mazzoni, V.; Ioriatti, C.; Deromedi, M.; Cristofaro, M.; Anfora, G. Live traps for adult brown marmorated stink bugs. Insects 2019, 10, 376. [Google Scholar] [CrossRef] [PubMed]
- Taylor, C.M.; Coffey, P.L.; Hamby, K.A.; Dively, G.P. Laboratory rearing of Halyomorpha halys: Methods to optimize survival and fitness of adults during and after diapause. J. Pest Sci. 2017, 90, 1069. [Google Scholar] [CrossRef]
- Cira, T.M.; Venette, R.C.; Aigner, J.; Kuhar, T.; Mullins, D.E.; Gabbert, S.E.; Hutchison, W.D. Cold tolerance of Halyomorpha halys (Hemiptera: Pentatomidae) across geographic and temporal scales. Environ. Entomol. 2016, 45, 484–491. [Google Scholar] [CrossRef]
- Machi, A.R.; Rodrigues Mayne, R.; Gava, M.A.; Arthur, P.B.; Arthur, V. Gamma radiation sterilization dose of adult males in asian tiger mosquito pupae. Insects 2019, 10, 101. [Google Scholar] [CrossRef]
- Garcia-Gonzales, F.; Simmons, L. Sperm viability matters in insect sperm competition. Curr. Biol. 2005, 15, 271–275. [Google Scholar] [CrossRef]
- Kean, J.M.; Stephens, A.E.A.; Wee, S.L.; Suckling, D.M. Optimizing strategies for eradication of discrete-generation lepidopteran pests using inherited sterility. In Area-Wide Control of Insect Pests: From reearch to fied impleentation; Vreysen, M.J.B., Robinson, A.S., Hendrichs, J., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 211–220. [Google Scholar]
- Suckling, D.M.; Tobin, P.C.; McCullough, D.G.; Herms, D.A. Combining tactics to exploit Allee effects for eradication of alien insect populations. J. Econ. Entomol. 2012, 105, 1–13. [Google Scholar] [CrossRef]
- Parker, A.; Mehta, K. Sterile insect technique: A model for dose optimization for improved sterile insect quality. Fla. Entomol. 2007, 90, 88–95. [Google Scholar] [CrossRef]
- Taylor, C.M.; Coffey, P.L.; DeLay, B.D.; Dively, G.P. The importance of gut symbionts in the development of the brown marmorated stink bug, Halyomorpha halys (Stål). PLoS ONE 2014, 9, e90312. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Gy | Batches | Eggs Per Batch | Eggs Laid | Hatched | Percentage Hatch | Corrected Hatch |
|---|---|---|---|---|---|---|
| 0 | 58 | 26 (25–27) | 1480 | 1360 | 92% | |
| 16 | 53 | 25 (23–26) | 1319 | 664 | 50% | 54.4% |
| 24 | 36 | 26 (24–28) | 853 | 287 | 34% | 37.0% |
| 32 | 36 | 24 (22–26) | 871 | 231 | 27% | 29.4% |
| Treatment | Male Type | Egg Status | Number of Batches | Total Eggs | Observed Hatch | Expected Hatch | X2 | p |
|---|---|---|---|---|---|---|---|---|
| Virgin males (5:1) | ||||||||
| 16 Gy | 5 ir males:1 male:5 virgin females | Laid | 21 | 557 | 42% | 62% 1 | 96.2 | <0.0001 |
| Hatched | 233 | |||||||
| Control | 5 males:5 virgin females | Laid | 4 | 72 | 85% | 100% | ||
| Hatched | 61 | |||||||
| Overwintered males (5:1) | ||||||||
| 16 Gy | 5 ir males:1 male:5 virgin females | Laid | 26 | 447 | 61% 2 | 62% 1 | 0.09 | 0.75 |
| Hatched | 274 | |||||||
| Control | 5 males:5 virgin females | Laid | 14 | 295 | 94% | 100% | ||
| Hatched | 277 | |||||||
| Treatment Using Virgin Males and Females | Number of Batches | Total Eggs | Eggs Hatched | Observed Hatch % | Expected Hatch % | X2 | p |
|---|---|---|---|---|---|---|---|
| 5 ir males:1 male:5 females | 18 | 398 | 207 | 52.0 | 62 1 | 16.86 | 0.0001 |
| 5 ir males:5 females | 15 | 373 | 148 | 39.7 | 54 2 | 30.80 | 0.0001 |
| 5 males:5 females | 33 | 794 | 646 | 81.4 | 100 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suckling, D.M.; Cristofaro, M.; Roselli, G.; Levy, M.C.; Cemmi, A.; Mazzoni, V.; Stringer, L.D.; Zeni, V.; Ioriatti, C.; Anfora, G. The Competitive Mating of Irradiated Brown Marmorated Stink Bugs, Halyomorpha halys, for the Sterile Insect Technique. Insects 2019, 10, 411. https://doi.org/10.3390/insects10110411
Suckling DM, Cristofaro M, Roselli G, Levy MC, Cemmi A, Mazzoni V, Stringer LD, Zeni V, Ioriatti C, Anfora G. The Competitive Mating of Irradiated Brown Marmorated Stink Bugs, Halyomorpha halys, for the Sterile Insect Technique. Insects. 2019; 10(11):411. https://doi.org/10.3390/insects10110411
Chicago/Turabian StyleSuckling, David Maxwell, Massimo Cristofaro, Gerardo Roselli, Mary Claire Levy, Alessia Cemmi, Valerio Mazzoni, Lloyd Damien Stringer, Valeria Zeni, Claudio Ioriatti, and Gianfranco Anfora. 2019. "The Competitive Mating of Irradiated Brown Marmorated Stink Bugs, Halyomorpha halys, for the Sterile Insect Technique" Insects 10, no. 11: 411. https://doi.org/10.3390/insects10110411
APA StyleSuckling, D. M., Cristofaro, M., Roselli, G., Levy, M. C., Cemmi, A., Mazzoni, V., Stringer, L. D., Zeni, V., Ioriatti, C., & Anfora, G. (2019). The Competitive Mating of Irradiated Brown Marmorated Stink Bugs, Halyomorpha halys, for the Sterile Insect Technique. Insects, 10(11), 411. https://doi.org/10.3390/insects10110411