
Dairy Product Consumption Interacts with Glucokinase (GCK) Gene Polymorphisms Associated with Insulin Resistance

 ,
,  , and
, and 
Abstract
1. Introduction
2. Results
2.1. Participants’ Characteristics
2.2. Interactions between Glucokinase Genotypes and Dairy Intake
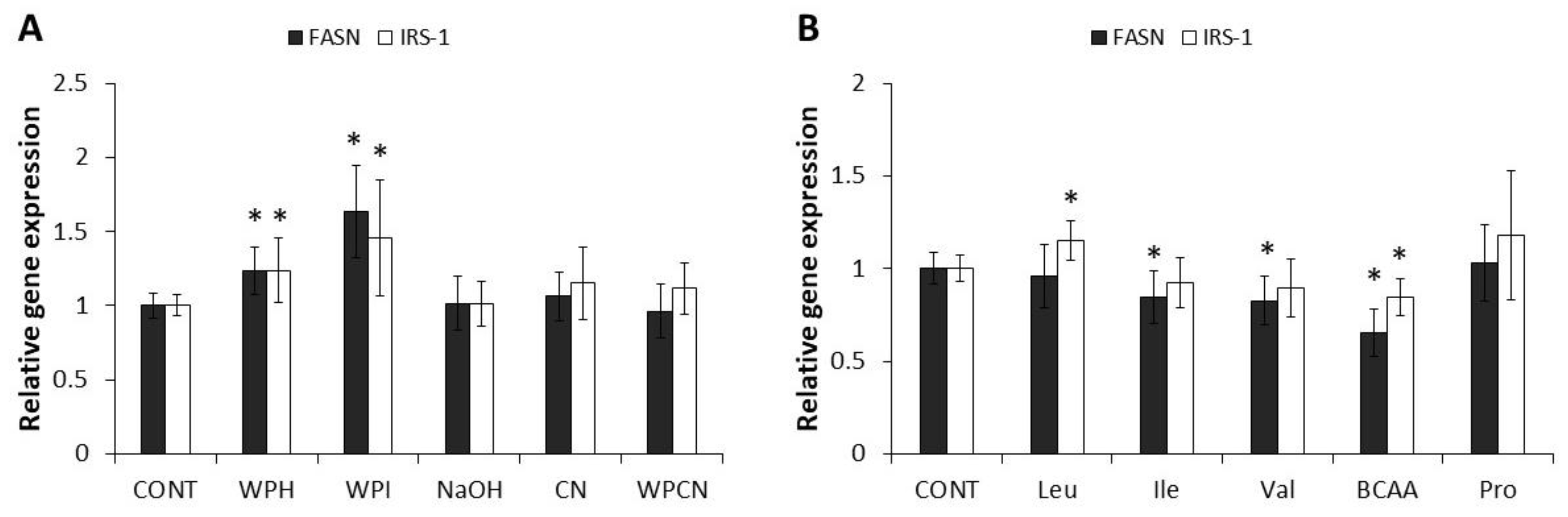
2.3. Effect of Dairy Proteins and Amino Acids on Glucose Metabolism Genes in HepG2 Cells
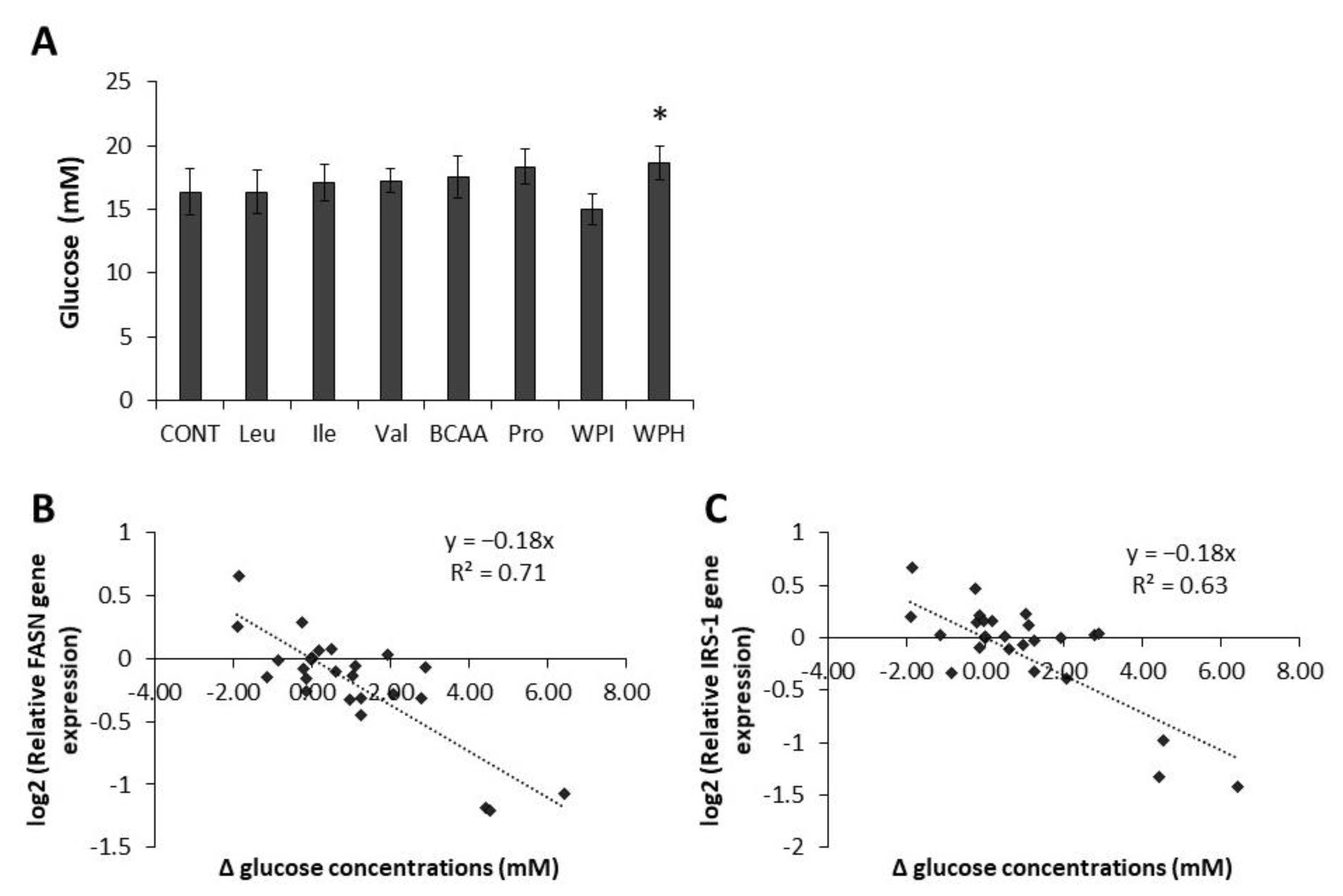
2.4. Glucose Concentrations in Cell Supernatants are Linked to FASN and IRS-1 Gene Expression
3. Discussion
4. Materials and Methods
4.1. Human Study
4.1.1. Study Population
4.1.2. Anthropometric Measurements
4.1.3. Dietary Intake
4.1.4. Biochemical Parameters
4.1.5. DNA Extraction and Genotyping
4.2. In Vitro Study
4.2.1. Materials
4.2.2. Cell Culture and Treatment
4.2.3. Gene Expression Analysis
4.2.4. Glucose Concentrations in Cell Supernatants
4.2.5. Cell Viability Assay
4.3. Statistical Analyses
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Gijsbers, L.; Ding, E.L.; Malik, V.S.; De Goede, J.; Geleijnse, J.M.; Soedamah-Muthu, S.S. Consumption of dairy foods and diabetes incidence: A dose-response meta-analysis of observational studies 1, 2. Am. J. Clin. Nutr. 2016, 103, 1111–1124. [Google Scholar] [CrossRef] [PubMed]
- Turner, K.M.; Keogh, J.B.; Clifton, P.M. Dairy consumption and insulin sensitivity: A systematic review of short- and long-term intervention studies. Nutr. Metab. Cardiovasc. Dis. 2015, 25, 3–8. [Google Scholar] [CrossRef] [PubMed]
- Barroso, I. Genetics of type 2 diabetes. Diabet. Med. 2005, 22, 517–535. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, M.I. Genomics, type 2 diabetes, and obesity. N. Engl. J. Med. 2010, 363, 2339–2350. [Google Scholar] [CrossRef] [PubMed]
- Muller, Y.L.; Piaggi, P.; Hoffman, D.; Huang, K.; Gene, B.; Kobes, S.; Thearle, M.S.; Knowler, W.C.; Hanson, R.L.; Baier, L.J.; et al. Common genetic variation in the glucokinase gene (GCK) is associated with type 2 diabetes and rates of carbohydrate oxidation and energy expenditure. Diabetologia 2014, 57, 1382–1390. [Google Scholar] [CrossRef] [PubMed]
- Rosta, K.; Al-Aissa, Z.; Hadarits, O.; Harreiter, J.; Nádasdi, Á.; Kelemen, F.; Bancher-Todesca, D.; Komlósi, Z.; Németh, L.; Rigó, J.; et al. Association study with 77 SNPs confirms the robust role for the rs10830963/G of MTNR1B variant and identifies two novel associations in gestational Diabetes Mellitus development. PLoS ONE 2017, 12, e0169781. [Google Scholar] [CrossRef] [PubMed]
- Raibert, M.; Playter, R.; Krummel, T. The use of a virtual reality haptic device in surgical training. Acad. Med. 1998, 73, 596–597. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Cui, H.; Chen, X.; Xie, W.; Chang, Y. Association of the glucokinase gene promoter polymorphism −30G >A (rs1799884) with gestational diabetes mellitus susceptibility: A case-control study and meta-analysis. Arch. Gynecol. Obstet. 2015, 292, 291–298. [Google Scholar] [CrossRef] [PubMed]
- Haeusler, R.A.; Camastra, S.; Astiarraga, B.; Nannipieri, M.; Anselmino, M.; Ferrannini, E. Decreased expression of hepatic glucokinase in type 2 diabetes. Mol. Metab. 2015, 4, 222–226. [Google Scholar] [CrossRef] [PubMed]
- Higuchi, N.; Kato, M.; Miyazaki, M.; Tanaka, M.; Kohjima, M.; Ito, T.; Nakamuta, M.; Enjoji, M.; Kotoh, K.; Takayanagi, R. Potential role of branched-chain amino acids in glucose metabolism through the accelerated induction of the glucose-sensing apparatus in the liver. J. Cell. Biochem. 2011, 112, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Pal, S.; Radavelli-Bagatini, S. The effects of whey protein on cardiometabolic risk factors. Obes. Rev. 2013, 14, 324–343. [Google Scholar] [CrossRef] [PubMed]
- Pasin, G.; Comerford, K.B. Dairy Foods and Dairy Proteins in the Management of Type 2 Diabetes: A Systematic Review of the Clinical Evidence. Adv. Nutr. 2015, 6, 245–259. [Google Scholar] [CrossRef] [PubMed]
- Mignone, L.E.; Wu, T.; Horowitz, M.; Rayner, C.K. Whey protein: The “whey” forward for treatment of type 2 diabetes? World J. Diabetes 2015, 6, 1274–1284. [Google Scholar] [CrossRef] [PubMed]
- Bruckbauer, A.; Gouffon, J.; Rekapalli, B.; Zemel, M.B. The effects of dairy components on energy partitioning and metabolic risk in mice: A microarray study. J. Nutrigenet. Nutrigenom. 2009, 2, 64–77. [Google Scholar] [CrossRef] [PubMed]
- Bouchard-Mercier, A.; Rudkowska, I.; Lemieux, S.; Couture, P.; Vohl, M.C. An interaction effect between glucokinase gene variation and carbohydrate intakes modulates the plasma triglyceride response to a fish oil supplementation. Genes Nutr. 2014, 9, 395. [Google Scholar] [CrossRef] [PubMed]
- Gloyn, A.L. Glucokinase (GCK) mutations in hyper- and hypoglycemia: Maturity-onset diabetes of the young, permanent neonatal diabetes, and hyperinsulinemia of infancy. Hum. Mutat. 2003, 22, 353–362. [Google Scholar] [CrossRef] [PubMed]
- Sotos-Prieto, M.; Guillén, M.; Vicente Sorli, J.; Portolés, O.; Guillem-Saiz, P.; Ignacio Gonzalez, J.; Qi, L.; Corella, D. Relevant associations of the glucokinase regulatory protein/glucokinase gene variation with TAG concentrations in a high-cardiovascular risk population: Modulation by the Mediterranean diet. Br. J. Nutr. 2012, 109, 193–201. [Google Scholar] [CrossRef] [PubMed]
- Yamada, K.; Yuan, X.; Ishiyama, S.; Ichikawa, F.; Koyama, K.-I.; Koyanagi, A.; Koyama, W.; Nonaka, K. Clinical characteristics of Japanese men with glucokinase gene beta-cell promoter variant. Diabetes Care 1997, 20, 1996–1998. [Google Scholar] [CrossRef]
- Rose, C.S.; Ek, J.; Urhammer, S.A.; Glu, C.; Borch-johnsen, K. A −30G >A polymorphism of the beta-cell-specific glucokinase promoter associates with hyperglycemia in the general population of whites. Diabetes 2005, 54, 3026–3031. [Google Scholar] [CrossRef] [PubMed]
- März, W.; Nauck, M.; Hoffmann, M.M.; Nagel, D.; Boehm, B.O.; Koenig, W.; Rothenbacher, D.; Winkelmann, B.R. G(−30)A polymorphism in the pancreatic promoter of the glucokinase gene associated with angiographic coronary artery disease and type 2 diabetes mellitus. Circulation 2004, 109, 2844–2849. [Google Scholar] [CrossRef] [PubMed]
- Pal, M. Recent advances in glucokinase activators for the treatment of type 2 diabetes. Drug Discov. Today 2009, 14, 784–792. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.; Ley, S.H.; Zheng, Y.; Wang, T.; Bray, G.A.; Sacks, F.M.; Qi, L. Genetic susceptibility to diabetes and long-term improvement of insulin resistance and beta-cell function during weight loss: The Preventing Overweight Using Novel Dietary Strategies (POUNDS LOST) trial. Am. J. Clin. Nutr. 2016, 104, 198–204. [Google Scholar] [CrossRef] [PubMed]
- Pereira, P.C. Milk nutritional composition and its role in human health. Nutrition 2014, 30, 619–627. [Google Scholar] [CrossRef] [PubMed]
- Rudkowska, I.; Julien, P.; Couture, P.; Lemieux, S.; Tchernof, A.; Barbier, O.; Vohl, M. Cardiometabolic risk factors are influenced by Stearoyl-CoA Desaturase (SCD)-1 gene polymorphisms and n-3 polyunsaturated fatty acid supplementation. Mol. Nutr. Food Res. 2014, 58, 1079–1086. [Google Scholar] [CrossRef] [PubMed]
- Cormier, H.; Rudkowska, I.; Lemieux, S.; Couture, P.; Julien, P.; Vohl, M.-C. Effects of FADS and ELOVL polymorphisms on indexes of desaturase and elongase activities: Results from a pre-post fish oil supplementation. Genes Nutr. 2014, 9, 437. [Google Scholar] [CrossRef] [PubMed]
- Callaway, C.; Chumlea, W.; Bouchard, C.; Himes, J.; Lohman, T.; Martin, A.; Mitchell, C.; Mueller, W.; Roche, A.; Seefeldt, V. Standardization of anthropometric measurements. In The Airlie (VA) Consensus Conference; Lohman, T., Roche, A., Martorel, R., Champaign, I., Eds.; Human Kinetics Publishers: Champaign, IL, USA, 1988; pp. 39–80. [Google Scholar]
- Goulet, J.; Nadeau, G.; Lapointe, A.; Lamarche, B.; Lemieux, S. Validity and reproducibility of an interviewer-administered food frequency questionnaire for healthy French-Canadian men and women. Nutr. J. 2004, 3, 13. [Google Scholar] [CrossRef] [PubMed]
- Desbuquois, B.; Aurbach, G.D. Use of polyethylene glycol to separate free and antibody-bound peptide hormones in radioimmunoassays. J. Clin. Endocrinol. Metab. 1971, 33, 732–738. [Google Scholar] [CrossRef] [PubMed]
- Richterich, R.; Dauwalder, H. Determination of plasma glucose by hexokinase-glucose-6-phosphate dehydrogenase method. Schweiz. Med. Wochenschr. 1971, 101, 615–618. [Google Scholar] [PubMed]
- Matthews, D.R.; Hosker, J.P.; Rudenski, A.S.; Naylor, B.A.; Treacher, D.F.; Turner, R.C. Homeostasis model assessment: Insulin resistance and beta-cell function from fasting plasma glucose and insulin concentrations in man. Diabetologia 1985, 28, 412–419. [Google Scholar] [CrossRef] [PubMed]
- Leys, C.; Ley, C.; Klein, O.; Bernard, P.; Licata, L. Detecting outliers: Do not use standard deviation around the mean, use absolute deviation around the median. J. Exp. Soc. Psychol. 2013, 49, 764–766. [Google Scholar] [CrossRef]
- Da Silva, M.S.; Rudkowska, I. Dairy nutrients and their effect on inflammatory profile in molecular studies. Mol. Nutr. Food Res. 2015, 59, 1249–1263. [Google Scholar] [CrossRef] [PubMed]
- Hoefle, A.S.; Bangert, A.M.; Stamfort, A.; Gedrich, K.; Rist, M.J.; Lee, Y.-M.; Skurk, T.; Daniel, H. Metabolic responses of healthy or prediabetic adults to bovine whey protein and sodium caseinate do not differ. J. Nutr. 2015, 145, 467–475. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Gayoso-Diz, P.; Otero-González, A.; Rodriguez-Alvarez, M.X.; Gude, F.; García, F.; De Francisco, A.; Quintela, A.G. Insulin resistance (HOMA-IR) cut-off values and the metabolic syndrome in a general adult population: Effect of gender and age: EPIRCE cross-sectional study. BMC Endocr. Disord. 2013, 13, 47. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| tSNP | Localisation | Genotype (n) | p-Values 1 | ||
|---|---|---|---|---|---|
| Genotype | Dairy | Interaction | |||
| rs2268573 C > A MAF = 0.39 | Intron | CC (39) AC (111) AA (60) | 0.93 | 0.91 | 0.24 |
| rs2908297 G > A MAF = 0.31 | Intron | GG (167) AG (36) + AA (7) | 0.94 | 0.99 | 0.96 |
| rs2971676 C > T MAF = 0.10 | Intron | CC (183) CT (24) + TT (3) | 0.94 | 0.68 | 0.60 |
| rs758989 A > G MAF = 0.37 | Intron | AA (62) AG (96) GG (52) | 0.62 | 0.90 | 0.10 |
| rs12673242 T > C MAF = 0.24 | Intron | TT (171) CT (36) + CC (3) | 0.27 | 0.59 | 0.53 |
| rs2908290 C > T MAF = 0.45 | Intron | CC (102) CT (88) TT (20) | 0.84 | 0.43 | 0.34 |
| rs2284777 G > A MAF = 0.12 | Intron | AA (152) AG (50) + GG (8) | 0.89 | 0.70 | 0.30 |
| rs2300584 T > C MAF = 0.19 | Intron | TT (134) CT (70) + CC (6) | 0.97 | 0.75 | 0.21 |
| rs1990458 G > A MAF = 0.34 | Intron | GG (79) AG (100) AA (31) | 0.59 | 0.69 | 0.69 |
| rs741038 C > T MAF = 0.28 | Intron | TT (93) CT (94) CC (23) | 0.97 | 0.38 | 0.08 |
| rs1799884 G > A MAF = 0.17 | 5′UTR | GG (135) AG (65) + AA (10) | 0.93 | 0.54 | 0.02 |
| rs2908277 C > T MAF = 0.20 | 3′UTR | CC (166) CT (41) + TT (3) | 0.78 | 0.80 | 0.67 |
| rs3757838 A > T MAF = 0.05 | Promoter | TT (186) AT (21) + AA (3) | 0.53 | 0.59 | 0.49 |
| Genotype | GG (n = 135) | AG (n = 65) + AA (n = 10) | p-Values | ||||
|---|---|---|---|---|---|---|---|
| Dairy intake | Low (n = 66) | High (n = 68) | Low (n = 38) | High (n = 37) | G | D | G × D |
| Fasting glucose | 4.98 ± 0.42 | 4.89 ± 0.48 | 5.11 ± 0.46 | 4.85 ± 0.45 | 0.30 | 0.21 | 0.33 |
| Fasting insulin | 79.25 ± 37.44 | 84.91 ± 35.58 | 88.24 ± 49.75 | 70.78 ± 27.76 | 0.66 | 0.89 | 0.035 |
| HOMA-IR | 2.54 ± 1.27 | 2.65 ± 1.09 | 2.88 ± 1.60 | 2.21 ± 0.88 | 0.93 | 0.54 | 0.02 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Da Silva, M.S.; Chartrand, D.; Vohl, M.-C.; Barbier, O.; Rudkowska, I. Dairy Product Consumption Interacts with Glucokinase (GCK) Gene Polymorphisms Associated with Insulin Resistance. J. Pers. Med. 2017, 7, 8. https://doi.org/10.3390/jpm7030008
Da Silva MS, Chartrand D, Vohl M-C, Barbier O, Rudkowska I. Dairy Product Consumption Interacts with Glucokinase (GCK) Gene Polymorphisms Associated with Insulin Resistance. Journal of Personalized Medicine. 2017; 7(3):8. https://doi.org/10.3390/jpm7030008
Chicago/Turabian StyleDa Silva, Marine S., Dominic Chartrand, Marie-Claude Vohl, Olivier Barbier, and Iwona Rudkowska. 2017. "Dairy Product Consumption Interacts with Glucokinase (GCK) Gene Polymorphisms Associated with Insulin Resistance" Journal of Personalized Medicine 7, no. 3: 8. https://doi.org/10.3390/jpm7030008
APA StyleDa Silva, M. S., Chartrand, D., Vohl, M.-C., Barbier, O., & Rudkowska, I. (2017). Dairy Product Consumption Interacts with Glucokinase (GCK) Gene Polymorphisms Associated with Insulin Resistance. Journal of Personalized Medicine, 7(3), 8. https://doi.org/10.3390/jpm7030008