Abstract
The high prevalence of obesity and of its associated diseases is a major problem worldwide. Genetic predisposition and the influence of environmental factors contribute to the development of obesity. Changes in the structure and functional activity of genes encoding adipocytokines are involved in the predisposition to weight gain and obesity. In this review, variants in genes associated with adipocyte function are examined, as are variants in genes associated with metabolic aberrations and the accompanying disorders in visceral obesity.
1. Introduction
High prevalence of obesity and of its associated diseases is a major problem worldwide [1]. Genetic predisposition and an influence of environmental factors that contribute to the development of obesity represent a set of causes of excess body weight [2,3,4,5,6]. Hereditary factors involved in the development of obesity lead to the formation of syndromic, monogenic, and polygenic types of obesity [7].
Monogenic obesity is a pathology caused by a mutation in a single gene. Monogenic obesity is extremely rare and is characterized by early onset, development in childhood, and extreme values of body weight [8,9]. Monogenic types of obesity are caused by a mutation in one of genes LEP, LEPR, POMC, PCSK1, and MC4R, which encode proteins of the leptin–melanocortin system. This system is key for the regulation of eating behavior and energy metabolism [10]. The leptin–melanocortin system is activated by leptin, which is secreted by adipocytes but exerts its action via leptin receptor, thereby leading to the activation of pro-opiomelanocortin. Under the influence of an enzyme called prohormone convertase 1, adrenocorticotropic hormone and α-melanocyte-stimulating hormone are generated from proopiomelanocortin; α-melanocyte-stimulating hormone in turn activates receptor MC4R, which launches satiety signaling [8]. Environmental factors have little effect on the development of nonsyndromic monogenic obesity but may be crucial for treatment. Physical activity, socioeconomic status, and diet type may influence obesity severity in these patients [11].
Syndromic obesity is obesity that develops in chromosomal and other genetic syndromes (e.g., Prader–Willi, fragile X, Bardet–Biedl, Cohen, and Albright) and is accompanied by a set of congenital aberrations [11].
Polygenic obesity is caused simultaneously by many genes and their interactions both with each other and with the external environment. Polygenic obesity is widespread in many populations [12]. In research on polygenic etiology of obesity by means of genome-wide association studies, 127 loci associated with obesity have been found in the human genome [7]. This number constantly changes as new studies come out regarding the polygenic etiology of obesity as a complex multifactorial disease [13]. Although polygenic obesity is the most common type of obesity and is the most responsive to clinical interventions, it is the least investigated owing to its sensitivity to environmental factors and because of its variation among ethnic groups [14].
Obesity is associated with various pathological metabolic changes in the human body: impaired glucose tolerance, insulin resistance, dyslipidemia, nonalcoholic fatty liver disease, and hypertension [15,16,17,18,19]. Visceral adiposity may have a negative effect on longevity [20]. From the point of view of the development of metabolic disorders in obesity, it is important to study visceral adipose tissue and its role as a hormonally active structure [21]. Adipocytes of visceral adipose tissue secrete a set of biomolecules: adipocytokines, which are signaling and regulatory peptides that regulate various metabolic processes [22,23,24]. In Whites with a BMI ≥ 25.0 kg/m2 and in Asians with a BMI ≥ 23.0 kg/m2, waist circumference (WC) measurement is recommended as a simple and informative method for diagnosing abdominal obesity. Values of WC ≥ 80 cm in women and WC ≥ 94 cm in men are said to correspond to abdominal obesity and increased risk of cardiovascular events [25,26]. This method does not apply to pregnant and lactating women.
Changes in the structure and function of genes encoding adipocytokines are involved in the predisposition to weight gain and obesity [7,27,28]. In this review, selected genes associated with adipocyte function are examined, as are variants in genes associated with metabolic aberrations that contribute to visceral obesity (Table 1).

Table 1.
Genes examined in this review that are associated with secretory activity of visceral adipocytes.
In this review, we analyzed the PubMed database [48] by means of the following search string: “abdominal obesity” [Title/Abstract] AND “single nucleotide polymorphism” [Title/Abstract]. We found 177 full-text articles for the period 2002–2022. Only 20 of the 177 articles (11.3%) contained information on polymorphic variants of genes associated with abdominal obesity. These 20 articles were included in the review.
At the next stage, we performed an additional search in PubMed for articles containing information about single-nucleotide polymorphisms in genes associated with adipocyte secretory activity: ADIPOQ, ADRB3, APLN, APLNR, CCL2, CCL7, FTO, GCG, GLP1R, GHRL, GIP, INS, LEP, NAMPT, PPY, PYY, RBP4, RETN, SCT, and UCP2. At this stage, we included 175 full-text articles in the review (Figure 1).
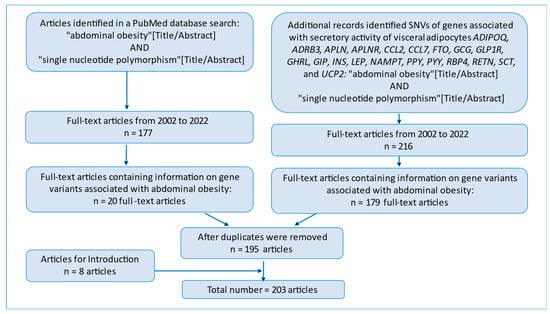
Figure 1.
A flowchart of the initial publications in PubMed.
We included an additional 8 full-text articles in Introduction. In this review, the total number of articles is 203.
Variants in the genes examined in our review participate in the development of polygenic obesity.
2. Results
2.1. ADIPOQ
The ADIPOQ gene encodes the protein adiponectin, which shares similarities with collagens X and VIII and complement factor C1q. The gene is expressed predominantly in adipose tissue, and the encoded protein circulates as various isoforms in blood plasma. Adiponectin participates in the regulation of many metabolic and hormonal processes, including carbohydrate and lipid metabolism, and has anti-inflammatory effects. Adiponectin interacts with two types of receptors: AdipoR1 and AdipoR2 [49]. Mutations in this gene can lead to adiponectin deficiency [50]. Adiponectin levels inversely correlate with both total body weight and visceral fat mass [51].
The ADIPOQ gene has many variants that result in low adiponectin levels, which correlate with obesity: the rs1391272583 variant in a Brazilian population [52], rs17366568 in a Malaysian population [53], rs266729 in a young Nigerian population [54], variants rs266729, rs16861205, rs1501299, rs3821799, and rs6773957 in a Finnish population [55], and variants rs17846866 and rs1501299 in the Indian population of Gujarat [56]. Other variants in this gene have been associated with type 2 diabetes mellitus (T2DM): rs62625753 and rs17366743 in a French white population [57], rs185847354 in a Japanese population [58], and rs2241766, rs2082940, and rs266729 in a Finnish population [55]. The ADIPOQ rs266729-G allele affects body fatness in response to dietary monounsaturated fatty acids [59].
2.2. ADRB3
This gene encodes a receptor that belongs to the β-adrenergic receptor family. The activity of the gene is regulated by catecholamines. ADRB3 is expressed mostly in adipose tissue and is responsible for the modulation of lipolysis and thermogenesis [60]. Lipolysis-produced free fatty acids via upregulation of ADRB3 accelerate transcription of the UCP1 gene and raise the activity of the UCP1 protein [61].
Several variants of the ADRB3 gene are associated with the development of obesity and T2DM. One of the first studied variants in this gene, rs4994, has been found to correlate with severe cases of obesity and insulin resistance in a Japanese population [62]. Subsequently, the rs4994 variant has been investigated in many populations and has shown an association with the risk of childhood and adolescent obesity and being overweight [63]. The correlation of rs4994 (ADRB3 gene) with obesity may be mediated by the impact of this polymorphism on adipokines. Carriers of its C allele have anomalous blood levels of adipokines and lipids [64]. Variants rs72655364 and rs72655365 of this gene correlate with T2DM in a Chinese population [65].
2.3. APLN and APLNR
The APLN gene codes for a preproprotein that is subsequently cleaved and activated in the endoplasmic reticulum. The protein apelin is secreted as a peptide hormone that binds to apelin receptors located in various organs. Apelin plays an important part in the regulation of many biological functions, including insulin secretion [66].
The rs2281068 variant (APLN gene) is associated with T2DM in the Chinese Han population [67], whereas the rs3115757 variant correlates with obesity among women in a Chinese population [68]. The receptor (encoded by the APLNR gene) to which apelin binds is expressed in the spleen, brain, placenta, and adipose tissues [69].
Studies on APLNR gene variants have not detected any associations with diabetes mellitus or obesity [70]. Some variants in the APLNR gene correlate with the risk of hypertension [71].
2.4. CCL2 and CCL7
Genes CCL2 and CCL7 are members of the CC subfamily of the chemokine superfamily, which plays a crucial role in immunomodulatory and inflammatory processes. The two genes are located side by side on chromosome 17 [72].
The serum concentration of CCL2 correlates with insulin resistance and a high body–mass index (BMI) [73]. In a Mexican population, the G allele of variant rs1024611 is associated with lower serum insulin levels, a lower BMI, lower adipose-tissue volume, and higher adiponectin levels than is the A allele [74]. The G allele of variant rs1024611 is less common among women with gestational diabetes mellitus, whereas its GG genotype is associated with a lower BMI among women with gestational diabetes mellitus [75].
CCL7 is overexpressed in the adipose tissue of obese people [76]. This upregulation is induced by elevated expression of interferon regulatory factor 5 in adipose tissue [77].
2.5. FTO
The FTO gene codes for a protein that plays an important part in the development of obesity and T2DM [78]. The FTO gene was one of the first identified loci correlating with obesity in genome-wide association studies [7]. This gene is widely expressed within the human body, with the highest expression in the brain. Several reports from different populations indicate an association of some variants of FTO with obesity in children and adults [79]. Two of the earliest genome-wide association studies in this field showed a correlation of FTO variants with early onset of obesity in a German population [80] and with T2DM in a Finnish population [81]. The rs1421085 variant of FTO is associated with impaired differentiation of adipocytes. The FTO SNP rs1421085 disrupts the binding site of repressor ARID5B [33]. The FTO SNP rs1558902 correlates with the BMI [82,83]. FTO variants implicated in obesity have been found in diverse populations: rs1421085, rs8050136, and rs9939609 in an Indian population [84], rs9939609 in a Russian population [85], rs9939609, rs1121980, and rs1558902 in a Japanese population [86,87], and rs9939609 in a Saudi Arabian population [88]. Rs9939609 is associated of with the risk of central obesity in Chinese children [89]. Rs9939609 has shown a significant correlation with male visceral obesity in an Indonesian population [90]. Increased abdominal fatness is associated with the AA genotype of a common SNP of FTO (rs9939609, T/A) when measured as waist circumference and intra-abdominal adipose tissue [91]. On the other hand, the rs9939609 polymorphism of this gene correlates with fat accumulation in the whole body without being associated with abdominal fat accumulation in Turkish adults [92]. Rs9939609 is in the same linkage region as rs8050136 (r2 = 1) [93]. A rare variant—rs140101381 (R80W)—is associated with early onset of obesity [27]. The T allele at the rs3751812 locus has been implicated in increased waist circumference in Asian-Indian populations [94]. Rs7185735 is associated with an increase in subcutaneous adipose tissue and a decrease in the visceral adipose tissue/subcutaneous adipose tissue ratio and correlates with the childhood BMI [95,96]. FTO rs9936385 is responsible for significant differences in the BMI between adults ≤50 years of age and adults aged >50 years [97].
2.6. GCG and GLP1R
The GCG gene encodes glucagon, a hormone that regulates blood glucose levels. GCG is expressed mainly in pancreatic α-cells in the islets of Langerhans and in the small intestine. As a consequence of a post-translational modification in α-cells of the pancreas, glucagon is generated, and in the small intestine, a post-translational modification results in the production of glucagon-like peptides 1 and 2, oxyntomodulin, and glicentin. Glicentin is a 69 amino acid peptide, which contains the entire sequences of oxyntomodulin (and hence glucagon) and glicentin-related pancreatic peptide [98].
GCG variants related to T2DM and obesity have been identified. In a Danish population, carriers of the homozygous GG genotype of rs4664447 have lower plasma glucose-stimulated insulin levels [99]. The rs12104705 variant correlates with general and abdominal obesity [34].
The GLP1R gene encodes a transmembrane receptor for glucagon-like peptide 1 [100]. The receptor is internalized by binding to GLP1 or its analogs, and activation of this receptor results in insulin secretion [35].
In 2004, an association of rs367543060 of the GLP1R gene with both impaired insulin secretion and aberrant insulin sensitivity was identified for the first time in patients with T2DM [101]. Variants rs2268641 and rs6923761 in this gene correlate with a high BMI [102,103]. Carriers of the AA genotype of rs6923761 have a higher risk of obesity and higher glucose levels [104]. Variant rs10305492 in a European population [105] and rs3765467 and rs10305492 in a Chinese population [106] have been implicated in metabolic syndrome and a higher risk of T2DM. Interaction between various factors, including the single-nucleotide polymorphisms (SNPs) that alter signaling, transport, and receptor activity, is key to the design of next-generation personalized agonists of GLP1R [107].
2.7. GHRL
The GHRL gene (ghrelin and obestatin prepropeptide) encodes a preproprotein that is later cleaved thereby yielding ghrelin and obestatin, which are expressed and secreted primarily in the stomach and much less often in the small intestine. The peptide ghrelin binds to its receptor (GHSR) in the hypothalamus and drives growth hormone secretion [108]. Ghrelin modulates many metabolic pathways and performs a crucial function in the reward system via the mesolimbic pathway [109]. Higher prevalence of variants rs34911341 and rs696217 has been found among obese people than in the general population [110,111].
Subsequent research has uncovered an association of various other GHRL variants with obesity: rs4684677 in a general-population cohort of European origin [112]; rs35682 and rs35683 in a white American population [113], and rs696217 in a Japanese population [114]; in addition, rs35681 correlates with the development of obesity in polycystic ovary syndrome [115].
Some variants in the GHRL gene are associated with disorders of carbohydrate metabolism. Rs27647 correlates with lower insulin levels in the oral glucose tolerance test (at 2 h after the glucose challenge) [112]. One of ghrelin’s effects related to weight changes is an alteration of eating behavior; for example, variants rs696217 and rs2075356 have been implicated in bulimia nervosa [116].
2.8. GIP
The GIP gene (gastric inhibitory polypeptide) is located on chromosome 17 and encodes an incretin hormone that regulates insulin secretion and ensures blood glucose homeostasis [117].
Three SNPs (rs3895874, rs3848460, and rs937301) have been analyzed in the 5′ region of the human GIP gene. Functional studies have revealed that in the promoter region of GIP, rare alleles of these three SNPs [haplotype GIP(−1920A)] correspond to significantly lower transcriptional activity than do the common alleles of these SNPs [haplotype GIP(−1920G)] [118]. Rs9904288, which is located at the 3′ end of GIP, is significantly associated with visceral fat area [119].
2.9. INS
The INS gene codes for the hormone insulin, which is responsible for the modulation of carbohydrate and lipid metabolism. The gene is located on chromosome 11 [120]. Insulin synthesis in pancreatic β-cells starts with preproinsulin [121]. During transport to the endoplasmic reticulum, the signal peptide located at the N terminus is cleaved off, then the remaining polypeptide is folded with the formation of disulfide bridges between the α and β chains, after which proinsulin is transported to the Golgi apparatus, where it is packaged into vesicles. Proinsulin is cleaved in vesicles and loses its C-peptide, which is subsequently secreted along with insulin molecules in equal amounts [122,123].
Some mutations in the insulin gene can lead to specific subtypes of diabetes, such as mutant INS gene–induced diabetes of youth (MIDY), maturity onset diabetes of the young (MODY), or neonatal diabetes [124,125]. Variants A24D, F48C, and R89C (INS gene), which are implicated in the development of neonatal diabetes, result in inefficient processing of proinsulin [126]. Rare variants R6H and R46Q have been described in familial cases of maturity onset diabetes of the young [127]. The A24D variant is associated with inefficient cleavage of preproinsulin [124], whereas the V92L variant weakens the affinity of insulin for insulin receptor [125], and both lead to mutant INS gene–induced diabetes of youth.
2.10. LEP
The leptin gene encodes a protein that is expressed by white adipocytes and secreted into the blood. Leptin plays an important role in the regulation of energy homeostasis and body weight control [128]. Leptin circulating in the blood binds to leptin receptor and has a central effect and peripheral effect. In the brain, leptin activates appetite-regulating signaling pathways that cause a decrease in food intake. The influence of metabolic stressors on leptin secretion has become a major focus of research on feeding behaviors [129]. The identification of central actions of leptin during signaling through its specific LepRls in neurons of neuroendocrine and hippocampal circuits has revealed complex integrated central control of body energy (expenditure vs. storage) and sophisticated self-defense of the brain, thus shedding light on how leptin modulates satiety and compulsion [130]. Reductions in energy reserves and in leptin production appear to cause a compensatory change in the reward system and the secretion of orexigenic and anorexigenic neuropeptides [131,132].
Leptin also regulates bone mass and the secretion of hypothalamic–pituitary–adrenal hormones. At the periphery, leptin accelerates basal metabolism and modulates the function of pancreatic β-cells and insulin secretion. In intestines, leptin activates protein kinase C and reduces glucose absorption [133]. Leptin levels also correlate with the waist-to-hip ratio and BMI. Leptin levels are higher in individuals with obesity compared to those without obesity [134].
Mutations in this gene may be the cause of severe obesity [133]. Mutations in the LEP gene have also been associated with the development of T2DM [128].
Mutations in the leptin gene leading to leptin deficiency are some of the causes of monogenic obesity with such symptoms as impaired satiety, hyperphagia, early onset of obesity, and many metabolic and immunological disorders [135]. These autosomal recessive pathogenic mutations are well studied and differ from the LEP variants that do not lead to monogenic obesity but may be a risk factor of obesity and other metabolic disorders.
In a study on common variants in the leptin gene, researchers showed an association of the AA genotype of the rs7799039 variant with the development of obesity in the population of South India [136]. The A allele of rs2167270 significantly correlates with an elevated risk of prediabetes in Jordan and in the population of South India [137,138,139]. Rs6966536 (allele G) of the LEP gene has been implicated in the development of obesity in a South African population [140].
Among African Americans, two SNPs of the LEP gene (rs4731427 and rs17151919) have been associated with weight, BMI, and WC. Among whites, rs2167270 and rs17151913 (in this gene) correlate with weight, whereas in women, rs28954369 is associated with weight, BMI, and WC [141]. In a study by Yaghootkar et al., missense variant Val94Met (rs17151919) was found to occur only among people of African descent, and its association with lower concentrations of leptin was specific to this origin [142].
Despite abundant research on leptin, the mechanisms of action of variants on the expression and stability of leptin or on its interaction with its receptor are still being investigated. A study by Hagglund et al. offers an explanation that includes the following mechanisms: (1) weaker affinity of leptin for its receptor, (2) blockade of leptin receptor, (3) destabilization of leptin, and (4) improper coagulation/aggregation of leptin [143].
2.11. NAMPT
This gene encodes an enzyme that catalyzes the condensation of nicotinamide with 5-phosphoribosyl-1-pyrophosphate. The gene is located on chromosome 7 [144]. The enzyme is secreted into extracellular space, and its secreted form is known as visfatin. The latter acts as a cytokine and adipokine [145].
Many NAMPT variants have been identified that correlate with changes in its metabolic function and adipokine activity. The T allele of the −948G/T variant is associated with lower fasting insulin levels [146] and higher levels of high-density lipoprotein cholesterol [147]. NAMPT variants rs10487818 and rs3801266 correlate with changes in body weight [148,149]. The rs3801267 variant has shown an association with a lower BMI in a Chinese population [150]. Rs34861192 and rs13237989 (NAMPT) affect insulin levels and the glycemic index [151]. The aforementioned variants cause either higher or lower serum levels of visfatin, which in turn leads to alterations of insulin and blood glucose levels [152].
2.12. PYY and PPY
Genes PPY and PYY code for two peptides from the neuropeptide family: pancreatic polypeptide (PP) and peptide YY (PYY). Both genes are located on chromosome 17 [153,154].
Neuropeptides typically affect the gut–brain axis by regulating appetite and energy homeostasis. The peptides of this family act through five receptors called “Y-receptors”: Y1, Y2, Y4, Y5, and Y6. These receptors differ in their tissue distribution, function, and selectivity of binding to various neuropeptides. Peptide PYY acts on the arcuate nucleus of the hypothalamus, where the Y-receptors are located that have strong affinity for PYY [155]. Gene variants in PYY are associated with changes in body weight and with obesity. Variants rs11684664, rs162430, and rs1058046 have been implicated in the development of obesity in various reports [156,157,158].
Peptide PPY is synthesized in the pancreas as a 94-amino-acid polypeptide; then, it is cleaved into 2 peptides, with only the 36-amino-acid PPY fragment being active [153]. The secreted peptide performs a function similar to that of PYY but has stronger affinity for receptor Y4. The PPY peptide is secreted by very rare γ-cells in pancreatic islets [159]. According to one research article, the GG genotype of variant rs231472 in the PPY gene correlates with the risk of obesity in children in Korea [160].
2.13. RBP4
The RBP4 gene is located on chromosome 10 and codes for a protein that binds retinol and helps to transport it from the skin to peripheral tissues. The protein is expressed mostly in the liver (and at lower levels in adipose tissues). Retinol bound to RBP4 is then imported into the cell by STRA6, which serves as a receptor of the RBP4–retinol complex [161].
RBP4 was first described as an adipokine in 2005 [162]. Variants rs34571439 and rs3758539 are associated with decreased insulin secretion in whites [163]. The rs34571439 variant (RBP4 gene) has been implicated in hypertriglyceridemia [164] and childhood obesity [165]. Variant rs7091052 is associated with the risk of gestational diabetes mellitus [166].
2.14. RETN
This gene encodes a protein called resistin. The resistin gene is located on chromosome 19 [167]. Resistin has a pleiotropic role in inflammation, in the biology of stress, and in the pathogenesis of obesity [168].
Variants in the RETN gene correlate with signs of metabolic syndrome. Investigation has revealed an association of variant rs1862513 with the risk of obesity and T2DM [169,170]. Other variants (rs3745367, rs1423096, and rs10401670) of this gene have been associated with some parameters of metabolic syndrome in various populations [171,172,173].
Differences in serum resistin levels may be related to alterations of methylation patterns in the RETN promoter region [174]. Variants in coding regions of RETN have a greater impact on this protein’s stability [175].
2.15. SCT
The SCT gene is located on chromosome 11 and codes for hormone secretin. Secretin stimulates bile and bicarbonate secretion in the duodenum, pancreas, and bile ducts [176]. Secretin expression activates brown adipose tissue, reduces central responses to appetizing food, and delays motivation for eating again after a meal [177,178].
2.16. UCP2
The UCP2 gene is in the same cluster as the UCP3 gene and is situated on chromosome 11 [179]. It is thought to take part in unregulated thermogenesis, obesity, and diabetes mellitus [180]. Variant rs659366 is associated with the risk of obesity [85,181,182,183,184], whereas variant rs660339 correlates with the risk of obesity in a specific population: Indonesians [185,186,187].
Modern molecular genetic and biochemical technologies are key to solving such fundamental problems as the formation of overweightness and obesity, thereby making it possible to identify genes and their products participating in the pathogenesis of obesity, to investigate the molecular mechanisms of relevant pathological processes, and to determine molecular biological heterogeneity of pathological phenotypes [7,188].
In addition to the determination of the BMI and WC, a Body Shape Index (ABSI) is used to diagnose obesity. ABSI takes into account body height, weight, and WC. ABSI has independent genetic associations [189,190]. ABSI can more precisely quantify abdominal obesity in metabolic syndrome [191]. Other indicators may also help to diagnose abdominal obesity, but all of them maintain a significant correlation with the BMI [192]. The association of polymorphisms of genes with ABSI may be an interesting research topic for clarifying functional significance of SNPs.
An effective method of early diagnosis and prevention of obesity is the identification of genetic markers of predisposition to this pathology and research on their characteristics in various populations [193]. Most results presented in the review have been obtained in case-control studies. This method complements data from genome-wide association studies [194,195,196]. Table 2 shows the main variants that are associated with metabolic disorders and located in the genes presented in this review.
Table 2.
Single-nucleotide variants associated with metabolic disorders.
Based on the analysis of functional significance of the variants, a scale of an individual’s genetic risk of obesity may be constructed. The creation of such a scale is an important task for modern endocrinology. The complexity of this task is due to population specificity of (i) prevalence rates of obesity, (ii) characteristics of climatic and social living conditions, and (iii) genetic factors. When a scale of individual genetic risk is created, it is important to take into account the ethnicity of the patients included in the research. For example, according to many studies, the same scale of individual genetic risk detects an association with cardiovascular events in one population and does not in another [197,198]. At present, there are not many published studies on the development of an ethnospecific scale of individual genetic risk of obesity. Most of these studies have been conducted on white cohorts, and the results may not be applicable to other populations. In research projects examining the association between a scale of individual genetic risk and diseases, Cox’s proportional hazards model and logistic regression analysis are used. Correlations between a genetic risk scale and anthropometric and biochemical parameters are assessed via linear regression. ROC (receiver operating characteristic), the area under the ROC curve (i.e., C-statistic), NRI, and integrated discrimination improvement are metrics of the quality of a scale of individual genetic risk as compared to traditional risk factors [199].
The risk of obesity, as assessed by the genetic risk scale, can be modified, for example, by a therapy aimed at weight loss and alleviation of comorbidities. It is now recognized that our everyday exercise and nutrition choices have long-term consequences for our brain function [200,201,202,203]. Early detection of a genetic risk may help to improve quality of life and life expectancy and to reduce economic costs of treatment.
3. Conclusions
It is necessary to study each population, both in terms of the nature of the variation of genes predisposing to diseases associated with excess body weight and in terms of specific features of their phenotypic manifestation. High-throughput sequencing technologies allow investigators to obtain new information about the variation of the structure of genes in groups from the general population and in clinical groups of overweight and obese individuals.
Author Contributions
A.B.: writing—original draft; D.I.: writing—original draft; E.S.: conceptualization, writing—original draft, funding acquisition, and participation in the discussion. All authors have read and agreed to the published version of the manuscript.
Funding
The work was carried out within the framework of Russian Science Foundation grant No. 21-15-00022.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We would like to thank Nikolay Shevchuk by the English editing. We thank the reviewers for their useful comments that helped to significantly improve our paper.
Conflicts of Interest
The authors declare that they have no conflict of interest.
References
- Caballero, B. Humans against Obesity: Who Will Win? Adv. Nutr. 2019, 10 (Suppl. S1), S4–S9. [Google Scholar] [CrossRef]
- Lin, X.; Li, H. Obesity: Epidemiology, Pathophysiology, and Therapeutics. Front. Endocrinol. 2021, 12, 706978. [Google Scholar] [CrossRef]
- Ling, C.; Rönn, T. Epigenetics in Human Obesity and Type 2 Diabetes. Cell Metab. 2019, 9, 1028–1044. [Google Scholar] [CrossRef] [PubMed]
- Herrera, B.M.; Lindgren, C.M. The genetics of obesity. Curr. Diab. Rep. 2010, 10, 498–505. [Google Scholar] [CrossRef]
- Vettori, A.; Pompucci, G.; Paolini, B.; Del Ciondolo, I.; Bressan, S.; Dundar, M.; Kenanoğlu, S.; Unfer, V.; Bertelli, M.; Geneob Project. Genetic background, nutrition and obesity: A review. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 1751–1761. [Google Scholar] [CrossRef]
- Hurtado, A.M.D.; Acosta, A. Precision Medicine and Obesity. Gastroenterol. Clin. N. Am. 2021, 50, 127–139. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.K.; Kumar, P.; Mahalingam, K. Molecular Genetics of Human Obesity: A Comprehensive Review. C R Biol. 2017, 340, 87–108. [Google Scholar] [CrossRef] [PubMed]
- Hebebrand, J.; Hinney, A.; Knoll, N.; Volckmar, A.L.; Scherag, A. Molecular genetic aspects of weight regulation. Dtsch. Arztebl. Int. 2013, 110, 338–344. [Google Scholar] [CrossRef] [PubMed]
- Muñoz, C.; Garcia-Vargas, G.G.; Morales, R.P. Monogenic, Polygenic and Multifactorial Obesity in Children: Genetic and Environmental Factor. Austin J. Nutr. Metab. 2017, 4, 1052. [Google Scholar]
- Littleton, S.H.; Berkowitz, R.I.; Grant, S.F.A. Genetic Determinants of Childhood Obesity. Mol. Diagn. Ther. 2020, 24, 653–663. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud, R.; Kimonis, V.; Butler, M.G. Genetics of Obesity in Humans: A Clinical Review. Int. J. Mol. Sci. 2022, 23, 11005. [Google Scholar] [CrossRef] [PubMed]
- GBD 2015 Obesity Collaborators. Health effects of overweight and obesity in 195 countries over 25 years. N. Engl. J. Med. 2017, 377, 13–27. [Google Scholar] [CrossRef] [PubMed]
- Loos, R.J.F.; Yeo, G.S.H. The Genetics of Obesity: From Discovery to Biology. Nat. Rev. Genet. 2022, 23, 120–133. [Google Scholar] [CrossRef] [PubMed]
- Heianza, Y.; Qi, L. Gene-Diet Interaction and Precision Nutrition in Obesity. Int. J. Mol. Sci. 2017, 18, 787. [Google Scholar] [CrossRef]
- Litwin, M.; Kułaga, Z. Obesity, metabolic syndrome, and primary hypertension. Pediatr. Nephrol. 2021, 36, 825–837. [Google Scholar] [CrossRef]
- Ahmed, B.; Sultana, R.; Greene, M.W. Adipose tissue and insulin resistance in obese. Biomed. Pharmacother. 2021, 137, 111315. [Google Scholar] [CrossRef]
- Brunner, K.T.; Henneberg, C.J.; Wilechansky, R.M.; Long, M.T. Nonalcoholic Fatty Liver Disease and Obesity Treatment. Curr. Obes. Rep. 2019, 8, 220–228. [Google Scholar] [CrossRef]
- Milić, S.; Lulić, D.; Štimac, D. Non-alcoholic fatty liver disease and obesity: Biochemical, metabolic and clinical presentations. World J. Gastroenterol. 2014, 20, 9330–9337. [Google Scholar] [CrossRef]
- Zhang, T.; Chen, J.; Tang, X.; Luo, Q.; Xu, D.; Yu, B. Interaction between adipocytes and high-density lipoprotein:new insights into the mechanism of obesity-induced dyslipidemia and atherosclerosis. Lipids Health Dis. 2019, 18, 223. [Google Scholar] [CrossRef]
- Yan, B.; Yang, J.; Zhao, B.; Wu, Y.; Bai, L.; Ma, X. Causal Effect of Visceral Adipose Tissue Accumulation on the Human Longevity: A Mendelian Randomization Study. Front Endocrinol. 2021, 1, 722187. [Google Scholar] [CrossRef]
- Cao, H. Adipocytokines in obesity and metabolic disease. J. Endocrinol. 2014, 220, 47–59. [Google Scholar] [CrossRef] [PubMed]
- Maximus, P.S.; Al Achkar, Z.; Hamid, P.F.; Hasnain, S.S.; Peralta, C.A. Adipocytokines: Are they the Theory of Everything? Cytokine 2020, 133, 155144. [Google Scholar] [CrossRef]
- Zorena, K.; Jachimowicz-Duda, O.; Ślęzak, D.; Robakowska, M.; Mrugacz, M. Adipokines and Obesity. Potential Link to Metabolic Disorders and Chronic Complications. Int. J. Mol. Sci. 2020, 21, 3570. [Google Scholar] [CrossRef] [PubMed]
- Wen, W.; Kato, N.; Hwang, J.Y.; Guo, X.; Tabara, Y.; Li, H.; Dorajoo, R.; Yang, X.; Tsai, F.J.; Li, S. Genome-wide association studies in East Asians identify new loci for waist-hip ratio and waist circumference. Sci. Rep. 2016, 20, 17958. [Google Scholar] [CrossRef] [PubMed]
- Dedov, I.I.; Shestakova, M.V.; Melnichenko, G.A.; Mazurina, N.V.; Andreeva, E.N.; Bondarenko, I.Z.; Gusova, Z.R.; Dzgoeva, F.K.; Eliseev, M.S.; Ershova, E.V.; et al. Interdisciplinary Clinical Practice Guidelines “Management of obesity and its comorbidities”. Obes. Metab. 2021, 18, 5–99. [Google Scholar] [CrossRef]
- Yumuk, V.; Tsigos, C.; Fried, M.; Schindler, K.; Busetto, L.; Micic, D.; Toplak, H. Obesity Management Task Force of the European Association for the Study of Obesity. European Guidelines for Obesity Management in Adults. Obes. Facts 2015, 8, 402–424. [Google Scholar] [CrossRef]
- Serra-Juhé, C.; Martos-Moreno, G.; De Pieri, F.B.; Flores, R.; Chowen, J.A.; Pérez-Jurado, L.A.; Argente, J. Heterozygous rare genetic variants in non-syndromic early-onset obesity. Int. J. Obes. 2020, 44, 830–841. [Google Scholar] [CrossRef]
- Ghoshal, K.; Chatterjee, T.; Chowdhury, S.; Sengupta, S.; Bhattacharyya, M. Adiponectin Genetic Variant and Expression Coupled with Lipid Peroxidation Reveal New Signatures in Diabetic Dyslipidemia. Biochem. Genet. 2021, 59, 781–798. [Google Scholar] [CrossRef]
- Iwabu, M.; Okada-Iwabu, M.; Yamauchi, T.; Kadowaki, T. Adiponectin/AdipoR Research and Its Implications for Lifestyle-Related Diseases. Front. Cardiovasc. Med. 2019, 6, 116. [Google Scholar] [CrossRef]
- Luo, Z.; Zhang, T.; Wang, S.; He, Y.; Ye, Q.; Cao, W. The Trp64Arg Polymorphism in Β3 Adrenergic Receptor (ADRB3) Gene Is Associated with Adipokines and Plasma Lipids: A Systematic Review, Meta-Analysis, and Meta-Regression. Lipids Health Dis. 2020, 19, 99. [Google Scholar] [CrossRef]
- Li, C.; Cheng, H.; Adhikari, B.K.; Wang, S.; Yang, N.; Liu, W.; Sun, J.; Wang, Y. The Role of Apelin-APJ System in Diabetes and Obesity. Front. Endocrinol. 2022, 13, 820002. [Google Scholar] [CrossRef]
- Ignacio, R.M.; Gibbs, C.R.; Lee, E.S.; Son, D.S. Differential Chemokine Signature between Human Preadipocytes and Adipocytes. Immune Netw. 2016, 16, 189–194. [Google Scholar] [CrossRef] [PubMed]
- Claussnitzer, M.; Dankel, S.N.; Kim, K.-H.; Quon, G.; Meuleman, W.; Haugen, C.; Glunk, V.; Sousa, I.S.; Beaudry, J.L.; Puviindran, V.; et al. FTO Obesity Variant Circuitry and Adipocyte Browning in Humans. N. Engl. J. Med. 2015, 373, 895–907. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhang, M.; Wang, J.J.; Wang, C.J.; Ren, Y.C.; Wang, B.Y.; Zhang, H.Y.; Yang, X.Y.; Zhao, Y.; Han, C.Y.; et al. Association of TCF7L2 and GCG Gene Variants with Insulin Secretion, Insulin Resistance, and Obesity in New-Onset Diabetes. Biomed. Environ. Sci. 2016, 29, 814–817. [Google Scholar] [CrossRef] [PubMed]
- Mayendraraj, A.; Rosenkilde, M.M.; Gasbjerg, L.S. GLP-1 and GIP Receptor Signaling in Beta Cells—A Review of Receptor Interactions and Co-Stimulation. Peptides 2022, 151, 170749. [Google Scholar] [CrossRef] [PubMed]
- Landgren, S.; Simms, J.A.; Thelle, D.S.; Strandhagen, E.; Bartlett, S.E.; Engel, J.A.; Jerlhag, E. The Ghrelin Signalling System Is Involved in the Consumption of Sweets. PLoS ONE 2011, 6, e18170. [Google Scholar] [CrossRef]
- Nauck, M.A.; Quast, D.R.; Wefers, J.; Pfeiffer, A.F.H. The evolving story of incretins (GIP and GLP-1) in metabolic and cardiovascular disease: A pathophysiological update. Diabetes Obes. Metab. 2021, 23 (Suppl. S3), 5–29. [Google Scholar] [CrossRef] [PubMed]
- Støy, J.; Edghill, E.L.; Flanagan, S.E.; Ye, H.; Paz, V.P.; Pluzhnikov, A.; Below, J.E.; Hayes, M.G.; Cox, N.J.; Lipkind, G.M.; et al. Insulin gene mutations as a cause of permanent neonatal diabetes. Proc. Natl. Acad. Sci. USA 2007, 104, 15040–15044. [Google Scholar] [CrossRef]
- Wabitsch, M.; Pridzun, L.; Ranke, M.; von Schnurbein, J.; Moss, A.; Brandt, S.; Kohlsdorf, K.; Moepps, B.; Schaab, M.; Funcke, J.B.; et al. Measurement of immunofunctional leptin to detect and monitor patients with functional leptin deficiency. Eur. J. Endocrinol. 2017, 176, 315–322. [Google Scholar] [CrossRef]
- Curat, C.A.; Wegner, V.; Sengenes, C.; Miranville, A.; Tonus, C.; Busse, R.; Bouloumie, A. Macrophages in human visceral adipose tissue: Increased accumulation in obesity and a source of resistin and visfatin. Diabetologia 2006, 49, 744–747. [Google Scholar] [CrossRef]
- Stromsdorfer, K.L.; Yamaguchi, S.; Yoon, M.J.; Moseley, A.C.; Franczyk, M.P.; Kelly, S.C.; Qi, N.; Imai, S.I.; Yoshino, J. NAMPT-Mediated NAD+ Biosynthesis in Adipocytes Regulates Adipose Tissue Function and Multi-Organ Insulin Sensitivity in Mice. Cell Rep. 2016, 16, 1851–1860. [Google Scholar] [CrossRef] [PubMed]
- Campbell, C.D.; Lyon, H.N.; Nemesh, J.; Drake, J.A.; Tuomi, T.; Gaudet, D.; Zhu, X.; Cooper, R.S.; Ardlie, K.G.; Groop, L.C.; et al. Association studies of BMI and type 2 diabetes in the neuropeptide Y pathway: A possible role for NPY2R as a candidate gene for type 2 diabetes in men. Diabetes 2007, 56, 1460–14607. [Google Scholar] [CrossRef] [PubMed]
- Blaner, W.S. Vitamin A signaling and homeostasis in obesity, diabetes, and metabolic disorders. Pharmacol. Ther. 2019, 197, 153–178. [Google Scholar] [CrossRef] [PubMed]
- Qi, Q.; Menzaghi, C.; Smith, S.; Liang, L.; de Rekeneire, N.; Garcia, M.E.; Lohman, K.K.; Miljkovic, I.; Strotmeyer, E.S.; Cummings, S.R.; et al. Genome-wide association analysis identifies TYW3/CRYZ and NDST4 loci associated with circulating resistin levels. Hum. Mol. Genet. 2012, 21, 4774–4780. [Google Scholar] [CrossRef] [PubMed]
- Rathwa, N.; Patel, R.; Palit, S.P.; Ramachandran, A.V.; Begum, R. Genetic variants of resistin and its plasma levels: Association with obesity and dyslipidemia related to type 2 diabetes susceptibility. Genomics 2019, 111, 980–985. [Google Scholar] [CrossRef]
- Schnabl, K.; Li, Y.; Klingenspor, M. The gut hormone secretin triggers a gut-brown fat-brain axis in the control of food intake. Exp. Physiol. 2020, 105, 1206–1213. [Google Scholar] [CrossRef]
- Oliveira, M.S.; Rheinheimer, J.; Moehlecke, M.; Rodrigues, M.; Assmann, T.S.; Leitão, C.B.; Trindade, M.R.M.; Crispim, D.; de Souza, B.M. UCP2, IL18, and miR-133a-3p are dysregulated in subcutaneous adipose tissue of patients with obesity. Mol. Cell. Endocrinol. 2020, 509, 110805. [Google Scholar] [CrossRef]
- Available online: https://pubmed.ncbi.nlm.nih.gov (accessed on 14 November 2022).
- Khoramipour, K.; Chamari, K.; Hekmatikar, A.A.; Ziyaiyan, A.; Taherkhani, S.; Elguindy, N.M.; Bragazzi, N.L. Adiponectin: Structure, Physiological Functions, Role in Diseases, and Effects of Nutrition. Nutrients 2021, 13, 1180. [Google Scholar] [CrossRef]
- Pruitt, K.D.; Tatusova, T.; Brown, G.R.; Maglott, D.R. NCBI Reference Sequences (RefSeq): Current Status, New Features and Genome Annotation Policy. Nucleic. Acids Res. 2012, 40, D130–D135. [Google Scholar] [CrossRef]
- Achari, A.E.; Jain, S.K. Adiponectin, a Therapeutic Target for Obesity, Diabetes, and Endothelial Dysfunction. Int. J. Mol. Sci. 2017, 18, 1321. [Google Scholar] [CrossRef]
- Bueno, A.C.; Sun, K.; Martins, C.S.; Elias, J.; Miranda, W.; Tao, C.; Foss-Freitas, M.C.; Barbieri, M.A.; Bettiol, H.; de Castro, M.; et al. A Novel ADIPOQ Mutation (p.M40K) Impairs Assembly of High-Molecular-Weight Adiponectin and Is Associated with Early-Onset Obesity and Metabolic Syndrome. J. Clin. Endocrinol. Metab. 2014, 99, E683–E693. [Google Scholar] [CrossRef] [PubMed]
- Apalasamy, Y.D.; Rampal, S.; Salim, A.; Moy, F.M.; Bulgiba, A.; Mohamed, Z. Association of ADIPOQ Gene with Obesity and Adiponectin Levels in Malaysian Malays. Mol. Biol. Rep. 2014, 41, 2917–2921. [Google Scholar] [CrossRef] [PubMed]
- Olusegun, E.; Ogundele, O.E.; Adekoya, K.O.; Osinubi, A.A.A.; Awofala, A.A.; Oboh, B.O. Association of Adiponectin Gene (ADIPOQ) Polymorphisms with Measures of Obesity in Nigerian Young Adults. Egypt. J. Med. Hum. Genet. 2018, 19, 123–127. [Google Scholar] [CrossRef]
- Siitonen, N.; Pulkkinen, L.; Lindström, J.; Kolehmainen, M.; Eriksson, J.G.; Venojärvi, M.; Ilanne-Parikka, P.; Keinänen-Kiukaanniemi, S.; Tuomilehto, J.; Uusitupa, M. Association of ADIPOQ Gene Variants with Body Weight, Type 2 Diabetes and Serum Adiponectin Concentrations: The Finnish Diabetes Prevention Study. BMC Med. Genet. 2011, 12, 5. [Google Scholar] [CrossRef] [PubMed]
- Palit, S.P.; Patel, R.; Jadeja, S.D.; Rathwa, N.; Mahajan, A.; Ramachandran, A.V.; Dhar, M.K.; Sharma, S.; Begum, R. A Genetic Analysis Identifies a Haplotype at Adiponectin Locus: Association with Obesity and Type 2 Diabetes. Sci. Rep. 2020, 10, 2904. [Google Scholar] [CrossRef]
- Vasseur, F.; Helbecque, N.; Dina, C.; Lobbens, S.; Delannoy, V.; Gaget, S.; Boutin, P.; Vaxillaire, M.; Leprêtre, F.; Dupont, S.; et al. Single-Nucleotide Polymorphism Haplotypes in the Both Proximal Promoter and Exon 3 of the APM1 Gene Modulate Adipocyte-Secreted Adiponectin Hormone Levels and Contribute to the Genetic Risk for Type 2 Diabetes in French Caucasians. Hum. Mol. Genet. 2002, 11, 2607–2614. [Google Scholar] [CrossRef] [PubMed]
- Kondo, H.; Shimomura, L.; Matsukawa, Y.; Kumada, M.; Takahashi, M.; Matsuda, M.; Ouchi, N.; Kihara, S.; Kawamoto, T.; Sumitsuji, S.; et al. Association of Adiponectin Mutation With Type 2 Diabetes A Candidate Gene for the Insulin Resistance Syndrome. Diabetes 2002, 51, 2325–2328. [Google Scholar] [CrossRef]
- Hammad, S.S.; Eck, P.; Sihag, J.; Chen, X.; Connelly, P.W.; Lamarche, B.; Couture, P.; Guay, V.; Maltais-Giguère, J.; West, S.G.; et al. Common Variants in Lipid Metabolism-Related1 Genes Associate with Fat Mass Changes in Response to Dietary Monounsaturated Fatty Acids in Adults with Abdominal Obesity. J. Nutr. 2019, 149, 1749–1756. [Google Scholar] [CrossRef]
- ADRB3 Adrenoceptor Beta 3 [Homo sapiens (Human)]. Available online: https://www.ncbi.nlm.nih.gov/gene/155 (accessed on 14 November 2022).
- Takenaka, A.; Nakamura, S.; Mitsunaga, F.; Inoue-Murayama, M.; Udono, T.; Suryobroto, B. Human-specific SNP in obesity genes, adrenergic receptor beta2 (ADRB2), Beta3 (ADRB3), and PPAR γ2 (PPARG), during primate evolution. PLoS ONE 2012, 7, e43461. [Google Scholar] [CrossRef]
- Kadowaki, H.; Yasuda, K.; Iwamoto, K.; Otabe, S.; Shimokawa, K.; Silver, K.; Walston, J.; Yoshinaga, H.; Kosaka, K.; Yamada, N.; et al. A Mutation in the Β3-Adrenergic Receptor Gene Is Associated with Obesity and Hyperinsulinemia in Japanese Subjects. Biochem. Biophys. Res. Commun 1995, 215, 555–560. [Google Scholar] [CrossRef]
- Xie, C.; Hua, W.; Zhao, Y.; Rui, J.; Feng, J.; Chen, Y.; Liu, Y.; Liu, J.; Yang, X.; Xu, X. The ADRB3 Rs4994 Polymorphism Increases Risk of Childhood and Adolescent Overweight/Obesity for East Asia’s Population: An Evidence-Based Meta-Analysis. Adipocyte 2020, 9, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Ryukm, J.A.; Zhang, X.; Ko, B.S.; Daily, J.W.; Park, S. Association of β3-adrenergic receptor rs4994 polymorphisms with the risk of type 2 diabetes: A systematic review and meta-analysis. Diabetes Res. Clin. Pract. 2017, 129, 86–96. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; Yang, T.L.; Tang, B.S.; Chen, X.; Huang, X.; Luo, X.H.; Zhu, Y.S.; Chen, X.P.; Hu, P.C.; Chen, J.; et al. Two Novel Functional Single Nucleotide Polymorphisms of ADRB3 Are Associated with Type 2 Diabetes in the Chinese Population. J. Clin. Endocrinol. Metab. 2013, 98, E1272–E1277. [Google Scholar] [CrossRef] [PubMed]
- Zhong, J.C.; Zhang, Z.Z.; Wang, W.; McKinnie, S.M.K.; Vederas, J.C.; Oudit, G.Y. Targeting the Apelin Pathway as a Novel Therapeutic Approach for Cardiovascular Diseases. Biochim. Biophys. Acta Mol. Basis. Dis. 2017, 1863, 1942–1950. [Google Scholar] [CrossRef]
- Zheng, H.; Fan, X.; Li, X.; Zhang, Y.; Fan, Y.; Zhang, N.; Song, Y.; Ren, F.; Shen, C.; Shen, J.; et al. The Association between Single Nucleotide Polymorphisms of the Apelin Gene and Diabetes Mellitus in a Chinese Population. J. Pediatr. Endocrinol. Metab. 2016, 29, 1397–1402. [Google Scholar] [CrossRef]
- Liao, Y.C.; Chou, W.W.; Li, Y.N.; Chuang, S.C.; Lin, W.Y.; Lakkakula, B.V.K.S.; Yu, M.L.; Juo, S.H.H. Apelin gene polymorphism influences apelin expression and obesity phenotypes in Chinese women. Am. J. Clin. Nutr. 2011, 94, 921–928. [Google Scholar] [CrossRef]
- Zhang, M.; Peng, F.; Lin, L.; Yu, M.; Huang, C.; Hu, D.; Guo, Q.; Xu, C.; Lin, J.; Zhang, M.; et al. Association Study of Apelin-APJ System Genetic Polymorphisms with Incident Metabolic Syndrome in a Chinese Population: A Case-Control Study. Oncotarget 2019, 10, 3807–3817. [Google Scholar] [CrossRef]
- Galon-Tilleman, H.; Yang, H.; Bednarek, M.A.; Spurlock, S.M.; Paavola, K.J.; Ko, B.; To, C.; Luo, J.; Tian, H.; Jermutus, L.; et al. Apelin-36 Modulates Blood Glucose and Body Weight Independently of Canonical APJ Receptor Signaling. J. Biol. Chem. 2017, 292, 1925–1933. [Google Scholar] [CrossRef]
- Yoshikawa, M.; Asaba, K.; Nakayama, T. The APLNR gene polymorphism rs7119375 is associated with an increased risk of development of essential hypertension in the Chinese population: A meta-analysis. Medicine 2020, 99, e22418. [Google Scholar] [CrossRef]
- Available online: https://www.ncbi.nlm.nih.gov/gene/ (accessed on 14 November 2022).
- Mikhailova, S.V.; Ivanoshchuk, D.E. Innate-Immunity Genes in Obesity. J. Pers. Med. 2021, 11, 1201. [Google Scholar] [CrossRef]
- Guzmán-Ornelas, M.O.; Petri, M.H.; Vázquez-Del Mercado, M.; Chavarría-Ávila, E.; Corona-Meraz, F.I.; Ruíz-Quezada, S.L.; Madrigal-Ruíz, P.M.; Castro-Albarrán, J.; Sandoval-García, F.; Navarro-Hernández, R.E. CCL2 Serum Levels and Adiposity Are Associated with the Polymorphic Phenotypes -2518A on CCL2 and 64ILE on CCR2 in a Mexican Population with Insulin Resistance. J. Diabetes Res. 2016, 2016, 5675739. [Google Scholar] [CrossRef] [PubMed]
- Teler, J.; Tarnowski, M.; Safranow, K.; Maciejewska, A.; Sawczuk, M.; Dziedziejko, V.; Sluczanowska-Glabowska, S.; Pawlik, A. CCL2, CCL5, IL4 and IL15 Gene Polymorphisms in Women with Gestational Diabetes Mellitus. Horm. Metab. Res. 2017, 49, 10–15. [Google Scholar] [CrossRef] [PubMed]
- Huber, J.; Kiefer, F.W.; Zeyda, M.; Ludvik, B.; Silberhumer, G.R.; Prager, G.; Zlabinger, G.J.; Stulnig, T.M. CC Chemokine and CC Chemokine Receptor Profiles in Visceral and Subcutaneous Adipose Tissue Are Altered in Human Obesity. J. Clin. Endocrinol. Metab. 2008, 93, 3215–3221. [Google Scholar] [CrossRef] [PubMed]
- Sindhu, S.; Thomas, R.; Kochumon, S.; Wilson, A.; Abu-Farha, M.; Bennakhi, A.; Al-Mulla, F.; Ahmad, R. Increased Adipose Tissue Expression of Interferon Regulatory Factor (IRF)-5 in Obesity: Association with Metabolic Inflammation. Cells 2019, 8, 1418. [Google Scholar] [CrossRef]
- Wei, J.; Liu, F.; Lu, Z.; Fei, Q.; Ai, Y.; He, P.C.; Shi, H.; Cui, X.; Su, R.; Klungland, A.; et al. Differential m 6 A, m 6 A m, and m 1 A Demethylation Mediated by FTO in the Cell Nucleus and Cytoplasm. Mol. Cell 2018, 71, 973–985.e5. [Google Scholar] [CrossRef]
- Povel, C.M.; Boer, J.M.; Onland-Moret, N.C.; Dollé, M.E.; Feskens, E.J.; van der Schouw, Y.T. Single nucleotide polymorphisms (SNPs) involved in insulin resistance, weight regulation, lipid metabolism and inflammation in relation to metabolic syndrome: An epidemiological study. Cardiovasc. Diabetol. 2012, 11, 133. [Google Scholar] [CrossRef]
- Hinney, A.; Nguyen, T.T.; Scherag, A.; Friedel, S.; Brönner, G.; Müller, T.D.; Grallert, H.; Illig, T.; Wichmann, H.E.; Rief, W.; et al. Genome Wide Association (GWA) Study for Early Onset Extreme Obesity Supports the Role of Fat Mass and Obesity Associated Gene (FTO) Variants. PLoS ONE 2007, 2, e1361. [Google Scholar] [CrossRef]
- Scott, L.J.; Mohlke, K.L.; Bonnycastle, L.L.; Willer, C.J.; Li, Y.; Duren, W.L.; Erdos, M.R.; Stringham, H.M.; Chines, P.S.; Jackson, A.U.; et al. A Genome-Wide Association Study of Type 2 Diabetes in Finns Detects Multiple Susceptibility Variants. Science 2007, 316, 1341–1345. [Google Scholar] [CrossRef]
- Larsson, S.C.; Burgess, S.; Michaëlsson, K. Genetic association between adiposity and gout: A Mendelian randomization study. Rheumatology 2018, 57, 2145–2148. [Google Scholar] [CrossRef]
- Fox, C.S.; Liu, Y.; White, C.C.; Feitosa, M.; Smith, A.V.; Heard-Costa, N.; Lohman, K.; GIANT Consortium; MAGIC Consortium; GLGC Consortium; et al. Genome-wide association for abdominal subcutaneous and visceral adipose reveals a novel locus for visceral fat in women. PLoS Genet. 2012, 8, e1002695. [Google Scholar] [CrossRef]
- Chauhan, G.; Tabassum, R.; Mahajan, A.; Dwivedi, O.P.; Mahendran, Y.; Kaur, I.; Nigam, S.; Dubey, H.; Varma, B.; Madhu, S.V.; et al. Common Variants of FTO and the Risk of Obesity and Type 2 Diabetes in Indians. J. Hum. Genet. 2011, 56, 720–726. [Google Scholar] [CrossRef]
- Baturin, A.K.; Sorokina, E.Y.; Pogozheva, A.V.; Keshabyants, E.E.; Kobelkova, I.v.; Kambarov, A.O.; Elizarova, E.v.; Tutelyan, V.A. The Association of Rs993609 Polymorphisms of Gene FTO and Rs659366 Polymorphisms of Gene UCP2 with Obesity among Arctic Russian Population. Vopr. Pitan. 2017, 86, 32–39. [Google Scholar] [CrossRef]
- Matsuo, T.; Nakata, Y.; Hotta, K.; Tanaka, K. The FTO genotype as a useful predictor of body weight maintenance: Initial data from a 5-year follow-up study. Metabolism 2014, 63, 912–917. [Google Scholar] [CrossRef]
- Kamura, Y.; Iwata, M.; Maeda, S.; Shinmura, S.; Koshimizu, Y.; Honoki, H.; Fukuda, K.; Ishiki, M.; Usui, I.; Fukushima, Y.; et al. FTO Gene Polymorphism Is Associated with Type 2 Diabetes through Its Effect on Increasing the Maximum BMI in Japanese Men. PLoS ONE 2016, 11, e0165523. [Google Scholar] [CrossRef] [PubMed]
- Cyrus, C.; Ismail, M.H.; Chathoth, S.; Vatte, C.; Hasen, M.; al Ali, A. Analysis of the Impact of Common Polymorphisms of the FTO and MC4R Genes with the Risk of Severe Obesity in Saudi Arabian Population. Genet. Test Mol. Biomark. 2018, 22, 170–177. [Google Scholar] [CrossRef] [PubMed]
- Xi, B.; Cheng, H.; Shen, Y.; Chandak, G.R.; Zhao, X.; Hou, D.; Wu, L.; Wang, X.; Mi, J. Study of 11 BMI-associated loci identified in GWAS for associations with central obesity in the Chinese children. PLoS ONE 2013, 8, e56472. [Google Scholar] [CrossRef] [PubMed]
- Salim, S.; Kartawidjajaputra, F.; Suwanto, A. Association of FTO rs9939609 and CD36 rs1761667 with Visceral Obesity. J. Nutr. Sci. Vitam. 2020, 66, S329–S335. [Google Scholar] [CrossRef] [PubMed]
- Kring, S.I.; Holst, C.; Zimmermann, E.; Jess, T.; Berentzen, T.; Toubro, S.; Hansen, T.; Astrup, A.; Pedersen, O.; Sørensen, T.I. FTO gene associated fatness in relation to body fat distribution and metabolic traits throughout a broad range of fatness. PLoS ONE 2008, 3, e2958. [Google Scholar] [CrossRef] [PubMed]
- Ağagündüz, D.; Gezmen-Karadağ, M. Association of FTO common variant (rs9939609) with body fat in Turkish individuals. Lipids Health Dis. 2019, 18, 212. [Google Scholar] [CrossRef]
- Kong, X.; Xing, X.; Zhang, X.; Hong, J.; Yang, W. Sexual Dimorphism of a Genetic Risk Score for Obesity and Related Traits among Chinese Patients with Type 2 Diabetes. Obes. Facts 2019, 12, 328–343. [Google Scholar] [CrossRef]
- Moore, S.C.; Gunter, M.J.; Daniel, C.R.; Reddy, K.S.; George, P.S.; Yurgalevitch, S.; Devasenapathy, N.; Ramakrishnan, L.; Chatterjee, N.; Chanock, S.J.; et al. Common genetic variants and central adiposity among Asian-Indians. Obesity 2012, 20, 1902–1908. [Google Scholar] [CrossRef] [PubMed]
- Monnereau, C.; Santos, S.; van der Lugt, A.; Jaddoe, V.W.V.; Felix, J.F. Associations of adult genetic risk scores for adiposity with childhood abdominal, liver and pericardial fat assessed by magnetic resonance imaging. Int. J. Obes. 2018, 42, 897–904. [Google Scholar] [CrossRef] [PubMed]
- Monnereau, C.; Vogelezang, S.; Kruithof, C.J.; Jaddoe, V.W.; Felix, J.F. Associations of genetic risk scores based on adult adiposity pathways with childhood growth and adiposity measures. BMC Genet. 2016, 17, 120. [Google Scholar] [CrossRef] [PubMed]
- Winkler, T.W.; Justice, A.E.; Graff, M.; Barata, L.; Feitosa, M.F.; Chu, S.; Czajkowski, J.; Esko, T.; Fall, T.; Kilpeläinen, T.O.; et al. The Influence of Age and Sex on Genetic Associations with Adult Body Size and Shape: A Large-Scale Genome-Wide Interaction Study. PLoS Genet. 2015, 11, e1005378. [Google Scholar] [CrossRef] [PubMed]
- GCG Glucagon [Homo sapiens (Human)]. Available online: https://www.ncbi.nlm.nih.gov/gene/2641 (accessed on 14 November 2022).
- Torekov, S.S.; Ma, L.; Grarup, N.; Hartmann, B.; Hainerová, I.A.; Kielgast, U.; Kissow, H.; Rosenkilde, M.; Lebl, J.; Witte, D.R.; et al. Homozygous Carriers of the G Allele of Rs4664447 of the Glucagon Gene (GCG) Are Characterised by Decreased Fasting and Stimulated Levels of Insulin, Glucagon and Glucagon-like Peptide (GLP)-1. Diabetologia 2011, 54, 2820–2831. [Google Scholar] [CrossRef] [PubMed]
- GLP1R Glucagon Like Peptide 1 Receptor [Homo sapiens (Human)]. Available online: https://www.ncbi.nlm.nih.gov/gene/2740 (accessed on 14 November 2022).
- Tokuyama, Y.; Matsui, K.; Egashira, T.; Nozaki, O.; Ishizuka, T.; Kanatsuka, A. Five Missense Mutations in Glucagon-like Peptide 1 Receptor Gene in Japanese Population. Diabetes Res. Clin. Pract. 2004, 66, 63–69. [Google Scholar] [CrossRef]
- Li, P.; Tiwari, H.K.; Lin, W.Y.; Allison, D.B.; Chung, W.K.; Leibel, R.L.; Yi, N.; Liu, N. Genetic Association Analysis of 30 Genes Related to Obesity in a European American Population. Int. J. Obes. 2014, 38, 724–729. [Google Scholar] [CrossRef]
- de Luis, D.A.; Aller, R.; de la Fuente, B.; Primo, D.; Conde, R.; Izaola, O.; Sagrado, M.G. Relation of the Rs6923761 Gene Variant in Glucagon-like Peptide 1 Receptor with Weight, Cardiovascular Risk Factor, and Serum Adipokine Levels in Obese Female Subjects. J. Clin. Lab. Anal. 2015, 29, 100–105. [Google Scholar] [CrossRef]
- Michałowska, J.; Miller-Kasprzak, E.; Seraszek-Jaros, A.; Mostowska, A.; Bogdański, P. Association of GLP1R Variants Rs2268641 and Rs6923761 with Obesity and Other Metabolic Parameters in a Polish Cohort. Front. Endocrinol. 2022, 13, 1000185. [Google Scholar] [CrossRef]
- Wessel, J.; Chu, A.Y.; Willems, S.M.; Wang, S.; Yaghootkar, H.; Brody, J.A.; Dauriz, M.; Hivert, M.F.; Raghavan, S.; Lipovich, L.; et al. Low-Frequency and Rare Exome Chip Variants Associate with Fasting Glucose and Type 2 Diabetes Susceptibility. Nat. Commun. 2015, 6, 5897. [Google Scholar] [CrossRef]
- Li, W.; Li, P.; Li, R.; Yu, Z.; Sun, X.; Ji, G.; Yang, X.; Zhu, L.; Zhu, S. GLP1R Single-Nucleotide Polymorphisms Rs3765467 and Rs10305492 Affect β Cell Insulin Secretory Capacity and Apoptosis through GLP-1. DNA Cell Biol. 2020, 39, 1700–1710. [Google Scholar] [CrossRef] [PubMed]
- El Eid, L.; Reynolds, C.A.; Tomas, A.; Jones, B. Biased Agonism and Polymorphic Variation at the GLP-1 Receptor: Implications for the Development of Personalised Therapeutics. Pharmacol. Res. 2022, 184, 106411. [Google Scholar] [CrossRef] [PubMed]
- GHRL Ghrelin and Obestatin Prepropeptide [Homo sapiens (Human)]. Available online: https://www.ncbi.nlm.nih.gov/gene/51738 (accessed on 14 November 2022).
- Jiao, Z.T.; Luo, Q. Molecular Mechanisms and Health Benefits of Ghrelin: A Narrative Review. Nutrients 2022, 14, 4191. [Google Scholar] [CrossRef] [PubMed]
- Ukkola, O.; Ravussin, E.; Jacobson, P.; Snyder, E.E.; Chagnon, M.; SjÖstrÖm, L.; Bouchard, C. RAPID COMMUNICATIONS: Mutations in the Preproghrelin/Ghrelin Gene Associated with Obesity in Humans. J. Clin. Endocrinol. Metab. 2001, 86, 3996–3999. [Google Scholar] [CrossRef]
- Steinle, N.I.; Pollin, T.I.; O’Connell, J.R.; Mitchell, B.D.; Shuldiner, A.R. Variants in the Ghrelin Gene Are Associated with Metabolic Syndrome in the Old Order Amish. J. Clin. Endocrinol. Metab. 2005, 90, 6672–6677. [Google Scholar] [CrossRef] [PubMed]
- Gueorguiev, M.; Lecoeur, C.; Meyre, D.; Benzinou, M.; Mein, C.A.; Hinney, A.; Vatin, V.; Weill, J.; Heude, B.; Hebebrand, J.; et al. Association Studies on Ghrelin and Ghrelin Receptor Gene Polymorphisms with Obesity. Obesity 2009, 17, 745–754. [Google Scholar] [CrossRef]
- Chung, W.K.; Patki, A.; Matsuoka, N.; Boyer, B.B.; Liu, N.; Musani, S.K.; Goropashnaya, A.V.; Tan, P.L.; Katsanis, N.; Johnson, S.B.; et al. Analysis of 30 Genes (355 SNPS) Related to Energy Homeostasis for Association with Adiposity in European-American and Yup’ik Eskimo Populations. Hum. Hered. 2009, 67, 193–205. [Google Scholar] [CrossRef]
- Imaizumi, T.; Ando, M.; Nakatochi, M.; Yasuda, Y.; Honda, H.; Kuwatsuka, Y.; Kato, S.; Kondo, T.; Iwata, M.; Nakashima, T.; et al. Effect of Dietary Energy and Polymorphisms in BRAP and GHRL on Obesity and Metabolic Traits. Obes. Res. Clin. Pract. 2018, 12, 39–48. [Google Scholar] [CrossRef]
- Wang, X.; Qu, F.; Wang, C.; Wang, Y.; Wang, D.; Zhao, M.; Yun, X.; Zheng, Q.; Xu, L. Variation Analysis of Ghrelin Gene in Chinese Patients with Obesity, Having Polycystic Ovarian Syndrome. Gynecol. Endocrinol. 2020, 36, 594–598. [Google Scholar] [CrossRef]
- Ando, T.; Komaki, G.; Naruo, T.; Okabe, K.; Takii, M.; Kawai, K.; Konjiki, F.; Takei, M.; Oka, T.; Takeuchi, K.; et al. Possible Role of Preproghrelin Gene Polymorphisms in Susceptibility to Bulimia Nervosa. Am. J. Med. Genet. Part B Neuropsychiatr. Genet. 2006, 141, 929–934. [Google Scholar] [CrossRef]
- GIP Gastric Inhibitory Polypeptide [Homo sapiens (Human)]. Available online: https://www.ncbi.nlm.nih.gov/gene/2695 (accessed on 14 November 2022).
- Chang, C.L.; Cai, J.J.; Cheng, P.J.; Chueh, H.Y.; Hsu, S.Y.T. Identification of Metabolic Modifiers That Underlie Phenotypic Variations in Energy-Balance Regulation. Diabetes 2011, 60, 726–734. [Google Scholar] [CrossRef] [PubMed]
- Nakayama, K.; Watanabe, K.; Boonvisut, S.; Makishima, S.; Miyashita, H.; Iwamoto, S. Common variants of GIP are associated with visceral fat accumulation in Japanese adults. Am. J. Physiol. Gastrointest. Liver. Physiol. 2014, 307, G1108–G1114. [Google Scholar] [CrossRef] [PubMed]
- INS Insulin [Homo sapiens (Human)]. Available online: https://www.ncbi.nlm.nih.gov/gene/3630 (accessed on 14 November 2022).
- Liu, M.; Weiss, M.A.; Arunagiri, A.; Yong, J.; Rege, N.; Sun, J.; Haataja, L.; Kaufman, R.J.; Arvan, P. Biosynthesis, Structure, and Folding of the Insulin Precursor Protein. Diabetes Obes. Metab. 2018, 20, 28–50. [Google Scholar] [CrossRef]
- Vakilian, M.; Tahamtani, Y.; Ghaedi, K. A Review on Insulin Trafficking and Exocytosis. Gene 2019, 706, 52–61. [Google Scholar] [CrossRef]
- Ghosh, S.; Mahalanobish, S.; Sil, P.C. Diabetes: Discovery of Insulin, Genetic, Epigenetic and Viral Infection Mediated Regulation. Nucleus 2021, 65, 283–297. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Sun, J.; Cui, J.; Chen, W.; Guo, H.; Barbetti, F.; Arvan, P. INS-Gene Mutations: From Genetics and Beta Cell Biology to Clinical Disease. Mol. Aspects Med. 2015, 42, 3–18. [Google Scholar] [CrossRef]
- Arneth, B. Insulin Gene Mutations and Posttranslational and Translocation Defects: Associations with Diabetes. Endocrine 2020, 70, 488–497. [Google Scholar] [CrossRef]
- Edghill, E.L.; Flanagan, S.E.; Patch, A.M.; Boustred, C.; Parrish, A.; Shields, B.; Shepherd, M.H.; Hussain, K.; Kapoor, R.R.; Malecki, M.; et al. Insulin Mutation Screening in 1044 Patients with Diabetes Mutations in the INS Gene Are a Common Cause of Neonatal Diabetes but a Rare Cause of Diabetes Diagnosed in Childhood or Adulthood. Diabetes 2008, 57, 1034–1042. [Google Scholar] [CrossRef]
- Boesgaard, T.W.; Pruhova, S.; Andersson, E.A.; Cinek, O.; Obermannova, B.; Lauenborg, J.; Damm, P.; Bergholdt, R.; Pociot, F.; Pisinger, C.; et al. Further Evidence That Mutations in INS Can Be a Rare Cause of Maturity-Onset Diabetes of the Young (MODY). BMC Med. Genet. 2010, 11, 42. [Google Scholar] [CrossRef]
- LEP Leptin [Homo sapiens (Human)]. Available online: https://www.ncbi.nlm.nih.gov/gene/3952 (accessed on 23 December 2022).
- Caldeira, R.S.; Panissa, V.L.G.; Inoue, D.S.; Campos, E.Z.; Monteiro, P.A.; Giglio, B.D.M.; Pimentel, G.D.; Hofmann, P.; Lira, F.S. Impact to short-term high intensity intermittent training on different storages of body fat, leptin and soluble leptin receptor levels in physically active non-obese men: A pilot investigation. Clin. Nutr. ESPEN 2018, 28, 186–192. [Google Scholar] [CrossRef]
- Hamilton, K.; Harvey, J. The neuronal actions of leptin and the implications for treating alzheimer’s disease. Pharmaceuticals 2021, 14, 52. [Google Scholar] [CrossRef] [PubMed]
- Guzmán, A.; Hernández-Coronado, C.G.; Rosales-Torres, A.M.; Hernández-Medrano, J.H. Leptin regulates neuropeptides associated with food intake and GnRH secretion. Ann. Endocrinol. 2019, 80, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Marcos, P.; Coveñas, R. Neuropeptidergic control of feeding: Focus on the galanin family of peptides. Int. J. Mol. Sci. 2021, 22, 2544. [Google Scholar] [CrossRef] [PubMed]
- P41159 LEP_HUMAN. Available online: https://www.uniprot.org/uniprotkb/P41159/entry (accessed on 23 December 2022).
- Eikelis, N.; Lambert, G.; Wiesner, G.; Kaye, D.; Schlaich, M.; Morris, M.; Hastings, J.; Socratous, F.; Esler, M. Extra-adipocyte leptin release in human obesity and its relation to sympathoadrenal function. Am. J. Physiol. Endocrinol. Metab 2004, 286, E744–E752. [Google Scholar] [CrossRef]
- Funcke, J.B.; von Schnurbein, J.; Lennerz, B.; Lahr, G.; Debatin, K.M.; Fischer-Posovszky, P.; Wabitsch, M. Monogenic forms of childhood obesity due to mutations in the leptin gene. Mol. Cell. Pediatr. 2014, 1, 3. [Google Scholar] [CrossRef]
- Dasgupta, S.; Salman, M.; Siddalingaiah, L.B.; Lakshmi, G.L.; Xaviour, D.; Sreenath, J. Genetic variants in leptin: Determinants of obesity and leptin levels in South Indian population. Adipocyte 2014, 4, 135–140. [Google Scholar] [CrossRef]
- Aljanabi, M.A.; Alfaqih, M.A.; Khanfar, M.; Amarin, Z.O.; Elsalem, L.; Saadeh, R.; Al-Mughales, F. Leptin and the GA genotype of rs2167270 of the LEP gene increase the risk of prediabetes. Biomed. Rep. 2021, 14, 44. [Google Scholar] [CrossRef]
- Lombard, Z.; Crowther, N.J.; van der Merwe, L.; Pitamber, P.; Norris, S.A.; Ramsay, M. Appetite regulation genes are associated with body mass index in black South African adolescents: A genetic association study. BMJ Open. 2012, 2, e000873. [Google Scholar] [CrossRef]
- Manju, S.K.; Anilkumar, T.R.; Vysakh, G.; Leena, B.K.; Lekshminarayan, V.; Kumar, P.G.; Shenoy, T.K. A Case-Control Study of the Association of Leptin Gene Polymorphisms with Plasma Leptin Levels and Obesity in the Kerala Population. J. Obes. 2022, 2022, 1040650. [Google Scholar] [CrossRef]
- Ashraf, R.; Khan, M.; Lone, S.; Bhat, M.; Rashid, S.; Majid, S.; Bashir, H. Implication of Leptin and Leptin Receptor Gene Variations in Type 2 Diabetes Mellitus: A Case-Control Study. J. Endocrinol. Metab. 2022, 12, 19–31. [Google Scholar] [CrossRef]
- Enns, J.E.; Taylor, C.G.; Zahradka, P. Variations in Adipokine Genes AdipoQ, Lep, and LepR are Associated with Risk for Obesity-Related Metabolic Disease: The Modulatory Role of Gene-Nutrient Interactions. J. Obes. 2011, 2011, 168659. [Google Scholar] [CrossRef] [PubMed]
- Yaghootkar, H.; Zhang, Y.; Spracklen, C.N.; Karaderi, T.; Huang, L.O.; Bradfield, J.; Schurmann, C.; Fine, R.S.; Preuss, M.H.; Kutalik, Z.; et al. Genetic Studies of Leptin Concentrations Implicate Leptin in the Regulation of Early Adiposity. Diabetes 2020, 69, 2806–2818. [Google Scholar] [CrossRef] [PubMed]
- Haglund, E.; Nguyen, L.; Schafer, N.P.; Lammert, H.; Jennings, P.A.; Onuchic, J.N. Uncovering the molecular mechanisms behind disease-associated leptin variants. J. Biol. Chem. 2018, 293, 12919–12933. [Google Scholar] [CrossRef] [PubMed]
- NAMPT Nicotinamide Phosphoribosyltransferase [Homo sapiens (Human)]. Available online: https://www.ncbi.nlm.nih.gov/gene/10135 (accessed on 14 November 2022).
- Fagerberg, L.; Hallstrom, B.M.; Oksvold, P.; Kampf, C.; Djureinovic, D.; Odeberg, J.; Habuka, M.; Tahmasebpoor, S.; Danielsson, A.; Edlund, K.; et al. Analysis of the Human Tissue-Specific Expression by Genome-Wide Integration of Transcriptomics and Antibody-Based Proteomics. Mol. Cell. Proteom. 2014, 13, 397–406. [Google Scholar] [CrossRef]
- Böttcher, Y.; Teupser, D.; Enigk, B.; Berndt, J.; Klöting, N.; Schön, M.R.; Thiery, J.; Blüher, M.; Stumvoll, M.; Kovacs, P. Genetic Variation in the Visfatin Gene (PBEF1) and Its Relation to Glucose Metabolism and Fat-Depot-Specific Messenger Ribonucleic Acid Expression in Humans. J. Clin. Endocrinol. Metab. 2006, 91, 2725–2731. [Google Scholar] [CrossRef]
- Johansson, L.M.; Johansson, L.E.; Ridderstråle, M. The Visfatin (PBEF1) G-948T Gene Polymorphism Is Associated with Increased High-Density Lipoprotein Cholesterol in Obese Subjects. Metabolism 2008, 57, 1558–1562. [Google Scholar] [CrossRef]
- Blakemore, A.I.F.; Meyre, D.; Delplanque, J.; Vatin, V.; Lecoeur, C.; Marre, M.; Tichet, J.; Balkau, B.; Froguel, P.; Walley, A.J. A Rare Variant in the Visfatin Gene (Nampt/Pbef1) Is Associated with Protection from Obesity. Obesity 2009, 17, 1549–1553. [Google Scholar] [CrossRef]
- Tabassum, R.; Mahendran, Y.; Dwivedi, O.P.; Chauhan, G.; Ghosh, S.; Marwaha, R.K.; Tandon, N.; Bharadwaj, D. Common Variants of IL6, LEPR, and PBEF1 Are Associated with Obesity in Indian Children. Diabetes 2012, 61, 626–631. [Google Scholar] [CrossRef]
- Rong, J.; Chu, M.; Xing, B.; Zhu, L.; Wang, S.; Tao, T.; Zhao, Y.; Jiang, L. Variations in the PBEF1 Gene Are Associated with Body Mass Index: A Population-Based Study in Northern China. Meta Gene 2015, 6, 65–68. [Google Scholar] [CrossRef]
- Zhou, Q.; Chen, B.; Ji, T.; Luo, M.; Luo, J. Association of Genetic Variants in RETN, NAMPT and ADIPOQ Gene with Glycemic, Metabolic Traits and Diabetes Risk in a Chinese Population. Gene 2018, 642, 439–446. [Google Scholar] [CrossRef]
- Kim, J.-E.; Kim, J.-S.; Jo, M.-J.; Cho, E.; Ahn, S.-Y.; Kwon, Y.-J.; Ko, G.-J.T.; Roles, A.; Kim, J.-E.; Kim, J.-S.; et al. The Roles and Associated Mechanisms of Adipokines in Development of Metabolic Syndrome. Molecules 2022, 27, 334. [Google Scholar] [CrossRef]
- PPY Pancreatic Polypeptide [Homo sapiens (Human)]. Available online: https://www.ncbi.nlm.nih.gov/gene/5539 (accessed on 14 November 2022).
- PYY Peptide YY [Homo sapiens (Human)]. Available online: https://www.ncbi.nlm.nih.gov/gene/5697 (accessed on 14 November 2022).
- Simpson, K.; Parker, J.; Plumer, J.; Bloom, S. CCK, PYY and PP: The Control of Energy Balance. Handb. Exp. Pharmacol. 2012, 209, 209–230. [Google Scholar] [CrossRef]
- Friedlander, Y.; Li, G.; Fornage, M.; Williams, O.D.; Lewis, C.E.; Schreiner, P.; Pletcher, M.J.; Enquobahrie, D.; Williams, M.; Siscovick, D.S. Candidate Molecular Pathway Genes Related to Appetite Regulatory Neural Network, Adipocyte Homeostasis and Obesity: Results from the CARDIA Study. Ann. Hum. Genet. 2010, 74, 387–398. [Google Scholar] [CrossRef] [PubMed]
- Siddiq, A.; Gueorguiev, M.; Samson, C.; Hercberg, S.; Heude, B.; Levy-Marchal, C.; Jouret, B.; Weill, J.; Meyre, D.; Walley, A.; et al. Single Nucleotide Polymorphisms in the Neuropeptide Y2 Receptor (NPY2R) Gene and Association with Severe Obesity in French White Subjects. Diabetologia 2007, 50, 574–584. [Google Scholar] [CrossRef]
- Ahituv, N.; Kavaslar, N.; Schackwitz, W.; Ustaszewska, A.; Martin, J.; Hébert, S.; Doelle, H.; Ersoy, B.; Kryukov, G.; Schmidt, S.; et al. Medical Sequencing at the Extremes of Human Body Mass. Am. J. Hum. Genet. 2007, 80, 779–791. [Google Scholar] [CrossRef]
- Perez-Frances, M.; van Gurp, L.; Abate, M.V.; Cigliola, V.; Furuyama, K.; Bru-Tari, E.; Oropeza, D.; Carreaux, T.; Fujitani, Y.; Thorel, F.; et al. Pancreatic Ppy-Expressing γ-Cells Display Mixed Phenotypic Traits and the Adaptive Plasticity to Engage Insulin Production. Nat. Commun. 2021, 12, 4458. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Lee, S.Y.; Kim, C.M. Association between Gene Polymorphisms and Obesity and Physical Fitness in Korean Children. Biol. Sport 2018, 35, 21–27. [Google Scholar] [CrossRef]
- RBP4 Retinol Binding Protein 4 [Homo sapiens (Human)]. Available online: https://www.ncbi.nlm.nih.gov/gene/5950 (accessed on 14 November 2022).
- Yang, Q.; Graham, T.E.; Mody, N.; Preitner, F.; Peroni, O.D.; Zabolotny, J.M.; Kotani, K.; Quadro, L.; Kahn, B.B. Serum Retinol Binding Protein 4 Contributes to Insulin Resistance in Obesity and Type 2 Diabetes. Nature 2005, 436, 356–362. [Google Scholar] [CrossRef]
- Craig, R.L.; Chu, W.S.; Elbein, S.C. Retinol Binding Protein 4 as a Candidate Gene for Type 2 Diabetes and Prediabetic Intermediate Traits. Mol. Genet. Metab. 2007, 90, 338–344. [Google Scholar] [CrossRef]
- Wu, Y.; Li, H.; Loos, R.J.F.; Qi, Q.; Hu, F.B.; Liu, Y.; Lin, X. RBP4 Variants Are Significantly Associated with Plasma RBP4 Levels and Hypertriglyceridemia Risk in Chinese Hans. J. Lipid. Res. 2009, 50, 1479–1486. [Google Scholar] [CrossRef]
- Codoñer-Franch, P.; Carrasco-Luna, J.; Allepuz, P.; Codoñer-Alejos, A.; Guillem, V. Association of RBP4 Genetic Variants with Childhood Obesity and Cardiovascular Risk Factors. Pediatr. Diabetes 2016, 17, 576–583. [Google Scholar] [CrossRef]
- Hu, S.; Ma, S.; Li, X.; Tian, Z.; Liang, H.; Yan, J.; Chen, M.; Tan, H. Relationships of SLC2A4, RBP4, PCK1, and PI3K Gene Polymorphisms with Gestational Diabetes Mellitus in a Chinese Population. Biomed. Res. Int. 2019, 2019, 7398063. [Google Scholar] [CrossRef]
- RETN Resistin [Homo sapiens (Human)]. Available online: https://www.ncbi.nlm.nih.gov/gene/56729 (accessed on 14 November 2022).
- Tripathi, D.; Kant, S.; Pandey, S.; Ehtesham, N.Z. Resistin in Metabolism, Inflammation, and Disease. FEBS J. 2020, 287, 3141–3149. [Google Scholar] [CrossRef] [PubMed]
- Hishida, A.; Wakai, K.; Okada, R.; Morita, E.; Hamajima, N.; Hosono, S.; Higaki, Y.; Turin, T.C.; Suzuki, S.; Motahareh, K.; et al. Significant Interaction between RETN -420 G/G Genotype and Lower BMI on Decreased Risk of Type 2 Diabetes Mellitus (T2DM) in Japanese--the J-MICC Study. Endocr. J. 2013, 60, 237–243. [Google Scholar] [CrossRef] [PubMed]
- Zayani, N.; Omezzine, A.; Boumaiza, I.; Achour, O.; Rebhi, L.; Rejeb, J.; ben Rejeb, N.; ben Abdelaziz, A.; Bouslama, A. Association of ADIPOQ, Leptin, LEPR, and Resistin Polymorphisms with Obesity Parameters in Hammam Sousse Sahloul Heart Study. J. Clin. Lab. Anal. 2017, 31, e22148. [Google Scholar] [CrossRef] [PubMed]
- Beckers, S.; Zegers, D.; van Camp, J.K.; Boudin, E.; Nielsen, T.L.; Brixen, K.; Andersen, M.; van Hul, W. Resistin Polymorphisms Show Associations with Obesity, but Not with Bone Parameters in Men: Results from the Odense Androgen Study. Mol. Biol. Rep. 2013, 40, 2467–2472. [Google Scholar] [CrossRef]
- Chung, C.M.; Lin, T.H.; Chen, J.W.; Leu, H.B.; Yin, W.H.; Ho, H.Y.; Sheu, S.H.; Tsai, W.C.; Chen, J.H.; Lin, S.J.; et al. Common Quantitative Trait Locus Downstream of RETN Gene Identified by Genome-Wide Association Study Is Associated with Risk of Type 2 Diabetes Mellitus in Han Chinese: A Mendelian Randomization Effect. Diabetes Metab. Res. Rev. 2014, 30, 232–240. [Google Scholar] [CrossRef]
- Ortega, L.; Navarro, P.; Riestra, P.; Gavela-Pérez, T.; Soriano-Guillén, L.; Garcés, C. Association of Resistin Polymorphisms with Resistin Levels and Lipid Profile in Children. Mol. Biol. Rep. 2014, 41, 7659–7664. [Google Scholar] [CrossRef]
- Nakatochi, M.; Ichihara, S.; Yamamoto, K.; Ohnaka, K.; Kato, Y.; Yokota, S.; Hirashiki, A.; Naruse, K.; Asano, H.; Izawa, H.; et al. Epigenome-Wide Association Study Suggests That SNPs in the Promoter Region of RETN Influence Plasma Resistin Level via Effects on DNA Methylation at Neighbouring Sites. Diabetologia 2015, 58, 2781–2790. [Google Scholar] [CrossRef]
- Elkhattabi, L.; Morjane, I.; Charoute, H.; Amghar, S.; Bouafi, H.; Elkarhat, Z.; Saile, R.; Rouba, H.; Barakat, A. In Silico Analysis of Coding/Noncoding SNPs of Human RETN Gene and Characterization of Their Impact on Resistin Stability and Structure. J. Diabetes Res. 2019, 2019, 4951627. [Google Scholar] [CrossRef]
- SCT Secretin [Homo sapiens (Human)]. Available online: https://www.ncbi.nlm.nih.gov/gene/6343 (accessed on 14 November 2022).
- Laurila, S.; Rebelos, E.; Honka, M.J.; Nuutila, P. Pleiotropic Effects of Secretin: A Potential Drug Candidate in the Treatment of Obesity? Front Endocrinol. 2021, 12, 1259. [Google Scholar] [CrossRef]
- Laurila, S.; Sun, L.; Lahesmaa, M.; Schnabl, K.; Laitinen, K.; Klén, R.; Li, Y.; Balaz, M.; Wolfrum, C.; Steiger, K.; et al. Secretin Activates Brown Fat and Induces Satiation. Nat. Metab. 2021, 3, 798–809. [Google Scholar] [CrossRef] [PubMed]
- UCP2 Uncoupling Protein 2 [Homo sapiens (Human)]. Available online: https://www.ncbi.nlm.nih.gov/gene/7351 (accessed on 14 November 2022).
- Demine, S.; Renard, P.; Arnould, T. Mitochondrial Uncoupling: A Key Controller of Biological Processes in Physiology and Diseases. Cells 2019, 8, 795. [Google Scholar] [CrossRef] [PubMed]
- Esterbauer, H.; Schneitler, C.; Oberkofler, H.; Ebenbichler, C.; Paulweber, B.; Sandhofer, F.; Ladurner, G.; Hell, E.; Strosberg, A.D.; Patsch, J.R.; et al. A Common Polymorphism in the Promoter of UCP2 Is Associated with Decreased Risk of Obesity in Middle-Aged Humans. Nat. Genet. 2001, 28, 178–183. [Google Scholar] [CrossRef] [PubMed]
- Bulotta, A.; Ludovico, O.; Coco, A.; di Paola, R.; Quattrone, A.; Carella, M.; Pellegrini, F.; Prudente, S.; Trischitta, V. The Common -866G/A Polymorphism in the Promoter Region of the UCP-2 Gene Is Associated with Reduced Risk of Type 2 Diabetes in Caucasians from Italy. J. Clin. Endocrinol. Metab. 2005, 90, 1176–1180. [Google Scholar] [CrossRef]
- Andersen, G.; Dalgaard, L.T.; Justesen, J.M.; Anthonsen, S.; Nielsen, T.; Thørner, L.W.; Witte, D.; Jørgensen, T.; Clausen, J.O.; Lauritzen, T.; et al. The Frequent UCP2-866G>A Polymorphism Protects against Insulin Resistance and Is Associated with Obesity: A Study of Obesity and Related Metabolic Traits among 17 636 Danes. Int. J. Obes. 2013, 37, 175–181. [Google Scholar] [CrossRef]
- Salopuro, T.; Pulkkinen, L.; Lindström, J.; Kolehmainen, M.; Tolppanen, A.M.; Eriksson, J.G.; Valle, T.T.; Aunola, S.; Ilanne-Parikka, P.; Keinänen-Kiukaanniemi, S. Variation in the UCP2 and UCP3 genes associates with abdominal obesity and serum lipids: The Finnish Diabetes Prevention Study. BMC Med. Genet. 2009, 10, 94. [Google Scholar] [CrossRef]
- Oktavianthi, S.; Trimarsanto, H.; Febinia, C.A.; Suastika, K.; Saraswati, M.R.; Dwipayana, P.; Arindrarto, W.; Sudoyo, H.; Malik, S.G. Uncoupling Protein 2 Gene Polymorphisms Are Associated with Obesity. Cardiovasc. Diabetol. 2012, 11, 41. [Google Scholar] [CrossRef]
- Martinez-Hervas, S.; Mansego, M.L.; de Marco, G.; Martinez, F.; Alonso, M.P.; Morcillo, S.; Rojo-Martinez, G.; Real, J.T.; Ascaso, J.F.; Redon, J.; et al. Polymorphisms of the UCP2 Gene Are Associated with Body Fat Distribution and Risk of Abdominal Obesity in Spanish Population. Eur. J. Clin. Invest. 2012, 42, 171–178. [Google Scholar] [CrossRef]
- Xu, L.; Chen, S.; Zhan, L. Association of Uncoupling Protein-2 -866G/A and Ala55Val Polymorphisms with Susceptibility to Type 2 Diabetes Mellitus: A Meta-Analysis of Case-Control Studies. Medicine 2021, 100, e24464. [Google Scholar] [CrossRef]
- Fall, T.; Mendelson, M.; Speliotes, E.K. Recent Advances in Human Genetics and Epigenetics of Adiposity: Pathway to Precision Medicine? Gastroenterology 2017, 152, 1695–1706. [Google Scholar] [CrossRef] [PubMed]
- Christakoudi, S.; Tsilidis, K.K.; Evangelou, E.; Riboli, E. A Body Shape Index (ABSI), hip index, and risk of cancer in the UK Biobank cohort. Cancer Med. 2021, 10, 5614–5628. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Hsu, L.-A.; Teng, M.-S.; Chou, H.-H.; Ko, Y.-L. Differential Genetic and Epigenetic Effects of the KLF14 Gene on Body Shape Indices and Metabolic Traits. Int. J. Mol. Sci. 2022, 23, 4165. [Google Scholar] [CrossRef] [PubMed]
- Nagayama, D.; Sugiura, T.; Choi, S.Y.; Shirai, K. Various Obesity Indices and Arterial Function Evaluated with CAVI—Is Waist Circumference Adequate to Define Metabolic Syndrome? Vasc. Health Risk. Manag. 2022, 18, 721–733. [Google Scholar] [CrossRef] [PubMed]
- Nagayama, D.; Fujishiro, K.; Watanabe, Y.; Yamaguchi, T.; Suzuki, K.; Saiki, A.; Shirai, K. A Body Shape Index (ABSI) as a Variant of Conicity Index Not Affected by the Obesity Paradox: A Cross-Sectional Study Using Arterial Stiffness Parameter. J. Pers. Med. 2022, 12, 2014. [Google Scholar] [CrossRef]
- Young, K.L.; Graff, M.; Fernandez-Rhodes, L.; North, K.E. Genetics of Obesity in Diverse Populations. Curr. Diab. Rep. 2018, 18, 145. [Google Scholar] [CrossRef] [PubMed]
- Locke, A.E.; Kahali, B.; Berndt, S.I.; Justice, A.E.; Pers, T.H.; Day, F.R.; Powell, C.; Vedantam, S.; Buchkovich, M.L.; Yang, J.; et al. Genetic studies of body mass index yield new insights for obesity biology. Nature 2015, 518, 197–206. [Google Scholar] [CrossRef]
- Pan, D.Z.; Miao, Z.; Comenho, C.; Rajkumar, S.; Koka, A.; Lee, S.H.T.; Alvarez, M.; Kaminska, D.; Ko, A.; Sinsheimer, J.S.; et al. Identification of TBX15 as an adipose master trans regulator of abdominal obesity genes. Genome Med. 2021, 13, 123. [Google Scholar] [CrossRef]
- Larsson, S.C.; Bäck, M.; Rees, J.M.B.; Mason, A.M.; Burgess, S. Body mass index and body composition in relation to 14 cardiovascular conditions in UK Biobank: A Mendelian randomization study. Eur. Heart J. 2020, 41, 221–226. [Google Scholar] [CrossRef]
- Beaney, K.E.; Cooper, J.A.; Shahid, S.U.; Ahmed, W.; Qamar, R.; Drenos, F.; Crockard, M.A.; Humphries, S.E. Clinical Utility of a Coronary Heart Disease Risk Prediction Gene Score in UK Healthy Middle-Aged Men and in the Pakistani Population. PLoS ONE 2015, 10, e0130754. [Google Scholar] [CrossRef]
- Iribarren, C.; Lu, M.; Jorgenson, E.; Martínez, M.; Lluis-Ganella, C.; Subirana, I.; Salas, E.; Elosua, R. Weighted Multi-marker Genetic Risk Scores for Incident Coronary Heart Disease among Individuals of African, Latino and East-Asian Ancestry. Sci. Rep. 2018, 8, 6853. [Google Scholar] [CrossRef]
- Semaev, S.; Shakhtshneider, E. Genetic Risk Score for Coronary Heart Disease: Review. J. Pers. Med. 2020, 10, 239. [Google Scholar] [CrossRef] [PubMed]
- Benbaibeche, H.; Bounihi, A.; Koceir, E.A. Leptin level as a biomarker of uncontrolled eating in obesity and overweight. Ir. J. Med. Sci. 2020, 190, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Poetsch, M.S.; Strano, A.; Guan, K. Role of Leptin in Cardiovascular Diseases. Front. Endocrinol. 2020, 11, 354. [Google Scholar] [CrossRef]
- Reyes-Barrera, J.; Sainz-Escárrega, V.H.; Medina-Urritia, A.X.; Jorge-Galarza, E.; Osorio-Alonso, H.; Torres-Tamayo, M.; Leal-Escobar, G.; Posadas-Romero, C.; Torre-Villalvazo, I.; Juárez-Rojas, J.G. Dysfunctional adiposity index as a marker of adipose tissue morpho-functional abnormalities and metabolic disorders in apparently healthy subjects. Adipocyte 2021, 10, 142–152. [Google Scholar] [CrossRef] [PubMed]
- Trinh, T.; Broxmeyer, H.E. Role for Leptin and Leptin Receptors in Stem Cells During Health and Diseases. Stem. Cell Rev. Rep. 2021, 17, 511–522. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).