
Regulation of Key Immune-Related Genes in the Heart Following Burn Injury



Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Animals, Burn Preformation and Sample Collection
2.3. RNA Isolation
2.4. First-Strand cDNA Synthesis
2.5. Real-Time RT² Profiler PCR Array (QIAGEN, Cat. no. PARN-018Z)
2.6. Analysis of Real-Time RT² Profiler PCR Array
2.7. String-DB Analysis of DEGs
2.8. Signaling Pathway Mapping for DEGs
2.9. Validation of Identified Genes by qPCR
2.10. Protein–Protein Interaction (PPI) Networks
2.11. Statistical Analysis
3. Results
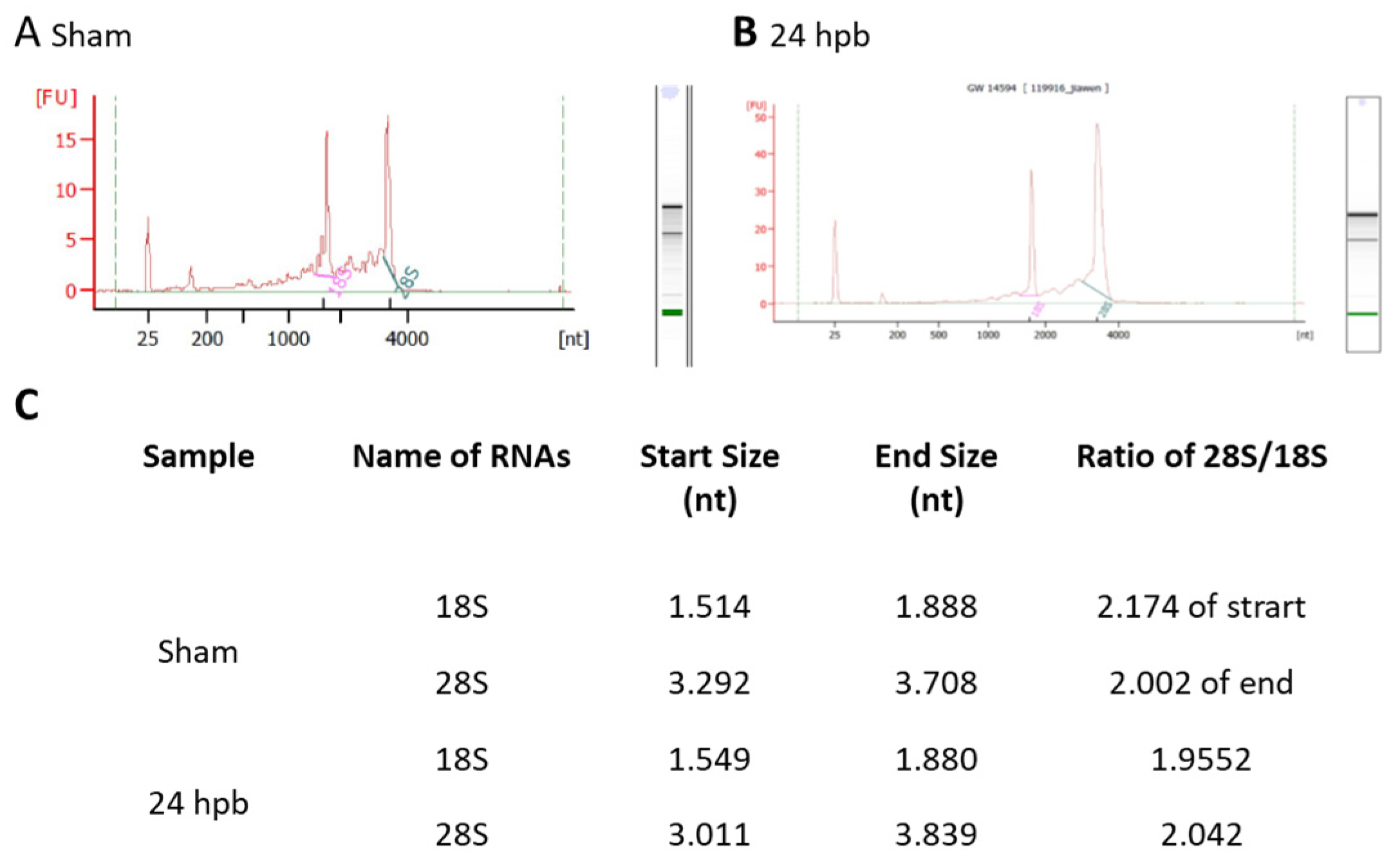
3.1. Highly Qualified RNA
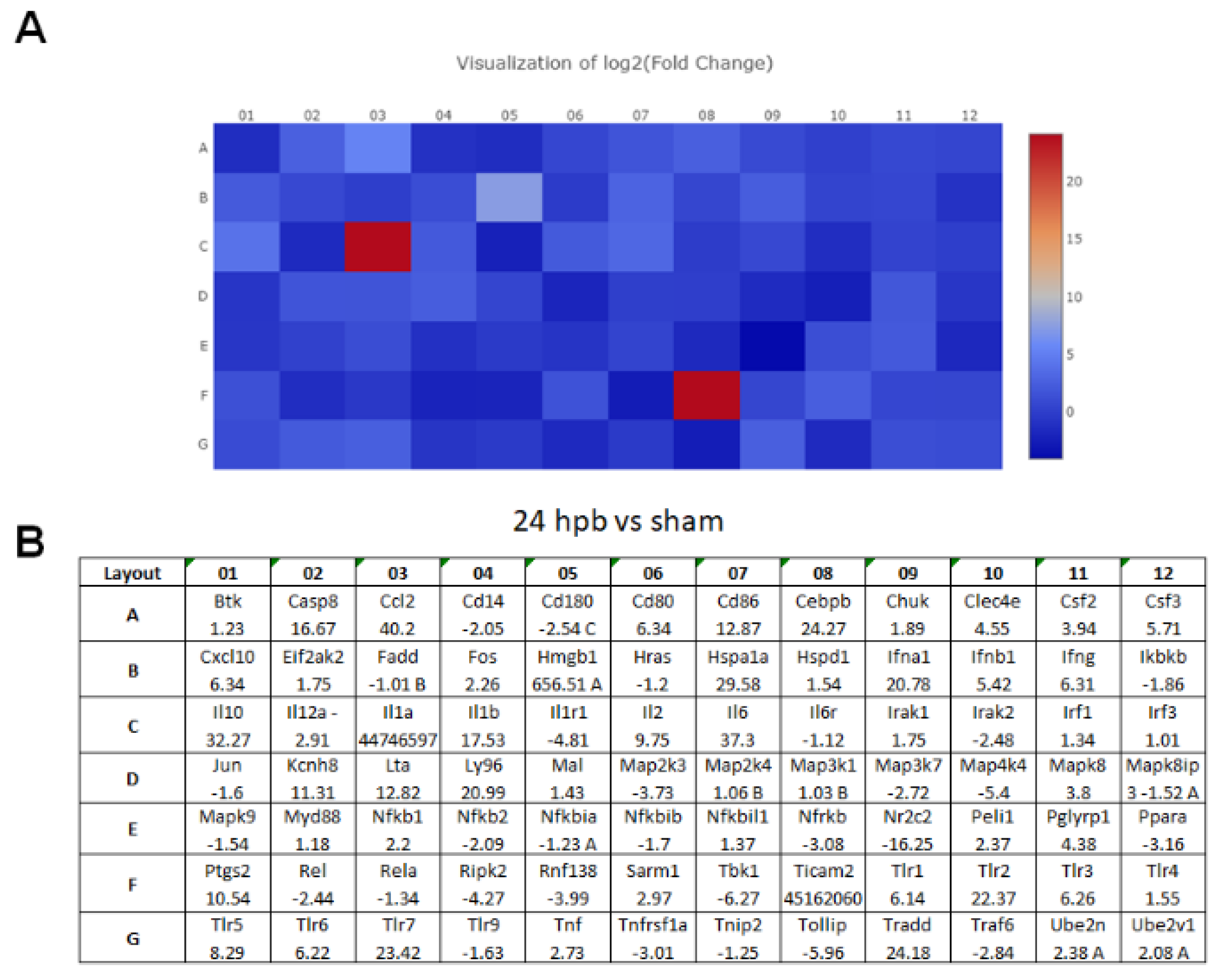
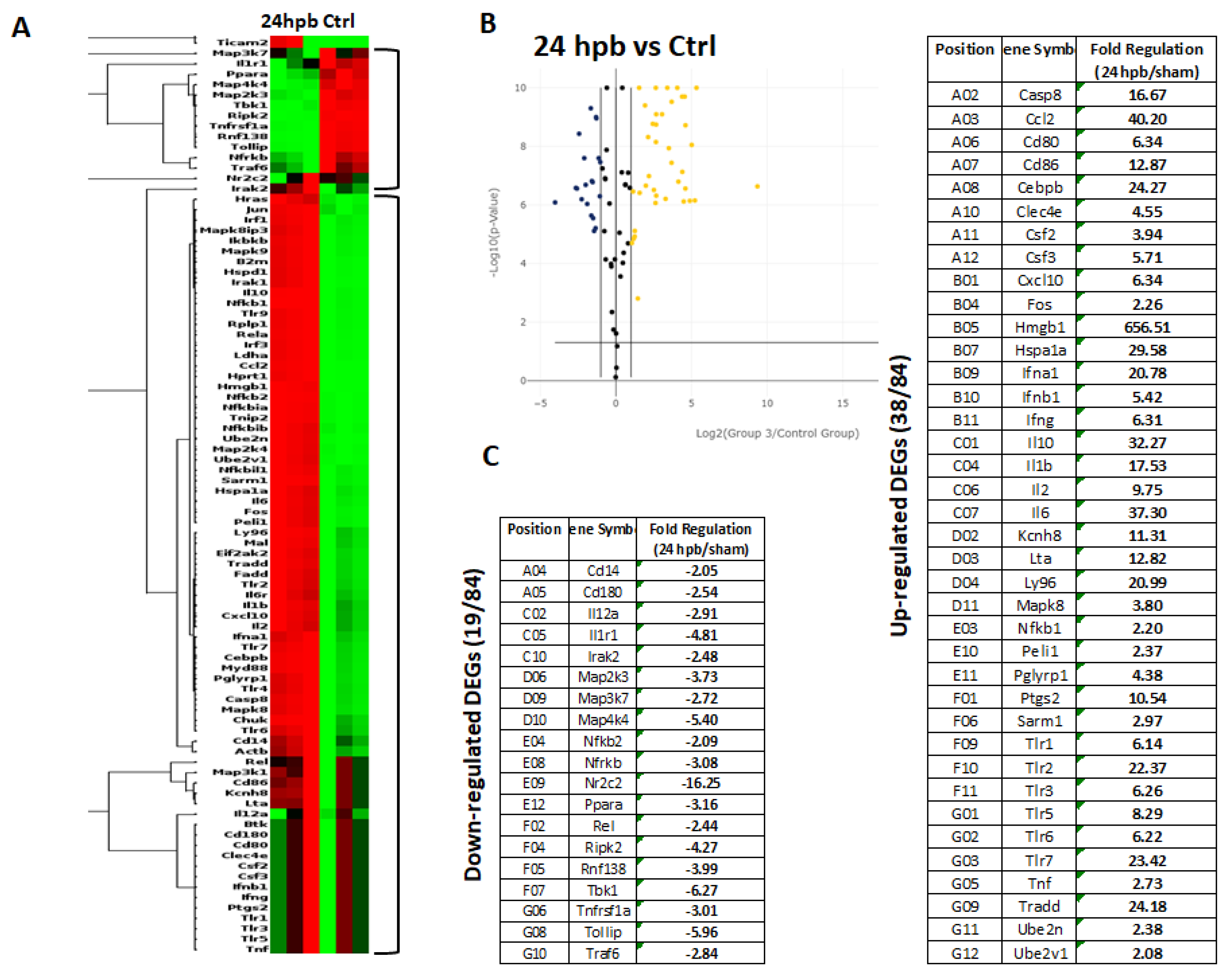
3.2. PCR Array in Cardiomyocytes
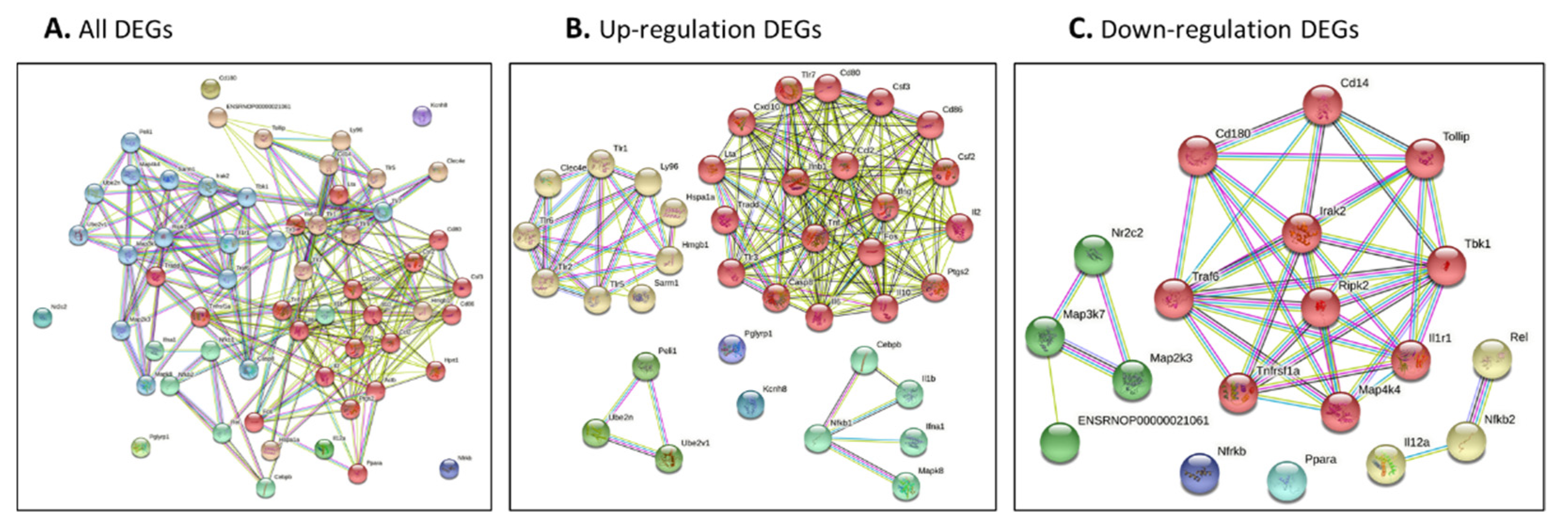
3.3. Analysis of Interaction Network and Function Network of Identified Genes after Burn Injury
3.3.1. Protein–Protein Interaction Network and Function Network of Identified Up-Regulated Genes after Burn Injury
3.3.2. Protein–Protein Interaction Network and Function Network of Identified Down-Regulated Genes after Burn Injury
3.4. The Analysis of Enriched Gene Ontology (GO) for Identified DEGs
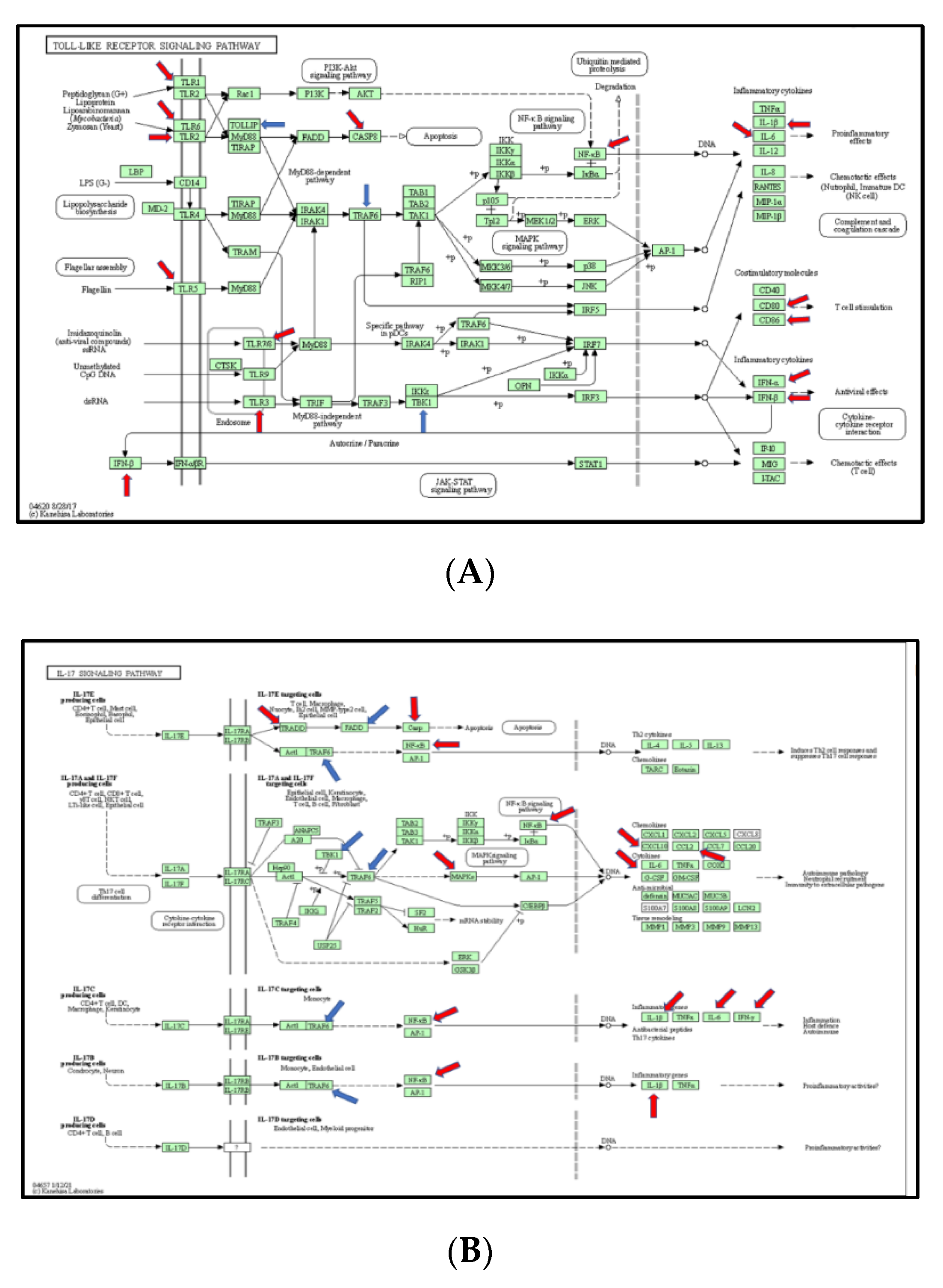
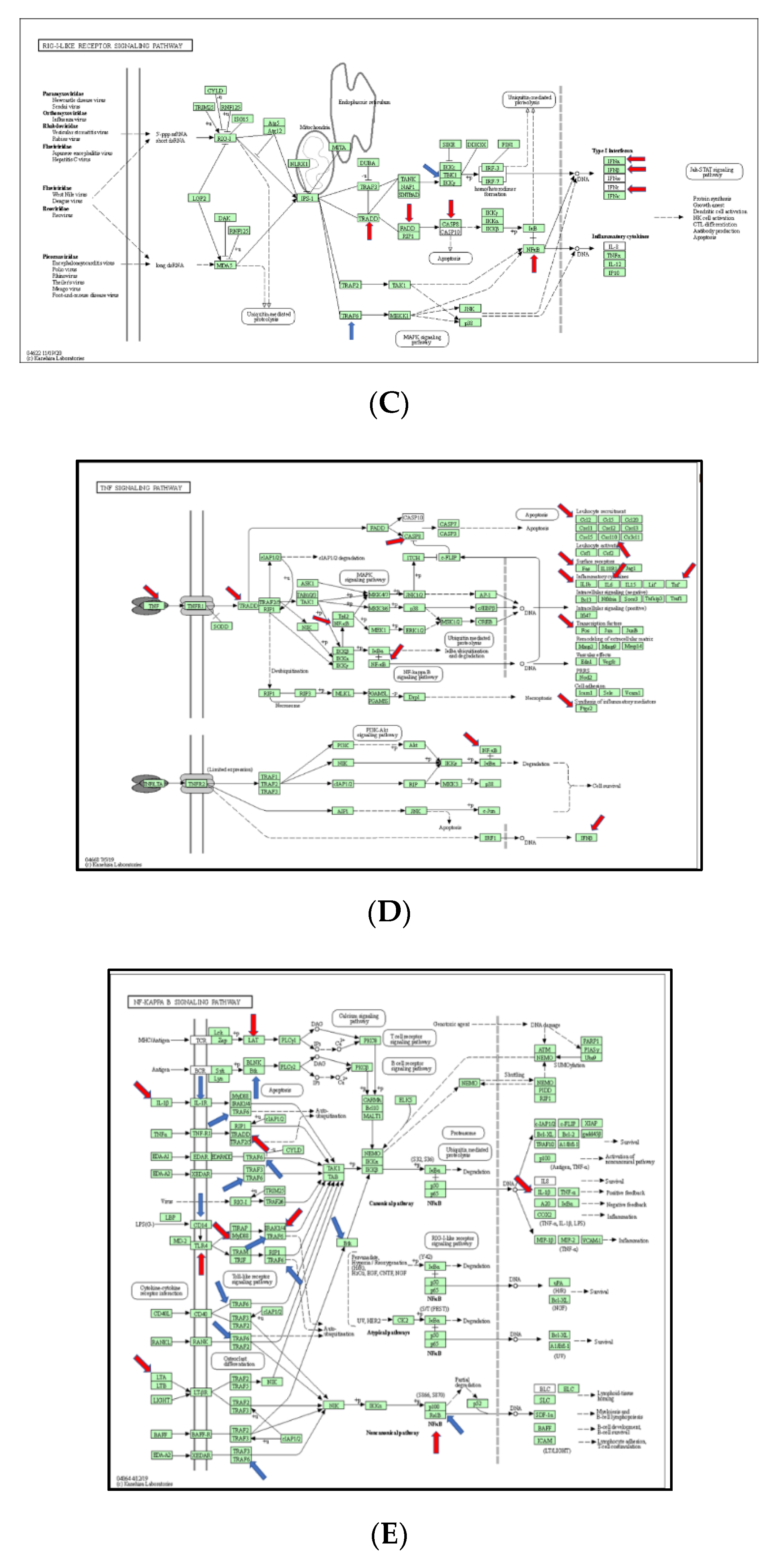
3.5. The Signaling Pathway Mapping for Identified DEGs
3.6. Function Analysis of Burn-Injury-Induced DEGs Related TLR Pathway
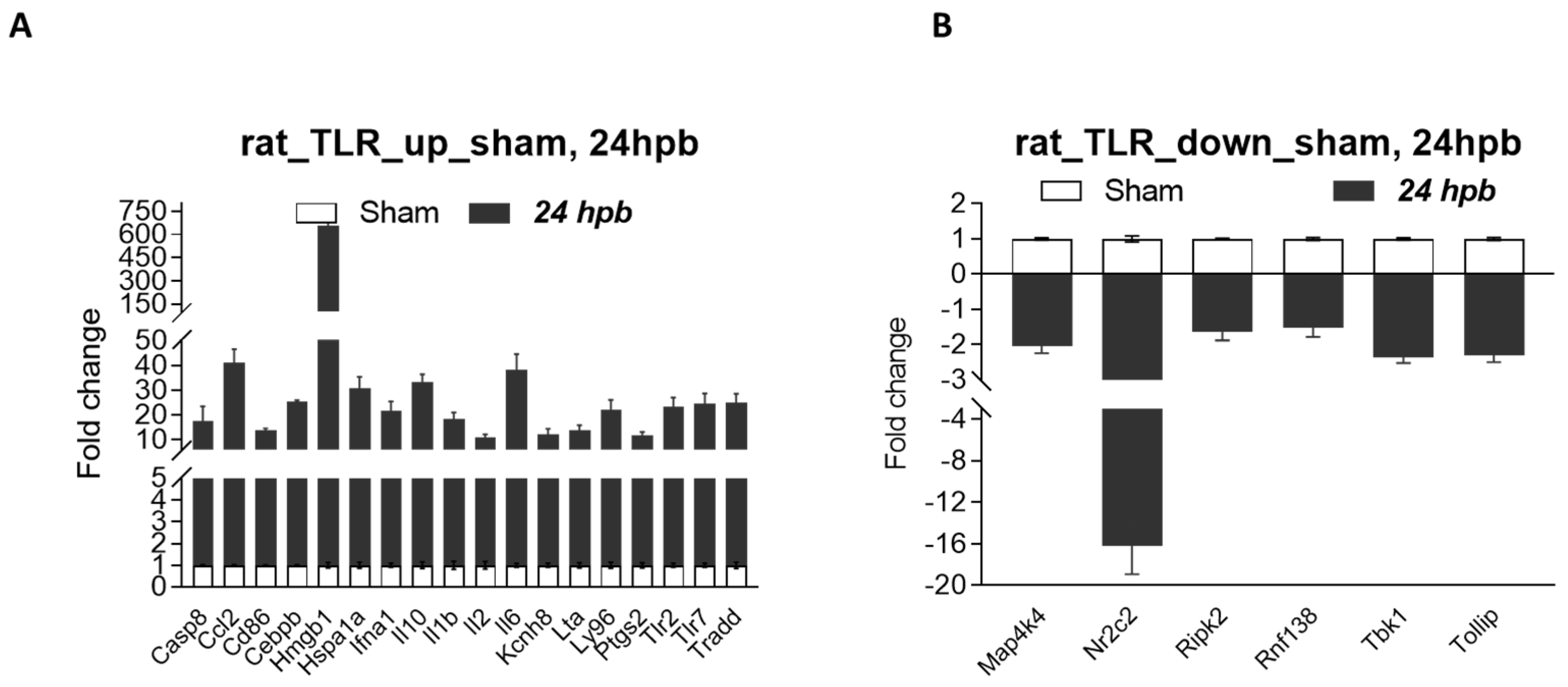
3.7. Validation of Burn-Injury-Induced TLR-Related DEGs Using qPCR
3.8. Characterizing Protein–Protein Interactions through STRING Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Translational Relevance
Abbreviations
| DAMP | damage-associated molecular pattern |
| DEG | differentially expressed gene |
| DEP | differentially expressed protein |
| Hpb | hours post-burn |
| Hmgb1 | high-mobility group box 1 |
| IL | interleukin |
| I/R | ischemia/reperfusion |
| MAP4K4 | mitogen-activated protein kinase kinase kinase kinase-4 |
| PAMP | pathogen-associated molecular pattern |
| PPI | protein–protein interaction |
| PRR | pattern-recognition receptor |
| Ripk2 | receptor-interacting serine-threonine protein kinase 2 |
| TBK1 | TANK-binding kinase 1 |
| TBSA | total body surface area |
| TLR | Toll-like receptors |
| Ube2n | ubiquitin-conjugating enzyme E2 N |
| Ube2v1 | ubiquitin-conjugating enzyme E2 variant 1 |
References
- Adams, H.R.; Baxter, C.R.; Izenberg, S.D. Decreased contractility and compliance of the left ventricle as complications of thermal trauma. Am. Heart J. 1984, 108, 1477–1487. [Google Scholar] [CrossRef]
- Horton, J.W.; Maass, D.L.; White, J.; Sanders, B. Myocardial inflammatory responses to sepsis complicated by previous burn injury. Surg. Infect. 2003, 4, 363–377. [Google Scholar] [CrossRef] [Green Version]
- Horton, J.W.; Maass, D.L.; White, J.; Sanders, B. Hypertonic saline-dextran suppresses burn-related cytokine secretion by cardiomyocytes. Am. J. Physiol. Heart Circ. Physiol. 2001, 280, H1591–H1601. [Google Scholar] [CrossRef]
- Maass, D.L.; Hybki, D.P.; White, J.; Horton, J.W. The time course of cardiac NF-kappaB activation and TNF-alpha secretion by cardiac myocytes after burn injury: Contribution to burn-related cardiac contractile dysfunction. Shock 2002, 17, 293–299. [Google Scholar] [CrossRef]
- Ballard-Croft, C.; White, D.J.; Maass, D.L.; Hybki, D.P.; Horton, J.W. Role of p38 mitogen-activated protein kinase in cardiac myocyte secretion of the inflammatory cytokine TNF-alpha. Am. J. Physiol. Heart Circ. Physiol. 2001, 280, H1970–H1981. [Google Scholar] [CrossRef]
- Hartman, M.H.T.; Groot, H.E.; Leach, I.M.; Karper, J.C.; van der Harst, P. Translational overview of cytokine inhibition in acute myocardial infarction and chronic heart failure. Trends Cardiovasc. Med. 2018, 28, 369–379. [Google Scholar] [CrossRef]
- Wen, J.J.; Williams, T.P.; Cummins, C.B.; Colvill, K.M.; Radhakrishnan, G.L.; Radhakrishnan, R.S. Effect of Mitochondrial Antioxidant (Mito-TEMPO) on Burn-Induced Cardiac Dysfunction. J. Am. Coll. Surg. 2021, 232, 642–655. [Google Scholar] [CrossRef]
- Wen, J.J.; Cummins, C.B.; Williams, T.P.; Radhakrishnan, R.S. The Genetic Evidence of Burn-Induced Cardiac Mitochondrial Metabolism Dysfunction. Biomedicines 2020, 8, 566. [Google Scholar] [CrossRef]
- Wen, J.J.; Cummins, C.B.; Szczesny, B.; Radhakrishnan, R.S. Cardiac Dysfunction after Burn Injury: Role of the AMPK-SIRT1-PGC1alpha-NFE2L2-ARE Pathway. J. Am. Coll. Surg. 2020, 230, 562–571. [Google Scholar] [CrossRef]
- Wen, J.J.; Cummins, C.B.; Radhakrishnan, R.S. Burn-Induced Cardiac Mitochondrial Dysfunction via Interruption of the PDE5A-cGMP-PKG Pathway. Int. J. Mol. Sci. 2020, 21, 2350. [Google Scholar] [CrossRef] [Green Version]
- Wen, J.J.; Cummins, C.; Radhakrishnan, R.S. Sildenafil Recovers Burn-Induced Cardiomyopathy. Cells 2020, 9, 1393. [Google Scholar] [CrossRef] [PubMed]
- Kawai, T.; Akira, S. The role of pattern-recognition receptors in innate immunity: Update on Toll-like receptors. Nat. Immunol. 2010, 11, 373–384. [Google Scholar] [CrossRef] [PubMed]
- Zitvogel, L.; Kepp, O.; Kroemer, G. Decoding cell death signals in inflammation and immunity. Cell 2010, 140, 798–804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, S.; Yu, M.; Fang, Q.; Zhang, L.; You, C.; Wang, X.; Liu, Y.; Han, C. Heme oxygenase-1 induction mitigates burn-associated early acute kidney injury via the TLR4 signaling pathway. Burns 2022, 48, 156–167. [Google Scholar] [CrossRef]
- Cornick, S.M.; Noronha, S.A.; Noronha, S.M.; Cezillo, M.V.; Ferreira, L.M.; Gragnani, A. Toll like receptors gene expression of human keratinocytes cultured of severe burn injury. Acta Cir. Bras. 2014, 29 (Suppl. S3), 33–38. [Google Scholar] [CrossRef] [Green Version]
- Finnerty, C.C.; Jeschke, M.G.; Branski, L.K.; Barret, J.P.; Dziewulski, P.; Herndon, D.N. Hypertrophic scarring: The greatest unmet challenge after burn injury. Lancet 2016, 388, 1427–1436. [Google Scholar] [CrossRef] [Green Version]
- Newton, K.; Dixit, V.M. Signaling in innate immunity and inflammation. Cold Spring Harb. Perspect. Biol. 2012, 4, a006049. [Google Scholar] [CrossRef]
- O’Neill, L.A.; Golenbock, D.; Bowie, A.G. The history of Toll-like receptors-redefining innate immunity. Nat. Rev. Immunol. 2013, 13, 453–460. [Google Scholar] [CrossRef]
- Hoesel, L.M.; Niederbichler, A.D.; Schaefer, J.; Ipaktchi, K.R.; Gao, H.; Rittirsch, D.; Pianko, M.J.; Vogt, P.M.; Sarma, J.V.; Su, G.L.; et al. C5a-blockade improves burn-induced cardiac dysfunction. J. Immunol. 2007, 178, 7902–7910. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Andersen, C.L.; Jensen, J.L.; Orntoft, T.F. Normalization of real-time quantitative reverse transcription-PCR data: A model-based variance estimation approach to identify genes suited for normalization, applied to bladder and colon cancer data sets. Cancer Res. 2004, 64, 5245–5250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keyloun, J.W.; Campbell, R.; Carney, B.C.; Yang, R.; Miller, S.A.; Detwiler, L.; Gautam, A.; Moffatt, L.T.; Hammamieh, R.; Jett, M.; et al. Early Transcriptomic Response to Burn injury: Severe Burns are Associated with Immune Pathway Shutdown. J. Burn Care Res. 2021, 43, 306–314. [Google Scholar] [CrossRef] [PubMed]
- Shen, H.; de Almeida, P.E.; Kang, K.H.; Yao, P.; Chan, C.W. Burn injury triggered dysfunction in dendritic cell response to TLR9 activation and resulted in skewed T cell functions. PLoS ONE 2012, 7, e50238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valvis, S.M.; Waithman, J.; Wood, F.M.; Fear, M.W.; Fear, V.S. The Immune Response to Skin Trauma Is Dependent on the Etiology of Injury in a Mouse Model of Burn and Excision. J. Investig. Dermatol. 2015, 135, 2119–2128. [Google Scholar] [CrossRef] [Green Version]
- Matsuura, H.; Matsumoto, H.; Osuka, A.; Ogura, H.; Shimizu, K.; Kang, S.; Tanaka, T.; Ueyama, M.; Shimazu, T. Clinical Importance of a Cytokine Network in Major Burns. Shock 2019, 51, 185–193. [Google Scholar] [CrossRef]
- Johnson, B.Z.; McAlister, S.; McGuire, H.M.; Palanivelu, V.; Stevenson, A.; Richmond, P.; Palmer, D.J.; Metcalfe, J.; Prescott, S.L.; Wood, F.M.; et al. Corrigendum: Pediatric Burn Survivors Have Long-Term Immune Dysfunction with Diminished Vaccine Response. Front. Immunol. 2020, 11, 598646. [Google Scholar] [CrossRef]
- Parment, K.; Zetterberg, A.; Ernerudh, J.; Bakteman, K.; Steinwall, I.; Sjoberg, F. Long-term immunosuppression in burned patients assessed by in vitro neutrophil oxidative burst (Phagoburst). Burns 2007, 33, 865–871. [Google Scholar] [CrossRef]
- Wen, H.; Xing, L.; Sun, K.; Xiao, C.; Meng, X.; Yang, J. Loganin attenuates intestinal injury in severely burned rats by regulating the toll-like receptor 4/NF-kappaB signaling pathway. Exp. Ther. Med. 2020, 20, 591–598. [Google Scholar] [CrossRef]
- D’Arpa, P.; Leung, K.P. Toll-Like Receptor Signaling in Burn Wound Healing and Scarring. Adv. Wound Care 2017, 6, 330–343. [Google Scholar] [CrossRef] [Green Version]
- Shupp, J.W.; Nasabzadeh, T.J.; Rosenthal, D.S.; Jordan, M.H.; Fidler, P.; Jeng, J.C. A review of the local pathophysiologic bases of burn wound progression. J. Burn Care Res. 2010, 31, 849–873. [Google Scholar] [CrossRef]
- Jaskille, A.D.; Jeng, J.C.; Sokolich, J.C.; Lunsford, P.; Jordan, M.H. Repetitive ischemia-reperfusion injury: A plausible mechanism for documented clinical burn-depth progression after thermal injury. J. Burn Care Res. 2007, 28, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Gill, R.; Tsung, A.; Billiar, T. Linking oxidative stress to inflammation: Toll-like receptors. Free Radic. Biol. Med. 2010, 48, 1121–1132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lanier, S.T.; McClain, S.A.; Lin, F.; Singer, A.J.; Clark, R.A. Spatiotemporal progression of cell death in the zone of ischemia surrounding burns. Wound Repair Regen. 2011, 19, 622–632. [Google Scholar] [CrossRef] [Green Version]
- Venereau, E.; Schiraldi, M.; Uguccioni, M.; Bianchi, M.E. HMGB1 and leukocyte migration during trauma and sterile inflammation. Mol. Immunol. 2013, 55, 76–82. [Google Scholar] [CrossRef] [PubMed]
- Chavez-Sanchez, L.; Madrid-Miller, A.; Chavez-Rueda, K.; Legorreta-Haquet, M.V.; Tesoro-Cruz, E.; Blanco-Favela, F. Activation of TLR2 and TLR4 by minimally modified low-density lipoprotein in human macrophages and monocytes triggers the inflammatory response. Hum. Immunol. 2010, 71, 737–744. [Google Scholar] [CrossRef] [PubMed]
- Greco, J.A., 3rd; Pollins, A.C.; Boone, B.E.; Levy, S.E.; Nanney, L.B. A microarray analysis of temporal gene expression profiles in thermally injured human skin. Burns 2010, 36, 192–204. [Google Scholar] [CrossRef] [Green Version]
- Horton, J.W.; Maass, D.L.; White, D.J.; Sanders, B.; Murphy, J. Effects of burn serum on myocardial inflammation and function. Shock 2004, 22, 438–445. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.; Maass, D.L.; White, D.J.; Horton, J.W. Effects of burn injury on myocardial signaling and cytokine secretion: Possible role of PKC. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2007, 292, R887–R896. [Google Scholar] [CrossRef]
- Jeschke, M.G.; Chinkes, D.L.; Finnerty, C.C.; Kulp, G.; Suman, O.E.; Norbury, W.B.; Branski, L.K.; Gauglitz, G.G.; Mlcak, R.P.; Herndon, D.N. Pathophysiologic response to severe burn injury. Ann. Surg. 2008, 248, 387–401. [Google Scholar] [CrossRef] [Green Version]
- Porter, C.; Tompkins, R.G.; Finnerty, C.C.; Sidossis, L.S.; Suman, O.E.; Herndon, D.N. The metabolic stress response to burn trauma: Current understanding and therapies. Lancet 2016, 388, 1417–1426. [Google Scholar] [CrossRef] [Green Version]
- Tompkins, R.G. Genomics of injury: The Glue Grant experience. J. Trauma Acute Care Surg. 2015, 78, 671–686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saxena, A.; Russo, I.; Frangogiannis, N.G. Inflammation as a therapeutic target in myocardial infarction: Learning from past failures to meet future challenges. Transl. Res. 2016, 167, 152–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hage, C.; Michaelsson, E.; Linde, C.; Donal, E.; Daubert, J.C.; Gan, L.M.; Lund, L.H. Inflammatory Biomarkers Predict Heart Failure Severity and Prognosis in Patients with Heart Failure With Preserved Ejection Fraction: A Holistic Proteomic Approach. Circ. Cardiovasc. Genet. 2017, 10, e001633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ridiandries, A.; Tan, J.T.M.; Bursill, C.A. The Role of Chemokines in Wound Healing. Int. J. Mol. Sci. 2018, 19, 3217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wahid, A.; Chen, W.; Wang, X.; Tang, X. High-mobility group box 1 serves as an inflammation driver of cardiovascular disease. Biomed Pharm. 2021, 139, 111555. [Google Scholar] [CrossRef] [PubMed]
- Kang, R.; Chen, R.; Zhang, Q.; Hou, W.; Wu, S.; Cao, L.; Huang, J.; Yu, Y.; Fan, X.G.; Yan, Z.; et al. HMGB1 in health and disease. Mol. Asp. Med. 2014, 40, 1–116. [Google Scholar] [CrossRef] [Green Version]
- Funayama, A.; Shishido, T.; Netsu, S.; Narumi, T.; Kadowaki, S.; Takahashi, H.; Miyamoto, T.; Watanabe, T.; Woo, C.H.; Abe, J.; et al. Cardiac nuclear high mobility group box 1 prevents the development of cardiac hypertrophy and heart failure. Cardiovasc. Res. 2013, 99, 657–664. [Google Scholar] [CrossRef] [Green Version]
- Coleman, L.G., Jr.; Maile, R.; Jones, S.W.; Cairns, B.A.; Crews, F.T. HMGB1/IL-1beta complexes in plasma microvesicles modulate immune responses to burn injury. PLoS ONE 2018, 13, e0195335. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.L.; Sun, L.; Guo, F.; Wang, F.; Liu, S.; Liang, X.; Wang, R.S.; Wang, Y.J.; Sun, Y.X. High-mobility group box-1 induces proinflammatory cytokines production of Kupffer cells through TLRs-dependent signaling pathway after burn injury. PLoS ONE 2012, 7, e50668. [Google Scholar] [CrossRef]
- Bujak, M.; Frangogiannis, N.G. The role of IL-1 in the pathogenesis of heart disease. Arch. Immunol. Ther. Exp. 2009, 57, 165–176. [Google Scholar] [CrossRef] [Green Version]
- Ing, D.J.; Zang, J.; Dzau, V.J.; Webster, K.A.; Bishopric, N.H. Modulation of cytokine-induced cardiac myocyte apoptosis by nitric oxide, Bak, and Bcl-x. Circ. Res. 1999, 84, 21–33. [Google Scholar] [CrossRef] [PubMed]
- Maass, D.L.; White, J.; Horton, J.W. IL-1beta and IL-6 act synergistically with TNF-alpha to alter cardiac contractile function after burn trauma. Shock 2002, 18, 360–366. [Google Scholar] [CrossRef] [PubMed]
- Frangogiannis, N.G.; Mendoza, L.H.; Lindsey, M.L.; Ballantyne, C.M.; Michael, L.H.; Smith, C.W.; Entman, M.L. IL-10 is induced in the reperfused myocardium and may modulate the reaction to injury. J. Immunol. 2000, 165, 2798–2808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Welsh, P.; Murray, H.M.; Ford, I.; Trompet, S.; de Craen, A.J.; Jukema, J.W.; Stott, D.J.; McInnes, I.B.; Packard, C.J.; Westendorp, R.G.; et al. Circulating interleukin-10 and risk of cardiovascular events: A prospective study in the elderly at risk. Arter. Thromb. Vasc. Biol. 2011, 31, 2338–2344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frangogiannis, N.G. Cardiac fibrosis: Cell biological mechanisms, molecular pathways and therapeutic opportunities. Mol. Asp. Med. 2019, 65, 70–99. [Google Scholar] [CrossRef]
- Gonzalez, A.; Schelbert, E.B.; Diez, J.; Butler, J. Myocardial Interstitial Fibrosis in Heart Failure: Biological and Translational Perspectives. J. Am. Coll. Cardiol. 2018, 71, 1696–1706. [Google Scholar] [CrossRef]
- Nguyen, M.N.; Kiriazis, H.; Gao, X.M.; Du, X.J. Cardiac Fibrosis and Arrhythmogenesis. Compr. Physiol. 2017, 7, 1009–1049. [Google Scholar] [CrossRef]
- Fiordelisi, A.; Iaccarino, G.; Morisco, C.; Coscioni, E.; Sorriento, D. NFkappaB is a Key Player in the Crosstalk between Inflammation and Cardiovascular Diseases. Int. J. Mol. Sci. 2019, 20, 1599. [Google Scholar] [CrossRef] [Green Version]
- Brasier, A.R. The NF-kappaB regulatory network. Cardiovasc. Toxicol. 2006, 6, 111–130. [Google Scholar] [CrossRef]
- Hayden, M.S.; Ghosh, S. Shared principles in NF-kappaB signaling. Cell 2008, 132, 344–362. [Google Scholar] [CrossRef] [Green Version]
- Ritchie, M.E. Nuclear factor-kappaB is selectively and markedly activated in humans with unstable angina pectoris. Circulation 1998, 98, 1707–1713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, N.; Gulick, J.; Osinska, H.; Yu, Y.; McLendon, P.M.; Shay-Winkler, K.; Robbins, J.; Yutzey, K.E. Ube2v1 Positively Regulates Protein Aggregation by Modulating Ubiquitin Proteasome System Performance Partially through K63 Ubiquitination. Circ. Res. 2020, 126, 907–922. [Google Scholar] [CrossRef] [PubMed]
- Eddins, M.J.; Carlile, C.M.; Gomez, K.M.; Pickart, C.M.; Wolberger, C. Mms2-Ubc13 covalently bound to ubiquitin reveals the structural basis of linkage-specific polyubiquitin chain formation. Nat. Struct. Mol. Biol. 2006, 13, 915–920. [Google Scholar] [CrossRef]
- Pulvino, M.; Liang, Y.; Oleksyn, D.; DeRan, M.; Van Pelt, E.; Shapiro, J.; Sanz, I.; Chen, L.; Zhao, J. Inhibition of proliferation and survival of diffuse large B-cell lymphoma cells by a small-molecule inhibitor of the ubiquitin-conjugating enzyme Ubc13-Uev1A. Blood 2012, 120, 1668–1677. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.; Fan, Y.H.; Xu, X.; Zhang, H.; Dou, J.; Tang, Y.; Zhong, X.; Rojas, Y.; Yu, Y.; Zhao, Y.; et al. A small-molecule inhibitor of UBE2N induces neuroblastoma cell death via activation of p53 and JNK pathways. Cell Death Dis. 2014, 5, e1079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Golforoush, P.A.; Narasimhan, P.; Chaves-Guerrero, P.P.; Lawrence, E.; Newton, G.; Yan, R.; Harding, S.E.; Perrior, T.; Chapman, K.L.; Schneider, M.D. Selective protection of human cardiomyocytes from anthracycline cardiotoxicity by small molecule inhibitors of MAP4K4. Sci. Rep. 2020, 10, 12060. [Google Scholar] [CrossRef] [PubMed]
- Fiedler, L.R.; Chapman, K.; Xie, M.; Maifoshie, E.; Jenkins, M.; Golforoush, P.A.; Bellahcene, M.; Noseda, M.; Faust, D.; Jarvis, A.; et al. MAP4K4 Inhibition Promotes Survival of Human Stem Cell-Derived Cardiomyocytes and Reduces Infarct Size In Vivo. Cell Stem Cell 2019, 24, 579–591.e512. [Google Scholar] [CrossRef] [Green Version]
- Breckwoldt, K.; Letuffe-Breniere, D.; Mannhardt, I.; Schulze, T.; Ulmer, B.; Werner, T.; Benzin, A.; Klampe, B.; Reinsch, M.C.; Laufer, S.; et al. Differentiation of cardiomyocytes and generation of human engineered heart tissue. Nat. Protoc. 2017, 12, 1177–1197. [Google Scholar] [CrossRef]
- Larhammar, M.; Huntwork-Rodriguez, S.; Rudhard, Y.; Sengupta-Ghosh, A.; Lewcock, J.W. The Ste20 Family Kinases MAP4K4, MINK1, and TNIK Converge to Regulate Stress-Induced JNK Signaling in Neurons. J. Neurosci. 2017, 37, 11074–11084. [Google Scholar] [CrossRef]
- Zohn, I.E.; Li, Y.; Skolnik, E.Y.; Anderson, K.V.; Han, J.; Niswander, L. p38 and a p38-interacting protein are critical for downregulation of E-cadherin during mouse gastrulation. Cell 2006, 125, 957–969. [Google Scholar] [CrossRef] [Green Version]
- Hrdinka, M.; Schlicher, L.; Dai, B.; Pinkas, D.M.; Bufton, J.C.; Picaud, S.; Ward, J.A.; Rogers, C.; Suebsuwong, C.; Nikhar, S.; et al. Small molecule inhibitors reveal an indispensable scaffolding role of RIPK2 in NOD2 signaling. EMBO J. 2018, 37, e99372. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, M.; Fujimoto, Y.; Lucas, P.C.; Nakano, H.; Fukase, K.; Nunez, G.; Inohara, N. A critical role of RICK/RIP2 polyubiquitination in Nod-induced NF-kappaB activation. EMBO J. 2008, 27, 373–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tao, M.; Scacheri, P.C.; Marinis, J.M.; Harhaj, E.W.; Matesic, L.E.; Abbott, D.W. ITCH K63-ubiquitinates the NOD2 binding protein, RIP2, to influence inflammatory signaling pathways. Curr. Biol. 2009, 19, 1255–1263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, S.; Wang, B.; Humphries, F.; Jackson, R.; Healy, M.E.; Bergin, R.; Aviello, G.; Hall, B.; McNamara, D.; Darby, T.; et al. Pellino3 ubiquitinates RIP2 and mediates Nod2-induced signaling and protective effects in colitis. Nat. Immunol. 2013, 14, 927–936. [Google Scholar] [CrossRef]
- Yamada, M.; Ohnishi, J.; Ohkawara, B.; Iemura, S.; Satoh, K.; Hyodo-Miura, J.; Kawachi, K.; Natsume, T.; Shibuya, H. NARF, an nemo-like kinase (NLK)-associated ring finger protein regulates the ubiquitylation and degradation of T cell factor/lymphoid enhancer factor (TCF/LEF). J. Biol. Chem. 2006, 281, 20749–20760. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.; Lu, Y.; Han, D.; Yao, R.; Wang, H.; Zhong, S.; Luo, Y.; Han, R.; Li, K.; Fu, J.; et al. Rnf138 deficiency promotes apoptosis of spermatogonia in juvenile male mice. Cell Death Dis. 2017, 8, e2795. [Google Scholar] [CrossRef] [Green Version]
- Ismail, I.H.; Gagne, J.P.; Genois, M.M.; Strickfaden, H.; McDonald, D.; Xu, Z.; Poirier, G.G.; Masson, J.Y.; Hendzel, M.J. The RNF138 E3 ligase displaces Ku to promote DNA end resection and regulate DNA repair pathway choice. Nat. Cell Biol. 2015, 17, 1446–1457. [Google Scholar] [CrossRef]
- Lv, P.; Li, C.; Wang, M.; Ren, J.; Zhang, Y.; Fu, G. TANK-binding kinase 1 alleviates myocardial ischemia/reperfusion injury through regulating apoptotic pathway. Biochem. Biophys. Res. Commun. 2020, 528, 574–579. [Google Scholar] [CrossRef]
- Deng, K.Q.; Wang, A.; Ji, Y.X.; Zhang, X.J.; Fang, J.; Zhang, Y.; Zhang, P.; Jiang, X.; Gao, L.; Zhu, X.Y.; et al. Suppressor of IKKvarepsilon is an essential negative regulator of pathological cardiac hypertrophy. Nat. Commun. 2016, 7, 11432. [Google Scholar] [CrossRef] [Green Version]
- Zhi, H.; Gong, F.H.; Cheng, W.L.; Zhu, K.; Chen, L.; Yao, Y.; Ye, X.; Zhu, X.Y.; Li, H. Tollip Negatively Regulates Vascular Smooth Muscle Cell-Mediated Neointima Formation by Suppressing Akt-Dependent Signaling. J. Am. Heart Assoc. 2018, 7, e006851. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Jiang, X.L.; Liu, Y.; Jiang, D.S.; Zhang, Y.; Zhang, R.; Chen, Y.; Yang, Q.; Zhang, X.D.; Fan, G.C.; et al. Toll-interacting protein (Tollip) negatively regulates pressure overload-induced ventricular hypertrophy in mice. Cardiovasc. Res. 2014, 101, 87–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wan, N.; Liu, X.; Zhang, X.J.; Zhao, Y.; Hu, G.; Wan, F.; Zhang, R.; Zhu, X.; Xia, H.; Li, H. Toll-interacting protein contributes to mortality following myocardial infarction through promoting inflammation and apoptosis. Br. J. Pharmacol. 2015, 172, 3383–3396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, G.; Ghosh, S. Negative regulation of toll-like receptor-mediated signaling by Tollip. J. Biol. Chem. 2002, 277, 7059–7065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeschke, M.G.; van Baar, M.E.; Choudhry, M.A.; Chung, K.K.; Gibran, N.S.; Logsetty, S. Burn injury. Nat. Rev. Dis. Primers 2020, 6, 11. [Google Scholar] [CrossRef] [PubMed]
- Sood, R.F.; Gibran, N.S.; Arnoldo, B.D.; Gamelli, R.L.; Herndon, D.N.; Tompkins, R.G. Inflammation the Host Response to Injury, I. Early leukocyte gene expression associated with age, burn size, and inhalation injury in severely burned adults. J. Trauma Acute Care Surg. 2016, 80, 250–257. [Google Scholar] [CrossRef] [Green Version]
- Xiao, W.; Mindrinos, M.N.; Seok, J.; Cuschieri, J.; Cuenca, A.G.; Gao, H.; Hayden, D.L.; Hennessy, L.; Moore, E.E.; Minei, J.P.; et al. A genomic storm in critically injured humans. J. Exp. Med. 2011, 208, 2581–2590. [Google Scholar] [CrossRef]
- Finnerty, C.C.; Jeschke, M.G.; Qian, W.J.; Kaushal, A.; Xiao, W.; Liu, T.; Gritsenko, M.A.; Moore, R.J.; Camp, D.G., 2nd; Moldawer, L.L.; et al. Determination of burn patient outcome by large-scale quantitative discovery proteomics. Crit. Care Med. 2013, 41, 1421–1434. [Google Scholar] [CrossRef] [Green Version]
- Seok, J.; Warren, H.S.; Cuenca, A.G.; Mindrinos, M.N.; Baker, H.V.; Xu, W.; Richards, D.R.; McDonald-Smith, G.P.; Gao, H.; Hennessy, L.; et al. Genomic responses in mouse models poorly mimic human inflammatory diseases. Proc. Natl. Acad. Sci. USA 2013, 110, 3507–3512. [Google Scholar] [CrossRef] [Green Version]
- Evers, L.H.; Bhavsar, D.; Mailander, P. The biology of burn injury. Exp. Dermatol. 2010, 19, 777–783. [Google Scholar] [CrossRef]
- Calum, H.; Moser, C.; Jensen, P.O.; Christophersen, L.; Maling, D.S.; van Gennip, M.; Bjarnsholt, T.; Hougen, H.P.; Givskov, M.; Jacobsen, G.K.; et al. Thermal injury induces impaired function in polymorphonuclear neutrophil granulocytes and reduced control of burn wound infection. Clin. Exp. Immunol. 2009, 156, 102–110. [Google Scholar] [CrossRef] [Green Version]
- Nielson, C.B.; Duethman, N.C.; Howard, J.M.; Moncure, M.; Wood, J.G. Burns: Pathophysiology of Systemic Complications and Current Management. J. Burn Care Res. 2017, 38, e469–e481. [Google Scholar] [CrossRef] [PubMed]
- Guillory, A.N.; Clayton, R.P.; Herndon, D.N.; Finnerty, C.C. Cardiovascular Dysfunction Following Burn Injury: What We Have Learned from Rat and Mouse Models. Int. J. Mol. Sci. 2016, 17, 53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Damme, L.; De Waele, J.J. Effect of decompressive laparotomy on organ function in patients with abdominal compartment syndrome: A systematic review and meta-analysis. Crit. Care 2018, 22, 179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boldeanu, L.; Boldeanu, M.V.; Bogdan, M.; Meca, A.D.; Coman, C.G.; Buca, B.R.; Tartau, C.G.; Tartau, L.M. Immunological approaches and therapy in burns (Review). Exp. Ther. Med. 2020, 20, 2361–2367. [Google Scholar] [CrossRef]
- MacConmara, M.P.; Maung, A.A.; Fujimi, S.; McKenna, A.M.; Delisle, A.; Lapchak, P.H.; Rogers, S.; Lederer, J.A.; Mannick, J.A. Increased CD4+ CD25+ T regulatory cell activity in trauma patients depresses protective Th1 immunity. Ann. Surg. 2006, 244, 514–523. [Google Scholar] [CrossRef]
- Schwacha, M.G. Macrophages and post-burn immune dysfunction. Burns 2003, 29, 1–14. [Google Scholar] [CrossRef]
- Vinaik, R.; Abdullahi, A.; Barayan, D.; Jeschke, M.G. NLRP3 inflammasome activity is required for wound healing after burns. Transl. Res. 2020, 217, 47–60. [Google Scholar] [CrossRef]
- Strudwick, X.L.; Cowin, A.J. The role of the inflammatory response in burn injury. In Hot Topics in Burn Injuries; IntechOpen: London, UK, 2018. [Google Scholar]
- Fang, Q.; Guo, S.; Zhou, H.; Han, R.; Wu, P.; Han, C. Astaxanthin protects against early burn-wound progression in rats by attenuating oxidative stress-induced inflammation and mitochondria-related apoptosis. Sci. Rep. 2017, 7, 41440. [Google Scholar] [CrossRef]
- Muszynski, J.A.; Nofziger, R.; Greathouse, K.; Nateri, J.; Hanson-Huber, L.; Steele, L.; Nicol, K.; Groner, J.I.; Besner, G.E.; Raffel, C.; et al. Innate immune function predicts the development of nosocomial infection in critically injured children. Shock 2014, 42, 313–321. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.T.; Friedrich, E.; Heuslein, J.L.; Pferdehirt, R.E.; Dangelo, N.M.; Natesan, S.; Christy, R.J.; Washburn, N.R. Reduction of burn progression with topical delivery of (antitumor necrosis factor-alpha)-hyaluronic acid conjugates. Wound Repair Regen. 2012, 20, 563–572. [Google Scholar] [CrossRef] [Green Version]
- Arslan, E.; Yavuz, M.; Dalay, C. The relationship between tumor necrosis factor (TNF)-alpha and survival following granulocyte-colony stimulating factor (G-CSF) administration in burn sepsis. Burns 2000, 26, 521–524. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Expressions. | 5′-Forward-3′ | 5′-Reverse-3′ | Amplicon Size (bp) | Accession # |
|---|---|---|---|---|
| Casp8 | AATAAAGACAACCCGAGGAACA | ATGCCACAGCCCATCTTCACACTA | 101 | NM_022277.1 |
| Ccl2 | GTCTCAGCCAGATGCAGTTAAT | CTGCTGGTGATTCTCTTGTAGTT | 105 | NM_031530.1 |
| Cd86 | CCGAGTGAGCTCGTAGTATTT | GGTACTTGGCATTCACGTTATC | 100 | NM_020081.1 |
| Cebpb | CTTGATGCAATCCGGATCAAAC | CCCGCAGGAACATCTTTAAGT | 113 | NM_024125.5 |
| Hmgb1 | TCGGCCTTCTTCTTGTTCTG | GTTGTTCCACATCTCTCCTAGTT | 108 | NM_012963.2 |
| Hspa1a | CGTTTGACACTCTGTTGCTTTC | ACGGCCAGGCAAGATTATAC | 89 | NM_031971.2 |
| Ifna | AGAGAGAGAGAGAGAGAGAGAGA | GTGTGATTCCACATTTGCAGTAG | 110 | XM_001076062.4 |
| IL1b | GAGGCCATAGCCCATGATTTA | CTCCTGCTTGACGATCCTTATC | 105 | NM_017019.2 |
| IL10 | AGTGGAGCAGGTGAAGAATG | GAGTGTCACGTAGGCTTCTATG | 109 | NM_012854.2 |
| IL2 | GCAGGCCACAGAATTGAAAC | CCAGCGTCTTCCAAGTGAA | 108 | NM_053836.1 |
| IL6 | GAAGTTAGAGTCACAGAAGGAGTG | GTTTGCCGAGTAGACCTCATAG | 105 | M26744.1 |
| Kcnh8 | GAAGACAGAGCCAAAGGAAGA | CAGGTGTCCTGAGATGTGATAAA | 96 | NM_145095.2 |
| Lta | GCGTCAGTTACCACAGAACA | GCCTCGGGCTTTCTTCTAAA | 100 | NM_080769.2 |
| Ly98 | CCGAAGCGCAAGGAAATTG | TGTGATGGCCCTTAGGAAATAG | 133 | NM_001024279.1 |
| Map4k4 | CTGGTGGAAGTGGTTGGAAA | CATCCTCGGTGACATCCATAAC | 97 | NM_001106904.1 |
| Nr2c2 | CTGATAGCCACTCCCACATTT | GAACTGTACCATCCTCACGTATC | 118 | NM_017323.2 |
| Ptgs2 | GGCCATGGAGTGGACTTAAA | GTCTTTGACTGTGGGAGGATAC | 132 | NM_017232.3 |
| Ripk2 | CCTTTGCCTCCTGTCTTTCT | CCTCTACCCAACCAGCATATT | 100 | NM_001191865.1 |
| Rnf 138 | TTACCAGCCTCTGTTTCACTAC | TGACTTGGCCTTCTCTACATTAC | 83 | NM_053588.3 |
| Tbk1 | GAAGAAGCTGAAGGAGGAGATG | CAGACTCCCGAAGAAGCTAAAG | 137 | NM_001106786.1 |
| Tlr2 | CCAAGAGGAAGCCCAAGAAA | CATGAGGTTCTCCACCCAATAG | 98 | NM_198769.2 |
| Tlr7 | AACCTTTCCCAGAGCATACAG | AGCCTCTGATGGGACAAATAAA | 110 | NM_001097582.1 |
| Tollip | CCCTCCTCTGATGTTGTATGTG | GGTAAGCAAGACAGGAGGTTAG | 108 | NM_001109668.1 |
| Tradd | CTGGACCTTCTGAAACCTAGATG | CTTAGGTCCCACGATGAGAATG | 106 | NM_001100480.1 |
| Position | Refseq | Symbol | Description | Fold Change |
|---|---|---|---|---|
| A01 | NM_001007798 | Btk | Bruton agammaglobulinemia tyrosine kinase | 1.05 |
| A02 | NM_022277 | Casp8 | Caspase 8 | 2.12 |
| A03 | NM_031530 | Ccl2 | Chemokine (C-C motif) ligand 2 | 43.61 |
| A04 | NM_021744 | Cd14 | CD14 molecule | 1.23 |
| A05 | NM_001106405 | Cd180 | CD180 molecule | 1.05 |
| A06 | NM_012926 | Cd80 | Cd80 molecule | 1.05 |
| A07 | NM_020081 | Cd86 | CD86 molecule | 1.08 |
| A08 | NM_024125 | Cebpb | CCAAT/enhancer binding protein (C/EBP), beta | 2.03 |
| A09 | NM_001107588 | Chuk | Conserved helix–loop–helix ubiquitous kinase | 1.55 |
| A10 | NM_001005897 | Clec4e | C-type lectin domain family 4, member e | 1.05 |
| A11 | NM_053852 | Csf2 | Colony stimulating factor 2 (granulocyte-macrophage) | 1.05 |
| A12 | NM_017104 | Csf3 | Colony stimulating factor 3 (granulocyte) | 1.05 |
| B01 | NM_139089 | Cxcl10 | Chemokine (C-X-C motif) ligand 10 | 1.51 |
| B02 | NM_019335 | Eif2ak2 | Eukaryotic translation initiation factor 2-alpha kinase 2 | 2.05 |
| B03 | NM_152937 | Fadd | Fas (TNFRSF6)-associated via death domain | 2.00 |
| B04 | NM_022197 | Fos | FBJ osteosarcoma oncogene | 4.18 |
| B05 | NM_012963 | Hmgb1 | High-mobility group box 1 | 54.92 |
| B06 | NM_001098241 | Hras | Harvey rat sarcoma virus oncogene | 2.16 |
| B07 | NM_031971 | Hspa1a | Heat shock 70 kD protein 1A | 2.47 |
| B08 | NM_022229 | Hspd1 | Heat shock protein 1 (chaperonin) | 4.07 |
| B09 | NM_001014786 | Ifna1 | Interferon-alpha 1 | 1.74 |
| B10 | NM_019127 | Ifnb1 | Interferon beta 1, fibroblast | 1.05 |
| B11 | NM_138880 | Ifng | Interferon gamma | 1.05 |
| B12 | NM_053355 | Ikbkb | Inhibitor of kappa light polypeptide gene enhancer in B-cells, kinase beta | 1.42 |
| C01 | NM_012854 | Il10 | Interleukin 10 | 5.49 |
| C02 | NM_053390 | Il12a | Interleukin 12a | 1.03 |
| C03 | NM_017019 | Il1a | Interleukin 1 alpha | 11,883,604.11 |
| C04 | NM_031512 | Il1b | gb Interleukin 1 beta | 1.47 |
| C05 | NM_013123 | Il1r1 | Interleukin 1 receptor, type I | −1.13 |
| C06 | NM_053836 | Il2 | Interleukin 2 | 1.49 |
| C07 | NM_012589 | Il6 | Interleukin 6 | 3.12 |
| C08 | NM_017020 | Il6r | Interleukin 6 receptor | 1.65 |
| C09 | NM_001127555 | Irak1 | Interleukin-1-receptor-associated kinase 1 | 4.64 |
| C10 | NM_001025422 | Irak2 | Interleukin-1-receptor-associated kinase 2 | 1.06 |
| C11 | NM_012591 | Irf1 | Interferon regulatory factor 1 | 3.11 |
| C12 | NM_001006969 | Irf3 | Interferon regulatory factor 3 | 2.66 |
| D01 | NM_021835 | Jun | Jun oncogene | 1.62 |
| D02 | NM_145095 | Kcnh8 | Potassium voltage-gated channel, subfamily H (eag-related), member 8 | 1.09 |
| D03 | NM_080769 | Lta | Lymphotoxin alpha (TNF superfamily, member 1) | 1.08 |
| D04 | NM_001024279 | Ly96 | Lymphocyte antigen 96 | 1.76 |
| D05 | NM_012798 | Mal | Mal, T-cell differentiation protein | 1.81 |
| D06 | NM_001100674 | Map2k3 | Mitogen-activated protein kinase kinase 3 | −1.43 |
| D07 | NM_001030023 | Map2k4 | Mitogen-activated protein kinase kinase 4 | 2.75 |
| D08 | NM_053887 | Map3k1 | Mitogen-activated protein kinase kinase kinase 1 | 1.08 |
| D09 | NM_001107920 | Map3k7 | Mitogen-activated protein kinase kinase kinase 7 | −1.05 |
| D10 | NM_001106904 | Map4k4 | Mitogen-activated protein kinase kinase kinase kinase 4 | −2.04 |
| D11 | NM_053829 | Mapk8 | Mitogen-activated protein kinase 8 | 2.32 |
| D12 | NM_001100673 | Mapk8ip3 | Mitogen-activated protein kinase 8 interacting protein 3 | 1.74 |
| E01 | NM_017322 | Mapk9 | Mitogen-activated protein kinase 9 | 1.71 |
| E02 | NM_198130 | Myd88 | Myeloid differentiation primary response gene 88 | 2.10 |
| E03 | NM_001276711 | Nfkb1 | Nuclear factor of kappa light polypeptide gene enhancer in B-cells 1 | 5.52 |
| E04 | NM_001008349 | Nfkb2 | Nuclear factor of kappa light polypeptide gene enhancer in B-cells 2, p49/p100 | 1.25 |
| E05 | NM_001105720 | Nfkbia | Nuclear factor of kappa light polypeptide gene enhancer in B-cells inhibitor, alpha | 2.12 |
| E06 | NM_030867 | Nfkbib | Nuclear factor of kappa light polypeptide gene enhancer in B-cells inhibitor, beta | 1.54 |
| E07 | NM_212509 | Nfkbil1 | Nuclear factor of kappa light polypeptide gene enhancer in B-cells inhibitor-like 1 | 3.40 |
| E08 | NM_001108133 | Nfrkb | Nuclear factor related to kappaB binding protein | −1.19 |
| E09 | NM_017323 | Nr2c2 | Nuclear receptor subfamily 2, group C, member 2 | 1.00 |
| E10 | NM_001100565 | Peli1 | Pellino 1 | 4.12 |
| E11 | NM_053373 | Pglyrp1 | Peptidoglycan recognition protein 1 | 2.17 |
| E12 | NM_013196 | Ppara | Peroxisome proliferator activated receptor alpha | −1.20 |
| F01 | NM_017232 | Ptgs2 | Prostaglandin-endoperoxide synthase 2 | 1.05 |
| F02 | XM_006221869 | Rel | V-rel avian reticuloendotheliosis viral oncogene homolog | 1.07 |
| F03 | NM_199267 | Rela | V-rel reticuloendotheliosis viral oncogene homolog A (avian) | 1.96 |
| F04 | NM_001191865 | Ripk2 | Receptor-interacting serine-threonine kinase 2 | −1.63 |
| F05 | NM_053588 | Rnf138 | Ring finger protein 138 | −1.53 |
| F06 | NM_001105817 | Sarm1 | Sterile alpha and TIR motif containing 1 | 3.57 |
| F07 | NM_001106786 | Tbk1 | TANK-binding kinase 1 | −2.36 |
| F08 | NM_001108890 | Ticam2 | Toll-like receptor adaptor molecule 2 | 11,993,941.01 |
| F09 | NM_001172120 | Tlr1 | Toll-like receptor 1 | 1.05 |
| F10 | NM_198769 | Tlr2 | Toll-like receptor 2 | 1.87 |
| F11 | NM_198791 | Tlr3 | Toll-like receptor 3 | 1.05 |
| F12 | NM_019178 | Tlr4 | Toll-like receptor 4 | 2.03 |
| G01 | NM_001145828 | Tlr5 | Toll-like receptor 5 | 1.05 |
| G02 | NM_207604 | Tlr6 | Toll-like receptor 6 | 1.48 |
| G03 | NM_001097582 | Tlr7 | Toll-like receptor 7 | 1.96 |
| G04 | NM_198131 | Tlr9 | Toll-like receptor 9 | 1.61 |
| G05 | NM_012675 | Tnf | Tumor necrosis factor (TNF superfamily, member 2) | 1.05 |
| G06 | NM_013091 | Tnfrsf1a | Tumor necrosis factor receptor superfamily, member 1a | −1.15 |
| G07 | NM_001024771 | Tnip2 | TNFAIP3 interacting protein 2 | 2.10 |
| G08 | NM_001109668 | Tollip | Toll interacting protein | −2.29 |
| G09 | NM_001100480 | Tradd | TNFRSF1A-associated via death domain | 2.02 |
| G10 | NM_001107754 | Traf6 | Tnf-receptor-associated factor 6 | −1.09 |
| G11 | NM_053928 | Ube2n | Ubiquitin-conjugating enzyme E2N (UBC13 homolog, yeast) | 6.17 |
| G12 | NM_001110345 | Ube2v1 | Ubiquitin-conjugating enzyme E2 variant 1 | 5.39 |
| Position | Symbol | Description | Fold Regulation (24 hpb vs. Sham) | |
|---|---|---|---|---|
| Toll-like receptor | ||||
| F10 | Tlr2 | Toll-like receptor 2 | 1.87 | |
| F12 | Tlr4 | Toll-like receptor 4 | 2.03 | |
| G03 | Tlr7 | Toll-like receptor 7 | 1.96 | |
| G04 | Tlr9 | Toll-like receptor 9 | 1.61 | |
| Effectors | ||||
| C09 | Irak1 | Interleukin-1 receptor-associated kinase 1 | 4.64 | |
| Interacting proteins and adaptors | ||||
| B07 | Hspa1a | Heat shock 70 kD protein 1A | 2.47 | |
| D12 | Mapk8ip3 | Mitogen-activated protein kinase 8 interacting protein 3 | 1.74 | |
| G08 | Tollip | Toll interacting protein | −2.29 | |
| Apoptosis | ||||
| A02 | Casp8 | Caspase 8 | 2.12 | |
| Ubiquitin-conjugating pathway | ||||
| E10 | Peli1 | Pellino 1 | 4.12 | |
| G11 | Ube2n | Ubiquitin-conjugating enzyme E2N (UBC13 homolog, yeast) | 6.17 | |
| G12 | Ube2v1 | Ubiquitin-conjugating enzyme E2 variant 1 | 5.39 | |
| A03 | Ccl2 | Chemokine (C-C motif) ligand 2 | 43.61 | |
| A08 | Cebpb | CCAAT/enhancer binding protein (C/EBP), beta | 2.03 | |
| A09 | Chuk | Conserved helix–loop–helix ubiquitous kinase | 1.55 | |
| B02 | Eif2ak2 | Eukaryotic translation initiation factor 2-alpha kinase 2 | 2.05 | |
| B04 | Fos | FBJ osteosarcoma oncogene | 4.18 | |
| B05 | Hmgb1 | High-mobility group box 1 | 54.92 | |
| B08 | Hspd1 | Heat shock protein 1 (chaperonin) | 4.07 | |
| Regulation of adaptive immunity | ||||
| C01 | Il10 | Interleukin 10 | 5.49 | |
| E03 | Nfkb1 | Nuclear factor of kappa light polypeptide gene enhancer in B-cells 1 | 5.52 | |
| E06 | Nfkbib | Nuclear factor of kappa light polypeptide gene enhancer in B-cells inhibitor, beta | 1.54 | |
| Downstream pathway of Toll-like receptors NFKB signaling | ||||
| C01 | Il10 | Interleukin 10 | 5.49 | |
| C09 | Irak1 | Interleukin-1-receptor-associated kinase 1 | 4.64 | |
| D10 | Map4k4 | Mitogen-activated protein kinase kinase kinase kinase 4 | −2.04 | |
| E03 | Nfkb1 | Nuclear factor of kappa light polypeptide gene enhancer in B-cells 1 | 5.52 | |
| E06 | Nfkbib | Nuclear factor of kappa light polypeptide gene enhancer in B-cells inhibitor, beta | 1.54 | |
| JNK/p38 signaling | ||||
| D12 | Mapk8ip3 | Mitogen-activated protein kinase 8 interacting protein 3 | 1.74 | |
| JAK/STAT signaling | ||||
| A03 | Ccl2 | Chemokine (C-C motif) ligand 2 | 43.61 | |
| F07 | Tbk1 | TANK-binding kinase 1 | −2.36 | |
| Interferon regulatory factor signaling | ||||
| B01 | Cxcl10 | Chemokine (C-X-C motif) ligand 10 | 1.51 | |
| C11 | Irf1 | Interferon regulatory factor 1 | 3.11 | |
| F07 | Tbk1 | TANK-binding kinase 1 | −2.36 | |
| Cytokine signaling | ||||
| A03 | Ccl2 | Chemokine (C-C motif) ligand 2 | 43.61 | |
| C09 | Irak1 | Interleukin-1-receptor-associated kinase 1 | 4.64 | |
| NFkB/IL6 pathway | ||||
| B09 | Ifna1 | Interferon-alpha 1 | 1.74 | |
| C03 | Il1a | Interleukin 1 alpha | 11,883,604.11 | |
| C07 | Il6 | Interleukin 6 | 3.12 | |
| JUN-MARK signaling | ||||
| D01 | Jun | Jun oncogene | 1.62 | |
| D04 | Ly96 | Lymphocyte antigen 96 | 1.76 | |
| D11 | Mapk8 | Mitogen-activated protein kinase 8 | 2.32 | |
| E01 | Mapk9 | Mitogen-activated protein kinase 9 | 1.71 | |
| Others | ||||
| E11 | Pglyrp1 | Peptidoglycan recognition protein 1 | 2.17 | |
| F04 | Ripk2 | Receptor-interacting serine-threonine kinase 2 | −1.63 | |
| F05 | Rnf138 | Ring finger protein 138 | −1.53 | |
| F06 | Sarm1 | Sterile alpha and TIR motif containing 1 | 3.57 | |
| F08 | Ticam2 | Toll-like receptor adaptor molecule 2 | 11,993,941.01 | |
| G09 | Tradd | TNFRSF1A-associated via death domain | 2.02 |
| Sub Ontologies | ID | Description | Observed Gene Count | Bachground Gene Count | Strength |
|---|---|---|---|---|---|
| GO-MF | GO:0035663 | Toll-like receptor 2 binding | 2 | 3 | 2.41 |
| GO-MF | GO:0071723 | Lipopeptide binding | 3 | 5 | 2.36 |
| GO-MF | GO:0035325 | Toll-like receptor binding | 4 | 14 | 2.04 |
| GO-MF | GO:0050135 | NAD(P)+ nucleosidase activity | 5 | 18 | 2.03 |
| GO-MF | GO:0061809 | NAD+ nucleotidase, cyclic ADP-ribose generating | 5 | 18 | 2.03 |
| GO-MF | GO:0005132 | Type i interferon receptor binding | 2 | 10 | 1.89 |
| GO-MF | GO:0038187 | Pattern recognition receptor activity | 4 | 21 | 1.87 |
| GO-MF | GO:0005149 | interleukin-1 receptor binding | 3 | 17 | 1.83 |
| GO-MF | GO:0005164 | Tumor necrosis factor receptor binding | 5 | 34 | 1.75 |
| GO-MF | GO:0035370 | UBC13-UEV1A complex | 2 | 4 | 2.29 |
| GO-CC | GO:0033256 | I-kappaB/NF-kappaB complex | 3 | 9 | 2.11 |
| GO-CC | GO:0046696 | Lipopolysaccharide receptor complex | 2 | 8 | 1.98 |
| GO-CC | GO:0045335 | Phagocytic vesicle | 4 | 135 | 1.06 |
| GO-CC | GO:0042025 | Host cell nucleus | 5 | 187 | 1.01 |
| GO-CC | GO:0030139 | Endocytic vesicle | 5 | 218 | 0.95 |
| GO-CC | GO:0045121 | Membrane raft | 10 | 446 | 0.94 |
| GO-CC | GO:0043235 | Receptor complex | 9 | 482 | 0.86 |
| GO-CC | GO:0009897 | External side of plasma membrane | 8 | 512 | 0.78 |
| GO-CC | GO:0098552 | Side of membrane | 9 | 730 | 0.68 |
| GO-CC | GO:0005615 | Extracellular space | 21 | 1776 | 0.66 |
| GO-BP | GO:0002874 | Regulation of chronic inflammatory response to antigenic stimulus | 3 | 3 | 2.59 |
| GO-BP | GO:0060559 | Positive regulation of calcidiol 1-monooxygenase activity | 3 | 3 | 2.59 |
| GO-BP | GO:0002876 | Positive regulation of chronic inflammatory response to antigenic stimulus | 2 | 2 | 2.59 |
| GO-BP | GO:0070340 | Detection of bacterial lipopeptide | 2 | 2 | 2.59 |
| GO-BP | GO:0071727 | Cellular response to triacyl bacterial lipopeptide | 2 | 2 | 2.59 |
| GO-BP | GO:0035711 | T-helper 1 cell activation | 3 | 4 | 2.46 |
| GO-BP | GO:0001781 | Neutrophil apoptotic process | 2 | 3 | 2.41 |
| GO-BP | GO:0060558 | Regulation of calcidiol 1-monooxygenase activity | 4 | 8 | 2.29 |
| GO-BP | GO:0071221 | Cellular response to bacterial lipopeptide | 4 | 8 | 2.29 |
| GO-BP | GO:0071726 | Cellular response to diacyl bacterial lipopeptide | 3 | 6 | 2.29 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wen, J.J.; Mobli, K.; Radhakrishnan, G.L.; Radhakrishnan, R.S. Regulation of Key Immune-Related Genes in the Heart Following Burn Injury. J. Pers. Med. 2022, 12, 1007. https://doi.org/10.3390/jpm12061007
Wen JJ, Mobli K, Radhakrishnan GL, Radhakrishnan RS. Regulation of Key Immune-Related Genes in the Heart Following Burn Injury. Journal of Personalized Medicine. 2022; 12(6):1007. https://doi.org/10.3390/jpm12061007
Chicago/Turabian StyleWen, Jake J., Keyan Mobli, Geetha L. Radhakrishnan, and Ravi S. Radhakrishnan. 2022. "Regulation of Key Immune-Related Genes in the Heart Following Burn Injury" Journal of Personalized Medicine 12, no. 6: 1007. https://doi.org/10.3390/jpm12061007
APA StyleWen, J. J., Mobli, K., Radhakrishnan, G. L., & Radhakrishnan, R. S. (2022). Regulation of Key Immune-Related Genes in the Heart Following Burn Injury. Journal of Personalized Medicine, 12(6), 1007. https://doi.org/10.3390/jpm12061007