
The Impact of Air Pollution Exposure on the MicroRNA Machinery and Lung Cancer Development




Abstract
1. Introduction to miRNA
1.1. Basic Information on miRNA
1.2. Methodological Approaches for miRNA Investigation
1.3. miRNA as a Biomarker
2. Lung Cancer and miRNA
3. Air Pollution and miRNA
3.1. The Effect of Air Pollution on miRNA Expression in Healthy Adults
3.2. The Effect of Air Pollution on miRNA Expression in Children
3.3. The Effect of Air Pollution on miRNA Expression in Elderly Subjects
3.4. The Effect of Air Pollution on miRNA Expression in Overweight/Obese Subjects
3.5. The Effect of Occupational Exposure to Polluted Air on miRNA Expression
3.6. The Effect of Tobacco Smoking on miRNA Expression
3.7. miRNA Expression in Subjects Exposed to Other Sources of Air Pollution
3.8. miRNAs Commonly Deregulated by Various Types of Air Pollutants
4. miRNA Affected by Air Pollution Exposure and Implicated in Lung Cancer
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| CLL | Chronic lymphocytic leukemia |
| COPD | Chronic obstructive pulmonary disease |
| ddPCR | Droplet digital PCR |
| DNA | Deoxyribonucleic acid |
| EC | Elemental carbon |
| IHD | Ischemic heart disease |
| LC | Lung cancer |
| LPG | Liquid petroleum gas |
| miRNA | MicroRNA molecule |
| NGS | Next generation sequencing |
| NSCLC | Non-small cell lung cancer |
| PAH | Polycyclic aromatic hydrocarbon |
| PM | Particle matters |
| pre-miRNA | Precursor miRNA |
| pri-miRNA | Primary miRNA |
| qRT-PCR | Quantitative real-time polymerase chain reaction |
| RISC | RNA-induced silencing complex |
| RNA | Ribonucleic acid |
| SCLC | Small cell lung cancer |
| VOC | Volatile organic compound |
| WHO | World Health Organization |
References
- Lee, R.C.; Feinbaum, R.L.; Ambros, V.; The, C. Elegans Heterochronic Gene Lin-4 Encodes Small RNAs with Antisense Complementarity to Lin-14. Cell 1993, 75, 843–854. [Google Scholar] [CrossRef]
- miRbase. Available online: http://mirbase.org/ (accessed on 1 December 2020).
- Kozomara, A.; Birgaoanu, M.; Griffiths-Jones, S. MiRBase: From MicroRNA Sequences to Function. Nucleic Acids Res. 2019, 47, D155–D162. [Google Scholar] [CrossRef] [PubMed]
- Alles, J.; Fehlmann, T.; Fischer, U.; Backes, C.; Galata, V.; Minet, M.; Hart, M.; Abu-Halima, M.; Grässer, F.A.; Lenhof, H.-P.; et al. An Estimate of the Total Number of True Human MiRNAs. Nucleic Acids Res. 2019, 47, 3353–3364. [Google Scholar] [CrossRef] [PubMed]
- Tomari, Y.; Zamore, P.D. MicroRNA Biogenesis: Drosha Can’t Cut It without a Partner. Curr. Biol. 2005, 15, R61–R64. [Google Scholar] [CrossRef]
- Michlewski, G.; Cáceres, J.F. Post-Transcriptional Control of MiRNA Biogenesis. RNA 2019, 25, 1–16. [Google Scholar] [CrossRef]
- Bernardo, B.C.; Charchar, F.J.; Lin, R.C.Y.; McMullen, J.R. A MicroRNA Guide for Clinicians and Basic Scientists: Background and Experimental Techniques. Heart Lung Circ. 2012, 21, 131–142. [Google Scholar] [CrossRef]
- Tian, T.; Wang, J.; Zhou, X. A Review: MicroRNA Detection Methods. Org. Biomol. Chem. 2015, 13, 2226–2238. [Google Scholar] [CrossRef]
- Bartošík, M.; Jiráková, L. Current Methods of MicroRNA Analysis. Klin. Onkol. 2018, 31. [Google Scholar] [CrossRef]
- Krepelkova, I.; Mrackova, T.; Izakova, J.; Dvorakova, B.; Chalupova, L.; Mikulik, R.; Slaby, O.; Bartos, M.; Ruzicka, V. Evaluation of MiRNA Detection Methods for the Analytical Characteristic Necessary for Clinical Utilization. Biotechniques 2019, 66, 277–284. [Google Scholar] [CrossRef]
- Tajadini, M.; Panjehpour, M.; Javanmard, S. Comparison of SYBR Green and TaqMan Methods in Quantitative Real-Time Polymerase Chain Reaction Analysis of Four Adenosine Receptor Subtypes. Adv. Biomed. Res. 2014, 3, 85. [Google Scholar] [CrossRef]
- González-Bermúdez, L.; Anglada, T.; Genescà, A.; Martín, M.; Terradas, M. Identification of Reference Genes for RT-QPCR Data Normalisation in Aging Studies. Sci. Rep. 2019, 9, 13970. [Google Scholar] [CrossRef] [PubMed]
- Backes, C.; Meese, E.; Keller, A. Specific MiRNA Disease Biomarkers in Blood, Serum and Plasma: Challenges and Prospects. Mol. Diagn. Ther. 2016, 20, 509–518. [Google Scholar] [CrossRef] [PubMed]
- Vishnoi, A.; Rani, S. MiRNA Biogenesis and Regulation of Diseases: An Overview. In MicroRNA Profiling; Rani, S., Ed.; Methods in Molecular Biology; Springer: New York, NY, USA, 2017; Volume 1509, pp. 1–10. ISBN 978-1-4939-6522-9. [Google Scholar]
- Wojciechowska, A.; Osiak, A.; Kozar-Kamińska, K. MicroRNA in Cardiovascular Biology and Disease. Adv. Clin. Exp. Med. 2017, 26, 868–874. [Google Scholar] [CrossRef] [PubMed]
- Calin, G.A.; Dumitru, C.D.; Shimizu, M.; Bichi, R.; Zupo, S.; Noch, E.; Aldler, H.; Rattan, S.; Keating, M.; Rai, K.; et al. Frequent Deletions and Down-Regulation of Micro-RNA Genes MiR15 and MiR16 at 13q14 in Chronic Lymphocytic Leukemia. Proc. Natl. Acad. Sci. USA 2002, 99, 15524–15529. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Available online: https://www.who.int/ (accessed on 1 December 2020).
- Pogribny, I.P. MicroRNA Dysregulation during Chemical Carcinogenesis. Epigenomics 2009, 1, 281–290. [Google Scholar] [CrossRef]
- Chen, T. The Role of MicroRNA in Chemical Carcinogenesis. J. Environ. Sci. Health Part C 2010, 28, 89–124. [Google Scholar] [CrossRef]
- Izzotti, A.; Pulliero, A. The Effects of Environmental Chemical Carcinogens on the MicroRNA Machinery. Int. J. Hyg. Environ. Health 2014, 217, 601–627. [Google Scholar] [CrossRef]
- Ceccaroli, C.; Pulliero, A.; Geretto, M.; Izzotti, A. Molecular Fingerprints of Environmental Carcinogens in Human Cancer. J. Environ. Sci. Health C Environ. Carcinog. Ecotoxicol. Rev. 2015, 33, 188–228. [Google Scholar] [CrossRef]
- Pogribny, I.P.; Beland, F.A.; Rusyn, I. The Role of MicroRNAs in the Development and Progression of Chemical-Associated Cancers. Toxicol. Appl. Pharmacol. 2016, 312, 3–10. [Google Scholar] [CrossRef]
- Li, M.; Huo, X.; Davuljigari, C.B.; Dai, Q.; Xu, X. MicroRNAs and Their Role in Environmental Chemical Carcinogenesis. Environ. Geochem. Health 2019, 41, 225–247. [Google Scholar] [CrossRef]
- Rzeszutek, I.; Singh, A. Small RNAs, Big Diseases. Int. J. Mol. Sci. 2020, 21, 5699. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Croce, C.M. The Role of MicroRNAs in Human Cancer. Signal Transduct. Target. Ther. 2016, 1, 15004. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Pan, X.; Cobb, G.P.; Anderson, T.A. MicroRNAs as Oncogenes and Tumor Suppressors. Dev. Biol. 2007, 302, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Celic, T.; Meuth, V.; Six, I.; Massy, Z.; Metzinger, L. The Mir-221/222 Cluster Is a Key Player in Vascular Biology via the Fine-Tuning of Endothelial Cell Physiology. CVP 2016, 15, 40–46. [Google Scholar] [CrossRef]
- Takamizawa, J.; Konishi, H.; Yanagisawa, K.; Tomida, S.; Osada, H.; Endoh, H.; Harano, T.; Yatabe, Y.; Nagino, M.; Nimura, Y.; et al. Reduced Expression of the Let-7 MicroRNAs in Human Lung Cancers in Association with Shortened Postoperative Survival. Cancer Res. 2004, 64, 3753–3756. [Google Scholar] [CrossRef] [PubMed]
- Travis, W.D.; Brambilla, E.; Burke, A.P.; Marx, A.; Nicholson, A.G. Introduction to The 2015 World Health Organization Classification of Tumors of the Lung, Pleura, Thymus, and Heart. J. Thorac. Oncol. 2015, 10, 1240–1242. [Google Scholar] [CrossRef]
- Noone, A.-M.; Cronin, K.A.; Altekruse, S.F.; Howlader, N.; Lewis, D.R.; Petkov, V.I.; Penberthy, L. Cancer Incidence and Survival Trends by Subtype Using Data from the Surveillance Epidemiology and End Results Program, 1992–2013. Cancer Epidemiol. Biomark. Prev. 2017, 26, 632–641. [Google Scholar] [CrossRef]
- Feng, B.; Zhang, K.; Wang, R.; Chen, L. Non-Small-Cell Lung Cancer and MiRNAs: Novel Biomarkers and Promising Tools for Treatment. Clin. Sci. 2015, 128, 619–634. [Google Scholar] [CrossRef]
- Patnaik, S.K.; Kannisto, E.D.; Mallick, R.; Vachani, A.; Yendamuri, S. Whole Blood MicroRNA Expression May Not Be Useful for Screening Non-Small Cell Lung Cancer. PLoS ONE 2017, 12, e0181926. [Google Scholar] [CrossRef]
- He, Q.; Fang, Y.; Lu, F.; Pan, J.; Wang, L.; Gong, W.; Fei, F.; Cui, J.; Zhong, J.; Hu, R.; et al. Analysis of Differential Expression Profile of MiRNA in Peripheral Blood of Patients with Lung Cancer. J. Clin. Lab. Anal. 2019, 33, e23003. [Google Scholar] [CrossRef]
- Singh, A.; Kant, R.; Saluja, T.S.; Tripathi, T.; Srivastava, K.; Naithani, M.; Gupta, A.; Mirza, A.A.; Prakash, V.; Singh, S.K. Differential Diagnosis of Non-Small Cell Lung Carcinoma by Circulating MicroRNA. J. Cancer Res. Ther. 2020, 16, 127–131. [Google Scholar] [CrossRef]
- Wozniak, M.B.; Scelo, G.; Muller, D.C.; Mukeria, A.; Zaridze, D.; Brennan, P. Circulating MicroRNAs as Non-Invasive Biomarkers for Early Detection of Non-Small-Cell Lung Cancer. PLoS ONE 2015, 10, e0125026. [Google Scholar] [CrossRef] [PubMed]
- Tai, M.C.; Yanagisawa, K.; Nakatochi, M.; Hotta, N.; Hosono, Y.; Kawaguchi, K.; Naito, M.; Taniguchi, H.; Wakai, K.; Yokoi, K.; et al. Blood-Borne MiRNA Profile-Based Diagnostic Classifier for Lung Adenocarcinoma. Sci. Rep. 2016, 6, 31389. [Google Scholar] [CrossRef] [PubMed]
- Feng, H.; Ge, F.; Du, L.; Zhang, Z.; Liu, D. MiR-34b-3p Represses Cell Proliferation, Cell Cycle Progression and Cell Apoptosis in Non-Small-Cell Lung Cancer (NSCLC) by Targeting CDK4. J. Cell. Mol. Med. 2019, 23, 5282–5291. [Google Scholar] [CrossRef] [PubMed]
- Olbromski, M.; Grzegrzolka, J.; Jankowska-Konsur, A.; Witkiewicz, W.; Podhorska-Okolow, M.; Dziegiel, P. MicroRNAs Modulate the Expression of the SOX18 Transcript in Lung Squamous Cell Carcinoma. Oncol. Rep. 2016, 36, 2884–2892. [Google Scholar] [CrossRef]
- Olbromski, M.; Rzechonek, A.; Grzegrzolka, J.; Glatzel-Plucinska, N.; Chachaj, A.; Werynska, B.; Podhorska-Okolow, M.; Dziegiel, P. Influence of MiR-7a and MiR-24-3p on the SOX18 Transcript in Lung Adenocarcinoma. Oncol. Rep. 2018, 39, 201–208. [Google Scholar] [CrossRef] [PubMed]
- D’Antona, P.; Cattoni, M.; Dominioni, L.; Poli, A.; Moretti, F.; Cinquetti, R.; Gini, E.; Daffrè, E.; Noonan, D.M.; Imperatori, A.; et al. Serum MiR-223: A Validated Biomarker for Detection of Early-Stage Non-Small Cell Lung Cancer. Cancer Epidemiol. Biomark. Prev. 2019, 28, 1926–1933. [Google Scholar] [CrossRef] [PubMed]
- Wen, Z.N.; Ling, Z.G.; Huang, Y.; Li, X. Expression and differential diagnostic value of serum microRNA for invasive pulmonary aspergillosis. Zhonghua Jie He He Hu Xi Za Zhi 2017, 40, 272–277. [Google Scholar] [CrossRef]
- Feng, Y.; Yang, C.; Hu, D.; Wang, X.; Liu, X. MiR-675 Promotes Disease Progression of Non-Small Cell Lung Cancer via Activating NF-ΚB Signaling Pathway. Cell. Mol. Biol. 2017, 63, 7–10. [Google Scholar] [CrossRef]
- Liang, Y.-Y.; Huang, J.-C.; Tang, R.-X.; Chen, W.-J.; Chen, P.; Cen, W.-L.; Shi, K.; Gao, L.; Gao, X.; Liu, A.-G.; et al. Clinical Value of MiR-198-5p in Lung Squamous Cell Carcinoma Assessed Using Microarray and RT-QPCR. World J. Surg. Oncol. 2018, 16, 22. [Google Scholar] [CrossRef]
- Liu, M.; Zeng, X.; Lu, Y.-X.; Mo, Y.-J.; Liao, T.-H.; Gan, C.; Lu, X.-Q. Study on Molecular Mechanism of MiRNA-29a in Promoting Proliferation and Invasion of Non-Small-Cell Lung Cancer by Inhibiting MTSS1. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 5531–5538. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Yu, L.; Wang, C.-M.; Zhou, X.-F. MiR-1275 Promotes Non-Small Cell Lung Cancer Cell Proliferation and Metastasis by Regulating LZTS3 Expression. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 2680–2687. [Google Scholar] [CrossRef] [PubMed]
- Maemura, K.; Watanabe, K.; Ando, T.; Hiyama, N.; Sakatani, T.; Amano, Y.; Kage, H.; Nakajima, J.; Yatomi, Y.; Nagase, T.; et al. Altered Editing Level of MicroRNAs Is a Potential Biomarker in Lung Adenocarcinoma. Cancer Sci. 2018, 109, 3326–3335. [Google Scholar] [CrossRef] [PubMed]
- Pak, M.G.; Lee, C.-H.; Lee, W.-J.; Shin, D.-H.; Roh, M.-S. Unique MicroRNAs in Lung Adenocarcinoma Groups According to Major TKI Sensitive EGFR Mutation Status. Diagn. Pathol. 2015, 10, 99. [Google Scholar] [CrossRef] [PubMed]
- Zhu, K.; Ding, H.; Wang, W.; Liao, Z.; Fu, Z.; Hong, Y.; Zhou, Y.; Zhang, C.-Y.; Chen, X. Tumor-Suppressive MiR-218-5p Inhibits Cancer Cell Proliferation and Migration via EGFR in Non-Small Cell Lung Cancer. Oncotarget 2016, 7, 28075–28085. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Hu, C.; Lian, L.; Hui, K.; Wang, L.; Qiao, Y.; Liu, L.; Liang, L.; Jiang, X. MiR-497 Suppresses Malignant Phenotype in Non-small Cell Lung Cancer via Targeting KDR. Oncol. Rep. 2019, 42, 443–452. [Google Scholar] [CrossRef]
- Goto, A.; Tanaka, M.; Yoshida, M.; Umakoshi, M.; Nanjo, H.; Shiraishi, K.; Saito, M.; Kohno, T.; Kuriyama, S.; Konno, H.; et al. The Low Expression of MiR-451 Predicts a Worse Prognosis in Non-Small Cell Lung Cancer Cases. PLoS ONE 2017, 12, e0181270. [Google Scholar] [CrossRef]
- Sun, X.; Xu, M.; Liu, H.; Ming, K. MicroRNA-219 Is Downregulated in Non-Small Cell Lung Cancer and Inhibits Cell Growth and Metastasis by Targeting HMGA2. Mol. Med. Rep. 2017, 16, 3557–3564. [Google Scholar] [CrossRef]
- Ye, M.-F.; Zhang, J.-G.; Guo, T.-X.; Pan, X.-J. MiR-504 Inhibits Cell Proliferation and Invasion by Targeting LOXL2 in Non Small Cell Lung Cancer. Biomed. Pharmacother. 2018, 97, 1289–1295. [Google Scholar] [CrossRef]
- Liu, S.; Ge, X.; Su, L.; Zhang, A.; Mou, X. MicroRNA-454 Inhibits Non-small Cell Lung Cancer Cells Growth and Metastasis via Targeting Signal Transducer and Activator of Transcription-3. Mol. Med. Rep. 2018, 17, 3979–3986. [Google Scholar] [CrossRef]
- Wu, Z.; Yuan, Q.; Yang, C.; Zhang, X.; Qi, P.; Huang, H.; Ma, Z. Downregulation of Oncogenic Gene TGFβR2 by MiRNA-107 Suppresses Non-Small Cell Lung Cancer. Pathol. Res. Pract. 2020, 216, 152690. [Google Scholar] [CrossRef] [PubMed]
- Qian, L.; Lin, L.; Du, Y.; Hao, X.; Zhao, Y.; Liu, X. MicroRNA-588 Suppresses Tumor Cell Migration and Invasion by Targeting GRN in Lung Squamous Cell Carcinoma. Mol. Med. Rep. 2016, 14, 3021–3028. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Wang, J.-J.; Li, J.-S.; Xu, Q.-Y. MiR-103 Functions as a Tumor Suppressor by Directly Targeting Programmed Cell Death 10 in NSCLC. Oncol. Res. 2018, 26, 519–528. [Google Scholar] [CrossRef]
- Pirooz, H.J.; Jafari, N.; Rastegari, M.; Fathi-Roudsari, M.; Tasharrofi, N.; Shokri, G.; Tamadon, M.; Sazegar, H.; Kouhkan, F. Functional SNP in MicroRNA-491-5p Binding Site of MMP9 3′-UTR Affects Cancer Susceptibility. J. Cell. Biochem. 2018, 119, 5126–5134. [Google Scholar] [CrossRef] [PubMed]
- Flamini, V.; Jiang, W.G.; Cui, Y. Therapeutic Role of MiR-140-5p for the Treatment of Non-Small Cell Lung Cancer. Anticancer Res. 2017, 37, 4319–4327. [Google Scholar] [CrossRef]
- Jeon, S.H.; Yoo, J.K.; Kim, C.M.; Lim, E.S.; Lee, S.J.; Lee, J.M.; Oh, S.-H.; Kim, J.K. The Novel Hsa-MiR-12528 Regulates Tumourigenesis and Metastasis through Hypo-Phosphorylation of AKT Cascade by Targeting IGF-1R in Human Lung Cancer. Cell Death Dis. 2018, 9, 493. [Google Scholar] [CrossRef]
- Xu, L.; Xu, X.; Huang, H.; Ma, Z.; Zhang, S.; Niu, P.; Chen, Y.; Ping, J.; Lu, P.; Yu, C.; et al. MiR-1260b Promotes the Migration and Invasion in Non-Small Cell Lung Cancer via Targeting PTPRK. Pathol. Res. Pract. 2018, 214, 776–783. [Google Scholar] [CrossRef]
- Fujiwara, Y.; Saito, M.; Robles, A.I.; Nishida, M.; Takeshita, F.; Watanabe, M.; Ochiya, T.; Yokota, J.; Kohno, T.; Harris, C.C.; et al. A Nucleolar Stress-Specific P53-MiR-101 Molecular Circuit Functions as an Intrinsic Tumor-Suppressor Network. EBioMedicine 2018, 33, 33–48. [Google Scholar] [CrossRef]
- Liang, G.; Meng, W.; Huang, X.; Zhu, W.; Yin, C.; Wang, C.; Fassan, M.; Yu, Y.; Kudo, M.; Xiao, S.; et al. MiR-196b-5p-Mediated Downregulation of TSPAN12 and GATA6 Promotes Tumor Progression in Non-Small Cell Lung Cancer. Proc. Natl. Acad. Sci. USA 2020, 117, 4347–4357. [Google Scholar] [CrossRef]
- Wang, H.X.; Wang, X.Y.; Fei, J.W.; Li, F.H.; Han, J.; Qin, X. MicroRNA-23B Inhibits Non-Small Cell Lung Cancer Proliferation, Invasion and Migration via Downregulation of RUNX2 and Inhibition of Wnt/Β-Catenin Signaling Pathway. J. Biol. Regul. Homeost. Agents 2020, 34, 825–835. [Google Scholar] [CrossRef]
- Ying, L.; Du, L.; Zou, R.; Shi, L.; Zhang, N.; Jin, J.; Xu, C.; Zhang, F.; Zhu, C.; Wu, J.; et al. Development of a Serum MiRNA Panel for Detection of Early Stage Non-Small Cell Lung Cancer. Proc. Natl. Acad. Sci. USA 2020, 117, 25036–25042. [Google Scholar] [CrossRef]
- Li, J.; Fang, H.; Jiang, F.; Ning, Y. External Validation of a Panel of Plasma MicroRNA Biomarkers for Lung Cancer. Biomark. Med. 2019, 13, 1557–1564. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.-L.; Zhang, Z.-L.; Zhu, X.-B.; Xu, L.; Lu, P.; Xu, M.; Liu, W.-J.; Zhang, X.-Y.; Yao, H.-M.; Ye, X.-W. Low Plasma MiR-25 Expression Is a Favorite Prognosis Factor in Non-Small Cell Lung Cancer. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 5251–5259. [Google Scholar] [CrossRef]
- Niu, Y.; Su, M.; Wu, Y.; Fu, L.; Kang, K.; Li, Q.; Li, L.; Hui, G.; Li, F.; Gou, D. Circulating Plasma MiRNAs as Potential Biomarkers of Non-Small Cell Lung Cancer Obtained by High-Throughput Real-Time PCR Profiling. Cancer Epidemiol. Biomark. Prev. 2019, 28, 327–336. [Google Scholar] [CrossRef]
- Sun, Y.; Mei, H.; Xu, C.; Tang, H.; Wei, W. Circulating MicroRNA-339-5p and -21 in Plasma as an Early Detection Predictors of Lung Adenocarcinoma. Pathol. Res. Pract. 2018, 214, 119–125. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhao, H.; Gao, X.; Wei, F.; Zhang, X.; Su, Y.; Wang, C.; Li, H.; Ren, X. Identification of a Three-MiRNA Signature as a Blood-Borne Diagnostic Marker for Early Diagnosis of Lung Adenocarcinoma. Oncotarget 2016, 7, 26070–26086. [Google Scholar] [CrossRef] [PubMed]
- Feng, M.; Zhao, J.; Wang, L.; Liu, J. Upregulated Expression of Serum Exosomal MicroRNAs as Diagnostic Biomarkers of Lung Adenocarcinoma. Ann. Clin. Lab. Sci. 2018, 48, 712–718. [Google Scholar] [PubMed]
- Zhou, G.-H.; Yang, W.-H.; Sun, B. Clinical Impact of Serum MiR-661 in Diagnosis and Prognosis of Non-Small Cell Lung Cancer. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 5696–5701. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Li, T.; Chen, G.; Yan, G.; Zhang, X.; Wan, Y.; Li, Q.; Zhu, B.; Zhuo, W. Identification of a Serum MicroRNA Expression Signature for Detection of Lung Cancer, Involving MiR-23b, MiR-221, MiR-148b and MiR-423-3p. Lung Cancer 2017, 114, 6–11. [Google Scholar] [CrossRef] [PubMed]
- Shang, A.Q.; Xie, Y.N.; Wang, J.; Sun, L.; Wei, J.; Lu, W.Y.; Lan, J.Y.; Wang, W.W.; Wang, L.; Wang, L.L. Predicative Values of Serum MicroRNA-22 and MicroRNA-126 Levels for Non-Small Cell Lung Cancer Development and Metastasis: A Case-Control Study. Neoplasma 2017, 64, 453–459. [Google Scholar] [CrossRef]
- Sun, M.; Song, J.; Zhou, Z.; Zhu, R.; Jin, H.; Ji, Y.; Lu, Q.; Ju, H. Comparison of Serum MicroRNA21 and Tumor Markers in Diagnosis of Early Non-Small Cell Lung Cancer. Dis. Markers 2016, 2016, 3823121. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Ding, M.; Xia, M.; Chen, S.; Van Le, A.; Soto-Gil, R.; Shen, Y.; Wang, N.; Wang, J.; Gu, W.; et al. A Five-MiRNA Panel Identified From a Multicentric Case-Control Study Serves as a Novel Diagnostic Tool for Ethnically Diverse Non-Small-Cell Lung Cancer Patients. EBioMedicine 2015, 2, 1377–1385. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Gu, X.; Liu, F.; Rui, Z.; Liu, M.; Zhao, L. Antitumor Effects of Hsa-miR661-3p on Non-small Cell Lung Cancer in Vivo and in Vitro. Oncol. Rep. 2019, 41, 2987–2996. [Google Scholar] [CrossRef] [PubMed]
- Kunita, A.; Morita, S.; Irisa, T.U.; Goto, A.; Niki, T.; Takai, D.; Nakajima, J.; Fukayama, M. MicroRNA-21 in Cancer-Associated Fibroblasts Supports Lung Adenocarcinoma Progression. Sci. Rep. 2018, 8, 8838. [Google Scholar] [CrossRef]
- Luo, J.; Shi, K.; Yin, S.-Y.; Tang, R.-X.; Chen, W.-J.; Huang, L.-Z.; Gan, T.-Q.; Cai, Z.-W.; Chen, G. Clinical Value of MiR-182-5p in Lung Squamous Cell Carcinoma: A Study Combining Data from TCGA, GEO, and RT-QPCR Validation. World J. Surg. Oncol. 2018, 16, 76. [Google Scholar] [CrossRef] [PubMed]
- Tessema, M.; Yingling, C.M.; Picchi, M.A.; Wu, G.; Ryba, T.; Lin, Y.; Bungum, A.O.; Edell, E.S.; Spira, A.; Belinsky, S.A. ANK1 Methylation Regulates Expression of MicroRNA-486-5p and Discriminates Lung Tumors by Histology and Smoking Status. Cancer Lett. 2017, 410, 191–200. [Google Scholar] [CrossRef]
- Chen, L.-J.; Li, X.-Y.; Zhao, Y.-Q.; Liu, W.-J.; Wu, H.-J.; Liu, J.; Mu, X.-Q.; Wu, H.-B. Down-Regulated MicroRNA-375 Expression as a Predictive Biomarker in Non-Small Cell Lung Cancer Brain Metastasis and Its Prognostic Significance. Pathol. Res. Pract. 2017, 213, 882–888. [Google Scholar] [CrossRef]
- Daugaard, I.; Knudsen, A.; Kjeldsen, T.E.; Hager, H.; Hansen, L.L. The Association between MiR-34 Dysregulation and Distant Metastases Formation in Lung Adenocarcinoma. Exp. Mol. Pathol. 2017, 102, 484–491. [Google Scholar] [CrossRef]
- Xie, K.; Wang, C.; Qin, N.; Yang, J.; Zhu, M.; Dai, J.; Jin, G.; Shen, H.; Ma, H.; Hu, Z. Genetic Variants in Regulatory Regions of MicroRNAs Are Associated with Lung Cancer Risk. Oncotarget 2016, 7, 47966–47974. [Google Scholar] [CrossRef]
- Zhang, Y.; Xu, X.; Zhang, M.; Wang, X.; Bai, X.; Li, H.; Kan, L.; Zhou, Y.; Niu, H.; He, P. MicroRNA-663a Is Downregulated in Non-Small Cell Lung Cancer and Inhibits Proliferation and Invasion by Targeting JunD. BMC Cancer 2016, 16, 315. [Google Scholar] [CrossRef]
- Xie, K.; Ma, H.; Liang, C.; Wang, C.; Qin, N.; Shen, W.; Gu, Y.; Yan, C.; Zhang, K.; Dai, N.; et al. A Functional Variant in MiR-155 Regulation Region Contributes to Lung Cancer Risk and Survival. Oncotarget 2015, 6, 42781–42792. [Google Scholar] [CrossRef]
- Tang, R.; Zhong, T.; Dang, Y.; Zhang, X.; Li, P.; Chen, G. Association between Downexpression of MiR-203 and Poor Prognosis in Non-Small Cell Lung Cancer Patients. Clin. Transl. Oncol. 2016, 18, 360–368. [Google Scholar] [CrossRef]
- Kotsyfakis, M.; Patelarou, E. MicroRNAs as Biomarkers of Harmful Environmental and Occupational Exposures: A Systematic Review. Biomarkers 2019, 24, 623–630. [Google Scholar] [CrossRef]
- Mancini, F.R.; Laine, J.E.; Tarallo, S.; Vlaanderen, J.; Vermeulen, R.; van Nunen, E.; Hoek, G.; Probst-Hensch, N.; Imboden, M.; Jeong, A.; et al. MicroRNA Expression Profiles and Personal Monitoring of Exposure to Particulate Matter. Environ. Pollut. 2020, 263, 114392. [Google Scholar] [CrossRef]
- Liu, Q.; Wang, W.; Jing, W. Indoor Air Pollution Aggravates Asthma in Chinese Children and Induces the Changes in Serum Level of MiR-155. Int. J. Environ. Health Res. 2019, 29, 22–30. [Google Scholar] [CrossRef]
- Krauskopf, J.; van Veldhoven, K.; Chadeau-Hyam, M.; Vermeulen, R.; Carrasco-Turigas, G.; Nieuwenhuijsen, M.; Vineis, P.; de Kok, T.M.; Kleinjans, J.C. Short-Term Exposure to Traffic-Related Air Pollution Reveals a Compound-Specific Circulating MiRNA Profile Indicating Multiple Disease Risks. Environ. Int. 2019, 128, 193–200. [Google Scholar] [CrossRef]
- Tsamou, M.; Vrijens, K.; Madhloum, N.; Lefebvre, W.; Vanpoucke, C.; Nawrot, T.S. Air Pollution-Induced Placental Epigenetic Alterations in Early Life: A Candidate MiRNA Approach. Epigenetics 2018, 13, 135–146. [Google Scholar] [CrossRef]
- Rodosthenous, R.S.; Kloog, I.; Colicino, E.; Zhong, J.; Herrera, L.A.; Vokonas, P.; Schwartz, J.; Baccarelli, A.A.; Prada, D. Extracellular Vesicle-Enriched MicroRNAs Interact in the Association between Long-Term Particulate Matter and Blood Pressure in Elderly Men. Environ. Res. 2018, 167, 640–649. [Google Scholar] [CrossRef] [PubMed]
- Krauskopf, J.; Caiment, F.; van Veldhoven, K.; Chadeau-Hyam, M.; Sinharay, R.; Chung, K.F.; Cullinan, P.; Collins, P.; Barratt, B.; Kelly, F.J.; et al. The Human Circulating MiRNome Reflects Multiple Organ Disease Risks in Association with Short-Term Exposure to Traffic-Related Air Pollution. Environ. Int. 2018, 113, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Espín-Pérez, A.; Krauskopf, J.; Chadeau-Hyam, M.; van Veldhoven, K.; Chung, F.; Cullinan, P.; Piepers, J.; van Herwijnen, M.; Kubesch, N.; Carrasco-Turigas, G.; et al. Short-Term Transcriptome and MicroRNAs Responses to Exposure to Different Air Pollutants in Two Population Studies. Environ. Pollut. 2018, 242, 182–190. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Li, H.; Cai, J.; Wang, C.; Lin, Z.; Liu, C.; Niu, Y.; Zhao, Z.; Li, W.; Kan, H. Fine Particulate Air Pollution and the Expression of MicroRNAs and Circulating Cytokines Relevant to Inflammation, Coagulation, and Vasoconstriction. Environ. Health Perspect. 2018, 126, 017007. [Google Scholar] [CrossRef] [PubMed]
- Pergoli, L.; Cantone, L.; Favero, C.; Angelici, L.; Iodice, S.; Pinatel, E.; Hoxha, M.; Dioni, L.; Letizia, M.; Albetti, B.; et al. Extracellular Vesicle-Packaged MiRNA Release after Short-Term Exposure to Particulate Matter Is Associated with Increased Coagulation. Part. Fibre Toxicol. 2017, 14, 32. [Google Scholar] [CrossRef]
- Vriens, A.; Nawrot, T.S.; Saenen, N.D.; Provost, E.B.; Kicinski, M.; Lefebvre, W.; Vanpoucke, C.; Van Deun, J.; De Wever, O.; Vrijens, K.; et al. Recent Exposure to Ultrafine Particles in School Children Alters MiR-222 Expression in the Extracellular Fraction of Saliva. Environ. Health 2016, 15, 80. [Google Scholar] [CrossRef] [PubMed]
- Rodosthenous, R.S.; Coull, B.A.; Lu, Q.; Vokonas, P.S.; Schwartz, J.D.; Baccarelli, A.A. Ambient Particulate Matter and MicroRNAs in Extracellular Vesicles: A Pilot Study of Older Individuals. Part. Fibre Toxicol. 2016, 13, 13. [Google Scholar] [CrossRef] [PubMed]
- Pavanello, S.; Bonzini, M.; Angelici, L.; Motta, V.; Pergoli, L.; Hoxha, M.; Cantone, L.; Pesatori, A.C.; Apostoli, P.; Tripodi, A.; et al. Extracellular Vesicle-Driven Information Mediates the Long-Term Effects of Particulate Matter Exposure on Coagulation and Inflammation Pathways. Toxicol. Lett. 2016, 259, 143–150. [Google Scholar] [CrossRef]
- Motta, V.; Favero, C.; Dioni, L.; Iodice, S.; Battaglia, C.; Angelici, L.; Vigna, L.; Pesatori, A.C.; Bollati, V. MicroRNAs Are Associated with Blood-Pressure Effects of Exposure to Particulate Matter: Results from a Mediated Moderation Analysis. Environ. Res. 2016, 146, 274–281. [Google Scholar] [CrossRef]
- Louwies, T.; Vuegen, C.; Panis, L.I.; Cox, B.; Vrijens, K.; Nawrot, T.S.; De Boever, P. MiRNA Expression Profiles and Retinal Blood Vessel Calibers Are Associated with Short-Term Particulate Matter Air Pollution Exposure. Environ. Res. 2016, 147, 24–31. [Google Scholar] [CrossRef]
- Li, X.; Lv, Y.; Hao, J.; Sun, H.; Gao, N.; Zhang, C.; Lu, R.; Wang, S.; Yin, L.; Pu, Y.; et al. Role of MicroRNA-4516 Involved Autophagy Associated with Exposure to Fine Particulate Matter. Oncotarget 2016, 7, 45385–45397. [Google Scholar] [CrossRef]
- Hou, L.; Barupal, J.; Zhang, W.; Zheng, Y.; Liu, L.; Zhang, X.; Dou, C.; McCracken, J.P.; Díaz, A.; Motta, V.; et al. Particulate Air Pollution Exposure and Expression of Viral and Human MicroRNAs in Blood: The Beijing Truck Driver Air Pollution Study. Environ. Health Perspect. 2016, 124, 344–350. [Google Scholar] [CrossRef]
- Bollati, V.; Angelici, L.; Rizzo, G.; Pergoli, L.; Rota, F.; Hoxha, M.; Nordio, F.; Bonzini, M.; Tarantini, L.; Cantone, L.; et al. Microvesicle-Associated MicroRNA Expression Is Altered upon Particulate Matter Exposure in Healthy Workers and in A549 Cells. J. Appl. Toxicol. 2014, 35, 59–67. [Google Scholar] [CrossRef]
- Fossati, S.; Baccarelli, A.; Zanobetti, A.; Hoxha, M.; Vokonas, P.S.; Wright, R.O.; Schwartz, J. Ambient Particulate Air Pollution and MicroRNAs in Elderly Men. Epidemiology 2014, 25, 68–78. [Google Scholar] [CrossRef]
- Velasco-Torres, Y.; Ruiz, V.; Montaño, M.; Pérez-Padilla, R.; Falfán-Valencia, R.; Pérez-Ramos, J.; Pérez-Bautista, O.; Ramos, C. Participation of the MiR-22-HDAC4-DLCO Axis in Patients with COPD by Tobacco and Biomass. Biomolecules 2019, 9, 837. [Google Scholar] [CrossRef]
- Khan, A.; Thatcher, T.H.; Woeller, C.F.; Sime, P.J.; Phipps, R.P.; Hopke, P.K.; Utell, M.J.; Krahl, P.L.; Mallon, T.M.; Thakar, J. Machine Learning Approach for Predicting Past Environmental Exposures From Molecular Profiling of Post-Exposure Human Serum Samples. J. Occup. Environ. Med. 2019, 61, S55–S64. [Google Scholar] [CrossRef]
- Du, Y.; Ding, Y.; Chen, X.; Mei, Z.; Ding, H.; Wu, Y.; Jie, Z. MicroRNA-181c Inhibits Cigarette Smoke–Induced Chronic Obstructive Pulmonary Disease by Regulating CCN1 Expression. Respir. Res. 2017, 18, 155. [Google Scholar] [CrossRef]
- Herberth, G.; Bauer, M.; Gasch, M.; Hinz, D.; Röder, S.; Olek, S.; Kohajda, T.; Rolle-Kampczyk, U.; von Bergen, M.; Sack, U.; et al. Maternal and Cord Blood MiR-223 Expression Associates with Prenatal Tobacco Smoke Exposure and Low Regulatory T-Cell Numbers. J. Allergy Clin. Immunol. 2014, 133, 543–550. [Google Scholar] [CrossRef]
- Marczylo, E.L.; Amoako, A.A.; Konje, J.C.; Gant, T.W.; Marczylo, T.H. Smoking Induces Differential MiRNA Expression in Human Spermatozoa: A Potential Transgenerational Epigenetic Concern? Epigenetics 2012, 7, 432–439. [Google Scholar] [CrossRef]
- Ruiz-Vera, T.; Ochoa-Martínez, Á.C.; Pruneda-Álvarez, L.G.; Zarazúa, S.; Pérez-Maldonado, I.N. Exposure to Biomass Smoke Is Associated with an Increased Expression of Circulating MiRNA-126 and MiRNA-155 in Mexican Women: A Pilot Study. Drug Chem. Toxicol. 2019, 42, 335–342. [Google Scholar] [CrossRef]
- Rider, C.F.; Yamamoto, M.; Günther, O.P.; Hirota, J.A.; Singh, A.; Tebbutt, S.J.; Carlsten, C. Controlled Diesel Exhaust and Allergen Coexposure Modulates MicroRNA and Gene Expression in Humans: Effects on Inflammatory Lung Markers. J. Allergy Clin. Immunol. 2016, 138, 1690–1700. [Google Scholar] [CrossRef]
- Song, M.-K.; Ryu, J.-C. Blood MiRNAs as Sensitive and Specific Biological Indicators of Environmental and Occupational Exposure to Volatile Organic Compound (VOC). Int. J. Hyg. Environ. Health 2015, 218, 590–602. [Google Scholar] [CrossRef]
- Fry, R.C.; Rager, J.E.; Bauer, R.; Sebastian, E.; Peden, D.B.; Jaspers, I.; Alexis, N.E. Air Toxics and Epigenetic Effects: Ozone Altered MicroRNAs in the Sputum of Human Subjects. Am. J. Physiol. Lung Cell. Mol. Physiol. 2014, 306, L1129–L1137. [Google Scholar] [CrossRef]
- Garofalo, M.; Di Leva, G.; Romano, G.; Nuovo, G.; Suh, S.-S.; Ngankeu, A.; Taccioli, C.; Pichiorri, F.; Alder, H.; Secchiero, P.; et al. MiR-221&222 Regulate TRAIL Resistance and Enhance Tumorigenicity through PTEN and TIMP3 Downregulation. Cancer Cell 2009, 16, 498–509. [Google Scholar] [CrossRef]
- Amini, S.; Abak, A.; Sakhinia, E.; Abhari, A. MicroRNA-221 and MicroRNA-222 in Common Human Cancers: Expression, Function, and Triggering of Tumor Progression as a Key Modulator. Lab. Med. 2019, 50, 333–347. [Google Scholar] [CrossRef]
- Zhang, L.; Hao, C.; Zhai, R.; Wang, D.; Zhang, J.; Bao, L.; Li, Y.; Yao, W. Downregulation of Exosomal Let-7a-5p in Dust Exposed- Workers Contributes to Lung Cancer Development. Respir. Res. 2018, 19, 235. [Google Scholar] [CrossRef]
- Li, X.; Wei, Y.; Wang, Z. MicroRNA-21 and Hypertension. Hypertens. Res. 2018, 41, 649–661. [Google Scholar] [CrossRef]
- Bica-Pop, C.; Cojocneanu-Petric, R.; Magdo, L.; Raduly, L.; Gulei, D.; Berindan-Neagoe, I. Overview upon MiR-21 in Lung Cancer: Focus on NSCLC. Cell. Mol. Life Sci. 2018, 75, 3539–3551. [Google Scholar] [CrossRef]
- Gao, Y.; Deng, K.; Liu, X.; Dai, M.; Chen, X.; Chen, J.; Chen, J.; Huang, Y.; Dai, S.; Chen, J. Molecular Mechanism and Role of MicroRNA-93 in Human Cancers: A Study Based on Bioinformatics Analysis, Meta-analysis, and Quantitative Polymerase Chain Reaction Validation. J. Cell. Biochem. 2019, 120, 6370–6383. [Google Scholar] [CrossRef]
- Micolucci, L.; Akhtar, M.M.; Olivieri, F.; Rippo, M.R.; Procopio, A.D. Diagnostic Value of MicroRNAs in Asbestos Exposure and Malignant Mesothelioma: Systematic Review and Qualitative Meta-Analysis. Oncotarget 2016, 7, 58606–58637. [Google Scholar] [CrossRef]
- Liu, R.; Zhang, Y.S.; Zhang, S.; Cheng, Z.M.; Yu, J.L.; Zhou, S.; Song, J. MiR-126-3p Suppresses the Growth, Migration and Invasion of NSCLC via Targeting CCR1. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 679–689. [Google Scholar] [CrossRef]
- Chen, Q.; Hu, H.; Jiao, D.; Yan, J.; Xu, W.; Tang, X.; Chen, J.; Wang, J. MiR-126-3p and MiR-451a Correlate with Clinicopathological Features of Lung Adenocarcinoma: The Underlying Molecular Mechanisms. Oncol. Rep. 2016, 36, 909–917. [Google Scholar] [CrossRef]
- Mitra, R.; Adams, C.M.; Jiang, W.; Greenawalt, E.; Eischen, C.M. Pan-Cancer Analysis Reveals Cooperativity of Both Strands of MicroRNA That Regulate Tumorigenesis and Patient Survival. Nat. Commun. 2020, 11, 968. [Google Scholar] [CrossRef]
- Gulei, D.; Raduly, L.; Broseghini, E.; Ferracin, M.; Berindan-Neagoe, I. The Extensive Role of MiR-155 in Malignant and Non-Malignant Diseases. Mol. Asp. Med. 2019, 70, 33–56. [Google Scholar] [CrossRef] [PubMed]
- Mahesh, G.; Biswas, R. MicroRNA-155: A Master Regulator of Inflammation. J. Interferon Cytokine Res. 2019, 39, 321–330. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Li, Z.; Ge, Q.; Wu, W.; Zhu, Q.; Luo, J.; Chen, L. Characterization of MicroRNA Transcriptome in Tumor, Adjacent, and Normal Tissues of Lung Squamous Cell Carcinoma. J. Thorac. Cardiovasc. Surg. 2015, 149, 1404–1414.e4. [Google Scholar] [CrossRef]
- Fu, Y.; Li, Y.; Wang, X.; Li, F.; Lu, Y. Overexpression of MiR-425-5p Is Associated with Poor Prognosis and Tumor Progression in Non-Small Cell Lung Cancer. Cancer Biomark. 2020, 27, 147–156. [Google Scholar] [CrossRef]
- Boxberger, N.; Hecker, M.; Zettl, U.K. Dysregulation of Inflammasome Priming and Activation by MicroRNAs in Human Immune-Mediated Diseases. J. Immunol. 2019, 202, 2177–2187. [Google Scholar] [CrossRef]
- Esteves, J.V.; Enguita, F.J.; Machado, U.F. MicroRNAs-Mediated Regulation of Skeletal Muscle GLUT4 Expression and Translocation in Insulin Resistance. J. Diabetes Res. 2017, 2017, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Gomez, J.L.; Chen, A.; Diaz, M.P.; Zirn, N.; Gupta, A.; Britto, C.; Sauler, M.; Yan, X.; Stewart, E.; Santerian, K.; et al. A Network of Sputum MicroRNAs Is Associated with Neutrophilic Airway Inflammation in Asthma. Am. J. Respir. Crit. Care Med. 2020, 202, 51–64. [Google Scholar] [CrossRef]
- Luo, P.; Wang, Q.; Ye, Y.; Zhang, J.; Lu, D.; Cheng, L.; Zhou, H.; Xie, M.; Wang, B. MiR-223-3p Functions as a Tumor Suppressor in Lung Squamous Cell Carcinoma by MiR-223-3p-Mutant P53 Regulatory Feedback Loop. J. Exp. Clin. Cancer Res. 2019, 38, 74. [Google Scholar] [CrossRef]
- IARC Working Group on the Evaluation of Carcinogenic Risks to Humans. In Outdoor Air Pollution; International Agency for Research on Cancer: Lyon, France, 2015; ISBN 978-92-832-0147-2.
- Rossnerova, A.; Pokorna, M.; Svecova, V.; Sram, R.J.; Topinka, J.; Zölzer, F.; Rossner, P. Adaptation of the Human Population to the Environment: Current Knowledge, Clues from Czech Cytogenetic and “Omics” Biomonitoring Studies and Possible Mechanisms. Mutat. Res. Rev. Mutat. Res. 2017, 773, 188–203. [Google Scholar] [CrossRef]
- Vineis, P.; Chatziioannou, A.; Cunliffe, V.T.; Flanagan, J.M.; Hanson, M.; Kirsch-Volders, M.; Kyrtopoulos, S. Epigenetic Memory in Response to Environmental Stressors. FASEB J. 2017, 31, 2241–2251. [Google Scholar] [CrossRef]
- Rossnerova, A.; Izzotti, A.; Pulliero, A.; Bast, A.; Rattan, S.I.S.; Rossner, P. The Molecular Mechanisms of Adaptive Response Related to Environmental Stress. Int. J. Mol. Sci. 2020, 21, 7053. [Google Scholar] [CrossRef] [PubMed]


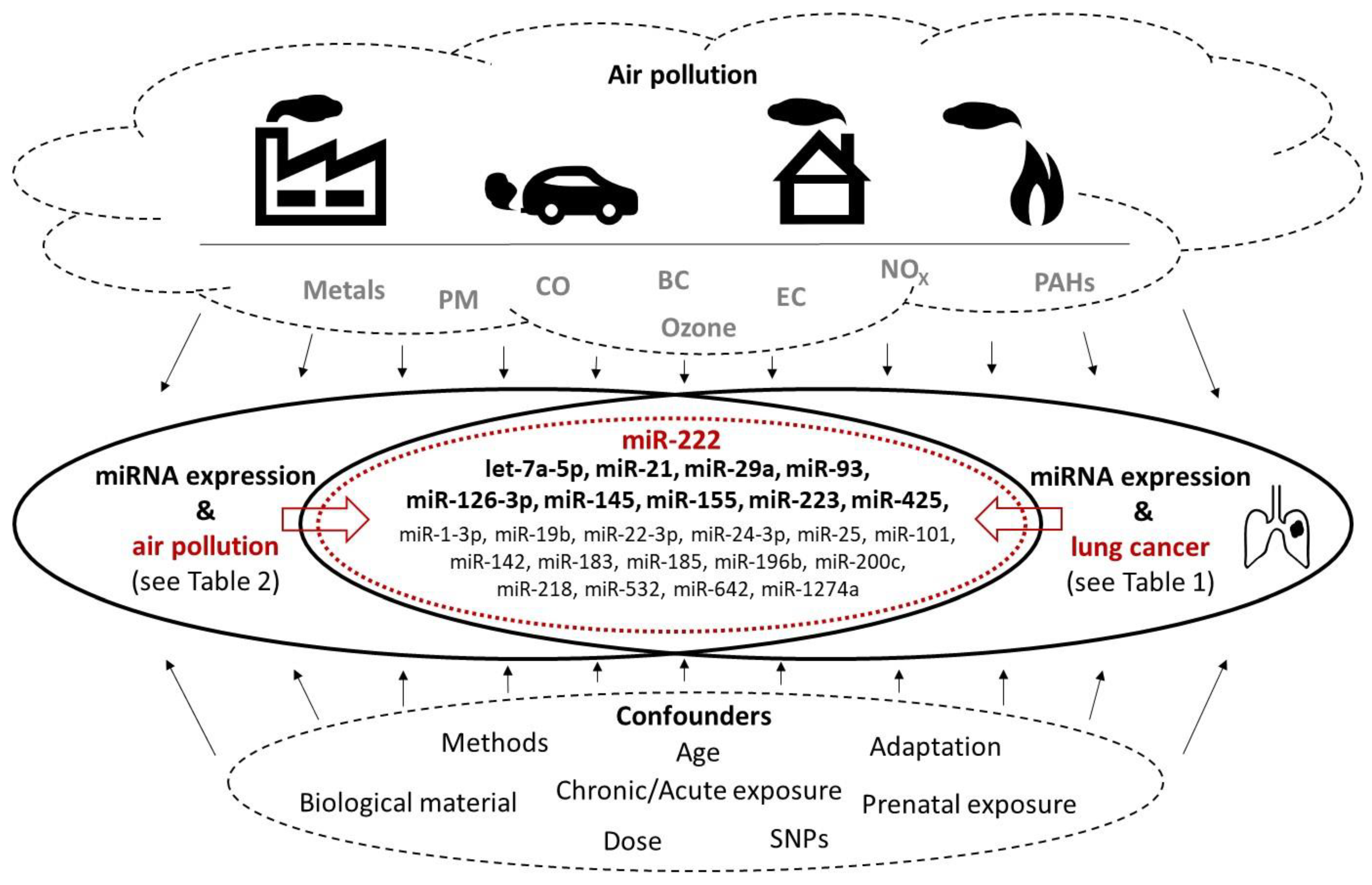{kind=link}
| miRNA | Tissue | Patients | Main Output | Method | Reference |
|---|---|---|---|---|---|
| let-7a-5p, miR-214-3p, miR-1291, miR-1-3p, miR-375 | Serum | 744 | DB | qRT-PCR | [64] |
| miR-29a-5p, miR-4491, miR-542-3p, miR-135a-5p | Blood | 145 | DB | Microarray + qRT-PCR | [33] |
| miR-2114, miR-2115, miR-449c | Blood | NS | DB | qRT-PCR | [34] |
| miR-210-3p, miR-126-3p, miR-145, miR-205-5p | Plasma | 471 | DB | qRT-PCR | [65] |
| miR-25 | Plasma | 114 | DB | qRT-PCR | [66] |
| miR-26a-5p, miR-126-5p, miR-139-5p, miR-152-3p, miR-200c-3p, miR-3135b, miR-151a-3p, miR-151a-5p, miR-151b, miR-550a-3p | Plasma | 437 | DB | qRT-PCR | [67] |
| miR-339-5p, miR-21 | Plasma | 28 | DB | Microarray + qRT-PCR | [68] |
| miR-532, miR-628-3p, miR-425 | Plasma | 201 | DB | qRT-PCR | [69] |
| let-7c, miR-122, miR-182, miR-193a-5p, miR-200c, miR-203, miR-218, miR-155, let-7b, miR-411, miR-450b-5p, miR-485-3p, miR-519a, miR-642, miR-517b, miR-520f, miR-206, miR-566, miR-661, miR-340, miR-1243, miR-720, miR-543, miR-1267 | Plasma | 100 | DB | Microarray | [35] |
| miR-107 | Serum | NS | OS | qRT-PCR | [54] |
| miR-223 | Serum | 75 | DB | ddPCR | [40] |
| miR-21-5p, miR-140-5p, miR-126-3p | Serum | 23 | DB | qRT-PCR | [70] |
| miR-661 | Serum | 150 | DB | qRT-PCR | [71] |
| miR-23b, miR-423-3p, miR-148b, miR-221 | Serum | 50 | DB | qRT-PCR | [72] |
| miR-21 | Serum | 50 | DB | NS | [41] |
| miR-22, miR-126 | Serum | 127 | DB | qRT-PCR | [73] |
| miR-451, miR-1290, miR-636, miR-30c, miR-22-3p, miR-19b, miR-486-5p, miR-20b, miR-93, miR-34b, miR-185, miR-126-5p, miR-93-3p, miR-1274a, miR-142-5p, miR-628-5p, miR-486-3p, miR-425, miR-645, miR-24 | Serum | 253 | DB | Microarray | [36] |
| miR-21 | Serum | 50 | DB | qRT-PCR | [74] |
| miR-483-5p, miR-193a-3p, miR-25, miR-214, miR-7 | Serum | 221 | DB | Microarray + RT-PCR | [75] |
| miR-196b-5p | Tissue | 713 | PTT | qRT-PCR | [62] |
| miR-497 | Tissue | 15 | OS | qRT-PCR | [49] |
| miR- 661-3p | Tissue | 12 | DB | qRT-PCR | [76] |
| miR-99a-5p | Tissue | 50 | OG | qRT-PCR | [46] |
| miR-29a | Tissue | 55 | OG | qRT-PCR | [44] |
| miR-101 | Tissue | 200 | PTT | qRT-PCR | [61] |
| miR-21 | Tissue | 89 | DB | qRT-PCR | [77] |
| miR-1275 | Tissue | 70 | OG | qRT-PCR | [45] |
| miR-12528 | Tissue | 20 | PTT | qRT-PCR | [59] |
| miR-182-5p | Tissue | 23 | DB | qRT-PCR | [78] |
| miR-491-5p | Tissue | 100 | PTT | qRT-PCR | [57] |
| miR-1260b | Tissue | 26 | PTT | qRT-PCR | [60] |
| miR-198-5p | Tissue | 23 | OG | Microarray + qRT-PCR | [43] |
| miR-454 | Tissue | 67 | OS | qRT-PCR | [53] |
| miR-504 | Tissue | 55 | OS | qRT-PCR | [52] |
| miR-7a, miR-24-3p | Tissue | 25/50 | PTT | ddPCR | [38,39] |
| miR-486-5p | Tissue | 262 | DB | qRT-PCR | [79] |
| miR-140-5p | Tissue | 19 | PTT | qRT-PCR | [58] |
| miR-103 | Tissue | 32 | PTT | qRT-PCR | [56] |
| miR-219 | Tissue | 32 | OS | qRT-PCR | [51] |
| miR-451 | Tissue | 72 | OS | qRT-PCR | [50] |
| miR-375 | Tissue | 60 | DB | qRT-PCR | [80] |
| miR-675 | Tissue | 92 | OG | qRT-PCR | [42] |
| miR-34b, miR-34c | Tissue | 52 | DB | qRT-PCR | [81] |
| miR-588 | Tissue | 85 | PTT | qRT-PCR | [55] |
| miR-200a-3p, miR-200a-5p, miR-200b-3p, miR-200b-5p, miR-429 | Tissue | 1341 | DB | qRT-PCR | [82] |
| miR-663a | Tissue | 62 | DB | qRT-PCR | [83] |
| miR-218-5p | Tissue | NS | OS | qRT-PCR | [48] |
| miR-155 | Tissue | 1341 | DB | qRT-PCR | [84] |
| miR-203 | Tissue | 125 | DB | qRT-PCR | [85] |
| miR-34c, miR-183, miR-210 | Tissue | 103 | OS, OG, OG | qRT-PCR | [47] |
| miR-23b | Tissue | NS | PTT | qRT-PCR | [63] |
| miR-34b-3p | Tissue | 100 | PTT | Microarray | [37] |
| Pollutant | miRNA | Tissue | Subjects/Characteristics | Method | Reference |
|---|---|---|---|---|---|
| Effects of Environmental Air Pollutants | |||||
| PM2.5, UFP (PM0.1), black carbon, soot | miR-24-3p, miR-4454, miR-4763-3p, miR-425-5p, let-7d-5p, miR-502-5p, and miR-505-3p were associated with PM2.5 exposure | Blood | 143, healthy | Microarray | [87] |
| PM10, PM2.5, PM0.1, HCHO, NO2 | miR-155 was associated with PM2.5 and HCHO exposure in the asthma group | Serum | 180, healthy/180 asthmatic children | qRT-PCR | [88] |
| PM10, PM2.5, NO, NO2, CO, CO2, BC and UFP | miR-28-3p, miR-222-3p, miR-146-5p, miR-30b-5p/30c-5p, miR-320a-3p/320b/320c/320d/320e, miR-532-5p, miR-192-5p/215-5p, miR-144-3p, miR-425-5p were associated with exposure to a mixture of pollutants; no effect for PM10 or PM2.5 alone | Plasma | 24, healthy | NGS | [89] |
| PM2.5 | Negative link of miR-21, miR-146a and miR-222 expression with PM2.5 in 2nd trimester; positive link of miR-20a and miR-21 in 1st trimester exposure | Placenta | 210 newborns | qRT-PCR | [90] |
| PM2.5 | miR-199a/b and miR-223–3p modified links between PM2.5 and systolic blood pressure | Extracellular vesicles | 22 healthy elderly | NanoStringnCounter® platform | [91] |
| PM10, PM2.5, black carbon, ultrafine particles and NO2 | 54 miRNAs associated with exposure | Plasma | 24 healthy/ischemic heart disease/COPD | NGS | [92] |
| PM10, PM2.5, NO, NO2, CO, CO2, BC and UFP | miR-197-3p, miR-29a-3p, miR-15a-5p, miR-16-5p and miR-92a-3p associated with exposure to pollutants | Blood | 50 healthy, 20 COPD, 19 ischemic heart disease | Sureprint G3 Human V19 miRNA 8 x 60K (Agilent) | [93] |
| PM2.5 | The expression of miR-21-5p, miR-187-3p, miR-146a-5p, miR-1-3p, miR-199a-5p was associated with the exposure | Blood | 55 healthy | qRT-PCR | [94] |
| PM10 | The expression of let-7c-5p; miR-106a-5p; miR-143-3p; miR-185-5p; miR-218-5p; miR-331-3p; miR-642-5p; miR-652-3p; miR-99b-5p was downregulated | Extracellular vesicles | 1630 overweight/obese | QuantStudio™ 12 K Flex Real Time PCR | [95] |
| PM2.5, UFP | miR-222 expression affected by UFP, but not PM2.5; no effect was observed for miR-146a | Saliva | 80 healthy children | qRT-PCR | [96] |
| PM2.5 | Expression of miR-126-3p, miR-19b-3p, miR-93-5p, miR-223-3p, miR-142-3p, miR-23a-3p, miR-150-5p, miR-15a-5p, miR-191-5p, let-7a-5p affected by the exposure | Serum | 22 healthy | NanoStringnCounter® platform | [97] |
| PM and associated metals | 17 miRNAs affected by the exposure, including mir-196b, miR-302b, miR-200c, miR-30d | Extracellular vesicles | 55 healthy steel plant workers | qRT-PCR | [98] |
| PM10 | miR-145, miR-197, miR-30b, miR-345, miR-26a, miR-425-5p, miR-331, miR-140-3p, miR-101 associated with the exposure | Blood | 90 obese subjects | TaqMan Low-Density Array | [99] |
| PM10 | Negative link with miR-21, miR-222, but not -miR-146a expression | Blood | 50 healthy adults | qRT-PCR | [100] |
| PM2.5, metals | Positive link with miR-4516 | Serum | 120 healthy subjects | miRCURY LNATM | [101] |
| PM10, PM2.5, elemental carbon | No effect of PM2.5 exposure; PM10 affected 12 miRNAs; EC affected 28 miRNAs in the controls and 29 in truck drivers; miR-125a-5p, miR-1274a, miR-600, miR-1283, miR-10a were common in both groups | Blood | 120 healthy subjects (truck drivers and controls) | NanoStringnCounter® platform | [102] |
| PM | Increased expression of miR-128 and miR-302c | Extracellular vesicles | 63 healthy steel plant workers | qRT-PCR | [103] |
| PM2.5, black carbon, organic carbon, sulfates | Negative links with miR-1, miR-126, miR-135a, miR-146a, miR-155, miR-21, miR-222 and miR-9 | Blood | 153 elderly healthy men | qRT-PCR | [104] |
| Effects of Cigarette/Tobacco Smoke Exposure | |||||
| Biomass smoke (BS), tobacco smoke | miR-22-3p downregulated after BS exposure | Serum | 50, COPD | qRT-PCR | [105] |
| Cigarette smoke | Expression of miR-29a, miR-93, let-7a, and let-7g affected | Serum | 775, healthy, smokers/non-smokers | Low-density PCR array | [106] |
| Cigarette smoke | Modulation of miR-181c expression | Lung tissue | 34 COPD | qRT-PCR | [107] |
| Tobacco smoke | Positive link with miR-223, but not with miR-155 | Maternal and cord blood | 441 mothers and newborns | qRT-PCR | [108] |
| Cigarette smoke | 28 miRNAs differentially expressed in smokers when compared with non-smokers | Spermatozoa | 13 healthy smokers and non-smokers | miRCURY LNATM | [109] |
| Effects of Other Pollutants | |||||
| Wood and LPG exhaust | miR-126 and miR-155 upregulated after wood smoke exposure | Plasma | 52, healthy | qRT-PCR | [110] |
| Diesel exhaust, allergen | miR-183-5p, miR-324-5p and miR-132-3p induced by allergen; no modulatory effect of diesel exhaust | Bronchoalveolar lavage | 15 atopic subjects | NanoStringnCounter® platform | [111] |
| VOC | Specific miRNAs for exposure to individual VOCs | Blood | 50 healthy exposed workers | Microarray | [112] |
| Ozone | Increased expression of miR-132, miR-143, miR-145, miR-199a, miR-199b-5p, miR-222, miR-223, miR-25, miR-424, and miR-582-5p after the exposure | Sputum | 20 healthy volunteers | Microarray | [113] |
| miRNA | Pollutant | References-Air Pollutants | References-Lung Cancer |
|---|---|---|---|
| miR-222 | mixture of pollutants; PM2.5; UFP; PM10; black carbon, organic carbon, sulphates; ozone | [89,90,96,100,104,113] | [114] |
| let-7a-5p | PM2.5; Cigarette smoke | [87,97] | [64] |
| miR-21 | PM2.5; PM10; black carbon, organic carbon, sulphates | [90,94,100,104] | [41,68,74,77] |
| miR-29a family | Cigarette smoke | [93,106] | [44] |
| miR-93 family | PM2.5; Cigarette smoke | [97,106] | [36] |
| miR-126 family | PM2.5; black carbon, organic carbon, sulphates; wood smoke | [97,104,110] | [65,70] |
| miR-145 | PM10; ozone | [99,113] | [65] |
| miR-155 | PM2.5, HCHO; black carbon, organic carbon, sulphates; wood smoke | [88,104,108,110] | [35,84] |
| miR-223 | PM2.5; tobacco smoke; ozone | [91,97,108,113] | [40] |
| miR-425 family | PM2.5; mixture of pollutants; PM10 | [87,89,99] | [36,69] |
| miR-1-3p | PM2.5 | [94] | [64] |
| miR-19bfamily | PM2.5 | [97] | [36] |
| miR-22-3p | Biomass smoke (BS), tobacco smoke | [105] | [36] |
| miR-24-3p | PM2.5, UFP (PM0.1), black carbon, soot | [87] | [38,39] |
| miR-25 | Ozone | [113] | [66,75] |
| miR-101 | PM10 | [99] | [61] |
| miR-142 family | PM2.5 | [97] | [36] |
| miR-183 family | Diesel exhaust, allergen | [111] | [47] |
| miR-185 | PM10 | [95] | [36] |
| miR-196b family | PM and associated metals | [98] | [62] |
| miR-200c | PM and associated metals | [98] | [35,67] |
| miR-218-5p | PM10 | [95] | [48] |
| miR-532 | PM10, PM2.5, NO, NO2, CO, CO2, BC and UFP | [89] | [69] |
| miR-642 family | PM10 | [35] | |
| miR-1274a | PM10, PM2.5, elemental carbon | [102] | [36] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sima, M.; Rossnerova, A.; Simova, Z.; Rossner, P., Jr. The Impact of Air Pollution Exposure on the MicroRNA Machinery and Lung Cancer Development. J. Pers. Med. 2021, 11, 60. https://doi.org/10.3390/jpm11010060
Sima M, Rossnerova A, Simova Z, Rossner P Jr. The Impact of Air Pollution Exposure on the MicroRNA Machinery and Lung Cancer Development. Journal of Personalized Medicine. 2021; 11(1):60. https://doi.org/10.3390/jpm11010060
Chicago/Turabian StyleSima, Michal, Andrea Rossnerova, Zuzana Simova, and Pavel Rossner, Jr. 2021. "The Impact of Air Pollution Exposure on the MicroRNA Machinery and Lung Cancer Development" Journal of Personalized Medicine 11, no. 1: 60. https://doi.org/10.3390/jpm11010060
APA StyleSima, M., Rossnerova, A., Simova, Z., & Rossner, P., Jr. (2021). The Impact of Air Pollution Exposure on the MicroRNA Machinery and Lung Cancer Development. Journal of Personalized Medicine, 11(1), 60. https://doi.org/10.3390/jpm11010060