Genomic Signals of Adaptation towards Mutualism and Sociality in Two Ambrosia Beetle Complexes

 ,
,  , ,
, ,
Abstract

1. Introduction
2. Materials and Methods
2.1. Beetle Collection
2.2. Reared Beetles
2.3. RNA Isolation, Library Preparation and Sequencing
2.4. Data Processing, Assembly, and Functional Annotation
2.5. Data Availability
2.6. Ortholog Group Identification
2.7. Evolutionary Analysis
2.8. Differential Expression Analysis
3. Results
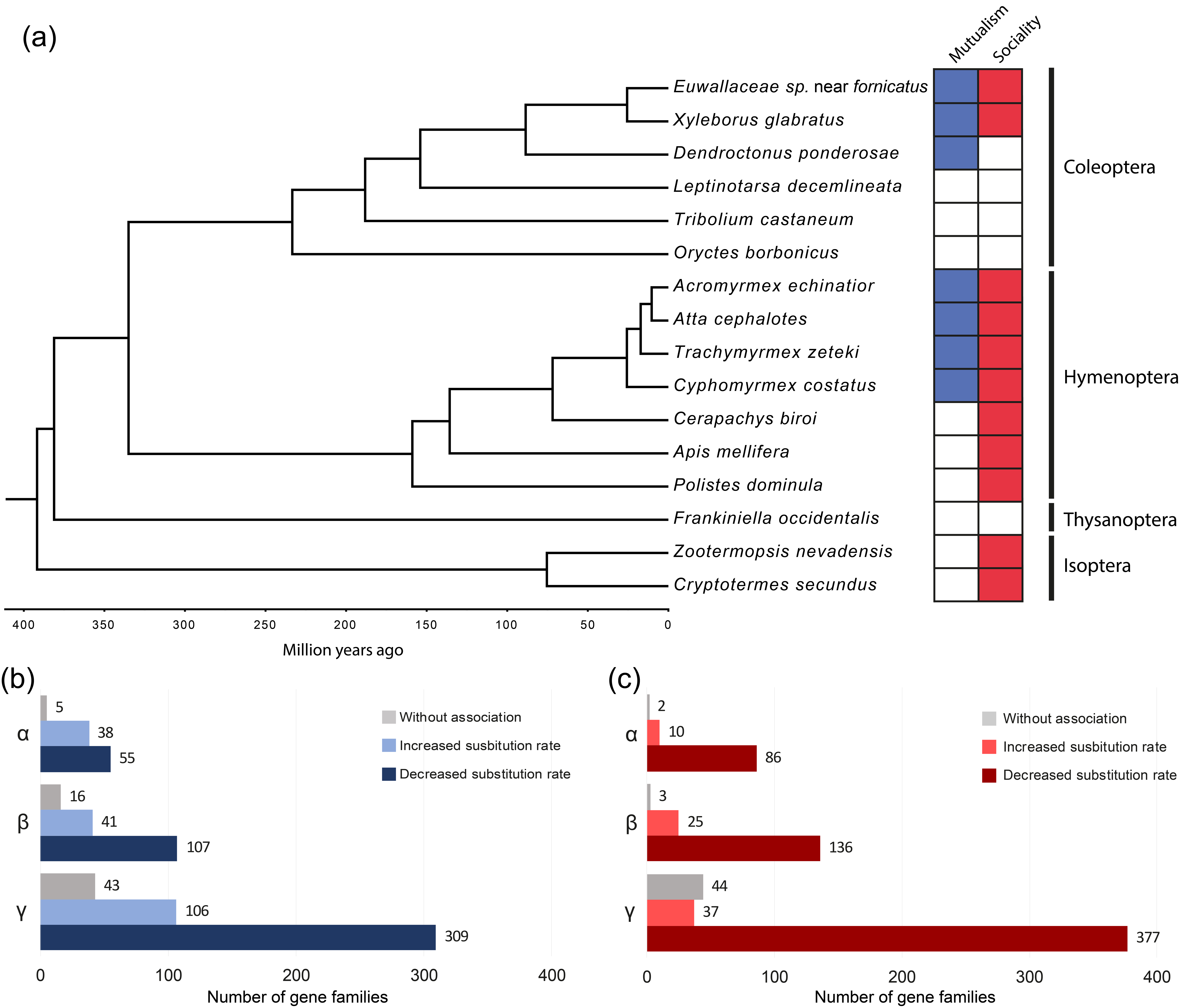
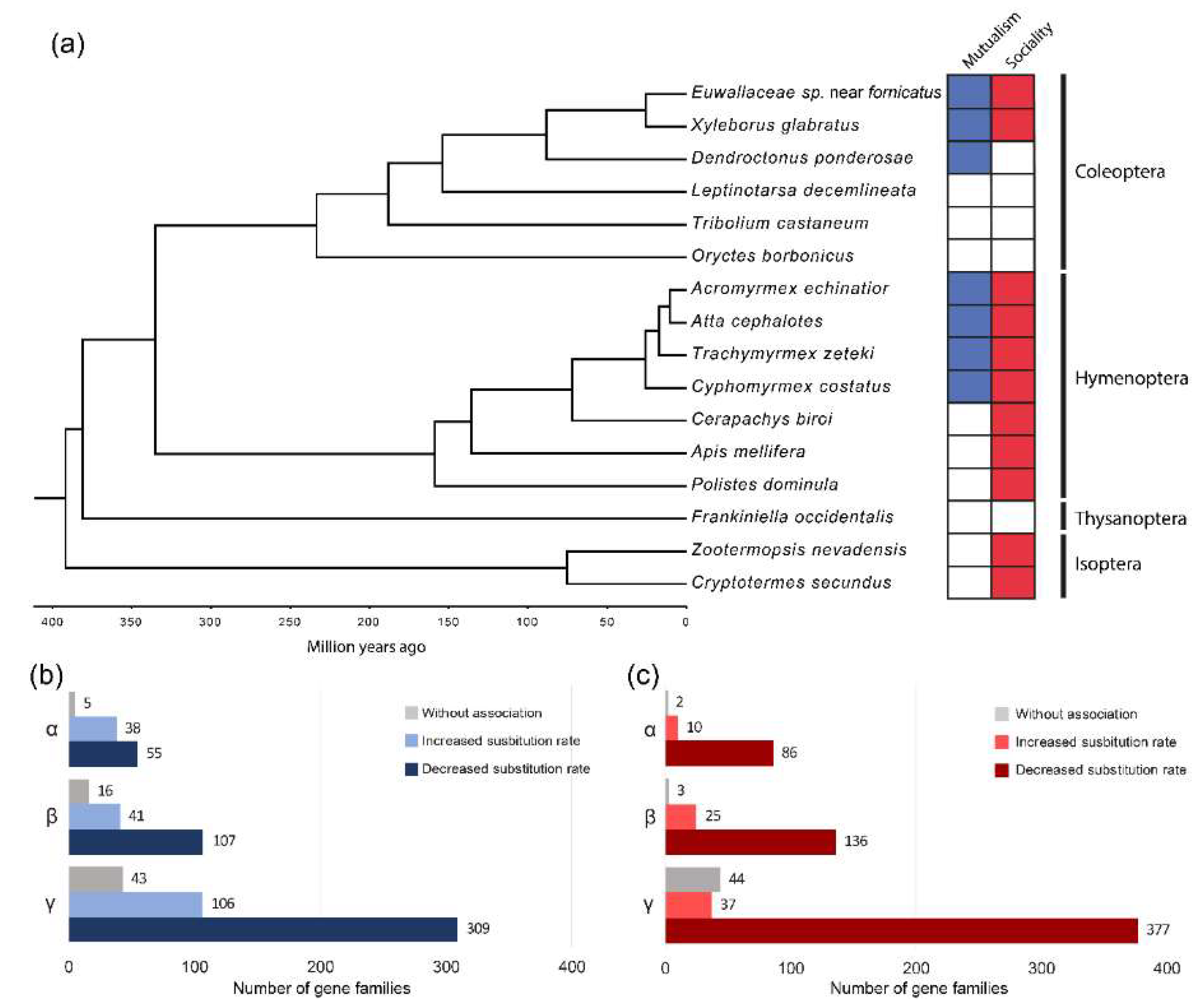
3.1. Gene Family Analysis
3.2. Genes under Selection in the Xyleborini Ambrosia Beetle Lineage
3.3. Changes in Evolutionary Rates Related to Fungus-Farming Mutualism and Sociality
4. Discussion
4.1. Evolutionary Changes Possibly Associated with Nutritional Symbiosis with Fungi
4.2. Evolutionary Changes Possibly Related to the Evolution of Behavior and Sociality.
4.3. Possible Causes of Accelerated or Decreased Evolutionary Rates
4.4. Transcriptional Fungal Profiles
5. Concluding Remarks
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mueller, U.G.; Gerardo, N. Fungus-farming insects: Multiple origins and diverse evolutionary histories. Proc. Natl. Acad. Sci. USA 2002, 99, 15247–15249. [Google Scholar] [CrossRef] [PubMed]
- Mueller, U.G.; Gerardo, N.M.; Aanen, D.K.; Six, D.L.; Schultz, T.R. The evolution of agriculture in insects. Annu. Rev. Ecol. Evol. Syst. 2005, 36, 563–595. [Google Scholar] [CrossRef]
- Jordal, B.H.; Cognato, A.I. Molecular phylogeny of bark and ambrosia beetles reveals multiple origins of fungus farming during periods of global warming. BMC Evol. Biol. 2012, 12, 133. [Google Scholar] [CrossRef] [PubMed]
- Johnson, A.J.; McKenna, D.D.; Jordal, B.H.; Cognato, A.I.; Smith, S.M.; Lemmon, A.R.; Lemmon, E.M.; Hulcr, J. Phylogenomics clarifies repeated evolutionary origins of inbreeding and fungus farming in bark beetles (Curculionidae, Scolytinae). Mol. Phylogenet. Evol. 2018, 127, 229–238. [Google Scholar] [CrossRef] [PubMed]
- Hulcr, J.; Stelinski, L.L. The ambrosia symbiosis: From evolutionary ecology to practical management. Annu. Rev. Entomol. 2017, 62, 285–303. [Google Scholar] [CrossRef] [PubMed]
- Vega, F.E.; Hofstetter, R.W. Bark Beetles: Biology and Ecology of Native and Invasive Species; Elsevier Science: Amsterdam, The Netherlands, 2014. [Google Scholar]
- Cruz, L.F.; Rocio, S.A.; Duran, L.G.; Menocal, O.; Garcia-Avila, C.D.J.; Carrillo, D. Developmental biology of Xyleborus bispinatus (Coleoptera: Curculionidae) reared on an artificial medium and fungal cultivation of symbiotic fungi in the beetle’s galleries. Fungal Ecol. 2018, 35, 116–126. [Google Scholar] [CrossRef]
- Kirkendall, L.R.; Biedermann, P.H.W.; Jordal, B.H. Chapter 3—Evolution and diversity of bark and ambrosia beetles. In Bark Beetles; Vega, F.E., Hofstetter, R.W., Eds.; Academic Press: San Diego, CA, USA, 2015; pp. 85–156. [Google Scholar]
- Bleiker, K.P.; Six, D.L. Dietary benefits of fungal associates to an eruptive herbivore: Potential implications of multiple associates on host population dynamics. Environ. Entomol. 2007, 36, 1384–1396. [Google Scholar] [CrossRef] [PubMed]
- Six, D.L. Ecological and evolutionary determinants of bark beetle—Fungus symbioses. Insects 2012, 3, 339–366. [Google Scholar] [CrossRef]
- Biedermann, P.H.W.; Rohlfs, M. Evolutionary feedbacks between insect sociality and microbial management. Curr. Opin. Insect Sci. 2017, 22, 92–100. [Google Scholar] [CrossRef] [PubMed]
- Biedermann, P.H.; Klepzig, K.D.; Taborsky, M. Fungus cultivation by ambrosia beetles: Behavior and laboratory breeding success in three Xyleborine species. Environ. Entomol. 2009, 38, 1096–1105. [Google Scholar] [CrossRef]
- Biedermann, P.H.; Taborsky, M. Larval helpers and age polyethism in ambrosia beetles. Proc. Natl. Acad. Sci. USA 2011, 108, 17064–17069. [Google Scholar] [CrossRef] [PubMed]
- Biedermann, P.H.; Klepzig, K.D.; Taborsky, M.; Six, D.L. Abundance and dynamics of filamentous fungi in the complex ambrosia gardens of the primitively eusocial beetle Xyleborinus saxesenii Ratzeburg (coleoptera: Curculionidae, scolytinae). FEMS Microbiol. Ecol. 2013, 83, 711–723. [Google Scholar] [CrossRef] [PubMed]
- Peer, K.; Taborsky, M. Delayed dispersal as a potential route to cooperative breeding in ambrosia beetles. Behav. Ecol. Sociobiol. 2007, 61, 729–739. [Google Scholar] [CrossRef]
- Biedermann, P.H.W.; Klepzig, K.D.; Taborsky, M. Costs of delayed dispersal and alloparental care in the fungus-cultivating ambrosia beetle Xyleborus affinis Eichhoff (Scolytinae: Curculionidae). Behav. Ecol. Sociobiol. 2011, 65, 1753–1761. [Google Scholar] [CrossRef]
- Kirkendall, L.R. Ecology and evolution of biased sex ratios in bark and ambrosia beetles. In Evolution and Diversity of Sex Ratio in Insects and Mites; Wrensch, D.L., Ebbert, M.A., Eds.; Chapman & Hall: New York, NY, USA, 1993; pp. 235–345. [Google Scholar]
- Normark, B.B.; Jordal, B.H.; Farrell, B.D. Origin of a haplodiploid beetle lineage. Proc. R. Soc. B Biol. Sci. 1999, 266, 2253–2253. [Google Scholar] [CrossRef]
- Peer, K.; Taborsky, M. Outbreeding depression, but no inbreeding depression in haplodiploid ambrosia beetles with regular sibling mating. Evol. Int. J. Org. Evol. 2005, 59, 317–323. [Google Scholar] [CrossRef]
- Kirkendall, L.R.; Kent, D.S.; Raffa, K.F. Interactions among males, females and offspring in bark and ambrosia beetles: The significance of living in tunnels for the evolution of social behavior. In The Evolution of Social Behaviour in Insects and Arachnids; Crespi, B.J., Choe, J.C., Eds.; Cambridge University Press: Cambridge, UK, 1997; pp. 181–215. [Google Scholar]
- Shik, J.Z.; Santos, J.C.; Seal, J.N.; Kay, A.; Mueller, U.G.; Kaspari, M. Metabolism and the rise of fungus cultivation by ants. Am. Nat. 2014, 184, 364–373. [Google Scholar] [CrossRef]
- Ranger, C.M.; Biedermann, P.H.W.; Phuntumart, V.; Beligala, G.U.; Ghosh, S.; Palmquist, D.E.; Mueller, R.; Barnett, J.; Schultz, P.B.; Reding, M.E.; et al. Symbiont selection via alcohol benefits fungus farming by ambrosia beetles. Proc. Natl. Acad. Sci. USA 2018, 115, 4447–4452. [Google Scholar] [CrossRef]
- Rehan, S.M.; Toth, A.L. Climbing the social ladder: The molecular evolution of sociality. Trends Ecol. Evol. 2015, 30, 426–433. [Google Scholar] [CrossRef]
- Toth, A.L.; Varala, K.; Henshaw, M.T.; Rodriguez-Zas, S.L.; Hudson, M.E.; Robinson, G.E. Brain transcriptomic analysis in paper wasps identifies genes associated with behaviour across social insect lineages. Proc. Biol. Sci. 2010, 277, 2139–2148. [Google Scholar] [CrossRef]
- Toth, A.L.; Tooker, J.F.; Radhakrishnan, S.; Minard, R.; Henshaw, M.T.; Grozinger, C.M. Shared genes related to aggression, rather than chemical communication, are associated with reproductive dominance in paper wasps (Polistes metricus). BMC Genom. 2014, 15, 75. [Google Scholar] [CrossRef] [PubMed]
- Mikheyev, A.S.; Linksvayer, T.A. Genes associated with ant social behavior show distinct transcriptional and evolutionary patterns. eLife 2015, 4, e04775. [Google Scholar] [CrossRef] [PubMed]
- Woodard, S.H.; Fischman, B.J.; Venkat, A.; Hudson, M.E.; Varala, K.; Cameron, S.A.; Clark, A.G.; Robinson, G.E. Genes involved in convergent evolution of eusociality in bees. Proc. Natl. Acad. Sci. USA 2011, 108, 7472–7477. [Google Scholar] [CrossRef] [PubMed]
- Harpur, B.A.; Kent, C.F.; Molodtsova, D.; Lebon, J.M.D.; Alqarni, A.S.; Owayss, A.A.; Zayed, A. Population genomics of the honey bee reveals strong signatures of positive selection on worker traits. Proc. Natl. Acad. Sci. USA 2014, 111, 2614–2619. [Google Scholar] [CrossRef] [PubMed]
- Johnson, B.R.; Tsutsui, N.D. Taxonomically restricted genes are associated with the evolution of sociality in the honey bee. BMC Genom. 2011, 12, 164. [Google Scholar] [CrossRef] [PubMed]
- Sumner, S. The importance of genomic novelty in social evolution. Mol. Ecol. 2014, 23, 26–28. [Google Scholar] [CrossRef] [PubMed]
- Weitekamp, C.A.; Libbrecht, R.; Keller, L. Genetics and evolution of social behavior in insects. Annu. Rev. Genet. 2017, 51, 219–239. [Google Scholar] [CrossRef] [PubMed]
- Toth, A.L.; Rehan, S.M. Molecular evolution of insect sociality: An eco-evo-devo perspective. Annu. Rev. Entomol. 2017, 62, 419–442. [Google Scholar] [CrossRef] [PubMed]
- Harrison, M.C.; Jongepier, E.; Robertson, H.M.; Arning, N.; Bitard-Feildel, T.; Chao, H.; Childers, C.P. Hemimetabolous genomes reveal molecular basis of termite eusociality. Nat. Ecol. Evol. 2018, 2, 557–566. [Google Scholar] [CrossRef]
- Nygaard, S.; Hu, H.; Li, C.; Schiott, M.; Chen, Z.; Yang, Z.; Xie, Q.; Ma, C.; Deng, Y.; Dikow, R.B.; Rabeling, C.; et al. Reciprocal genomic evolution in the ant-fungus agricultural symbiosis. Nat. Commun. 2016, 7, 12233. [Google Scholar] [CrossRef]
- Favreau, E.; Martínez-Ruiz, C.; Rodrigues Santiago, L.; Hammond, R.L.; Wurm, Y. Genes and genomic processes underpinning the social lives of ants. Curr. Opin. Insect Sci. 2018, 25, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Lutzoni, F.; Pagel, M. Accelerated evolution as a consequence of transitions to mutualism. Proc. Natl. Acad. Sci. USA 1997, 94, 11422–11427. [Google Scholar] [CrossRef] [PubMed]
- Rubin, B.E.R.; Moreau, C.S. Comparative genomics reveals convergent rates of evolution in ant–plant mutualisms. Nat. Commun. 2016, 7, 12679. [Google Scholar] [CrossRef] [PubMed]
- Stouthamer, R.; Rugman-Jones, P.; Thu, P.Q.; Eskalen, A.; Thibault, T.; Hulcr, J.; Wang, L.-J.; Jordal, B.H.; Chen, C.-Y.; Cooperband, M.; et al. Tracing the origin of a cryptic invader: Phylogeography of the Euwallacea fornicatus (Coleoptera: Curculionidae: Scolytinae) species complex. Agric. For. Entomol. 2017, 19, 366–375. [Google Scholar] [CrossRef]
- Freeman, S.; Sharon, M.; Maymon, M.; Mendel, Z.; Protasov, A.; Aoki, T.; Eskalen, A.; O’Donnell, K. Fusarium euwallaceae sp. Nov.—A symbiotic fungus of Euwallacea sp., an invasive ambrosia beetle in Israel and California. Mycologia 2013, 105, 1595–1606. [Google Scholar] [CrossRef] [PubMed]
- Mayfield, A.E.I.; Pena, J.E.; Crane, J.H.; Smith, J.A.; Branch, C.L.; Ottoson, E.D.; Hughes, M. Ability of the redbay ambrosia beetle (Coleoptera: Curculionidae: Scolytinae) to bore into young avocado (Lauraceae) plants and transmit the laurel wilt pathogen (Raffaelea sp.). Fla. Entomol. 2008, 91, 485–487. [Google Scholar] [CrossRef]
- Kasson, M.T.; O’Donnell, K.; Rooney, A.P.; Sink, S.; Ploetz, R.C.; Ploetz, J.N.; Konkol, J.L.; Carrillo, D.; Freeman, S.; Mendel, Z.; et al. An inordinate fondness for Fusarium: Phylogenetic diversity of fusaria cultivated by ambrosia beetles in the genus Euwallacea on avocado and other plant hosts. Fungal Genet. Biol. 2013, 56, 147–157. [Google Scholar] [CrossRef]
- O’Donnell, K.; Libeskind-Hadas, R.; Hulcr, J.; Bateman, C.; Kasson, M.T.; Ploetz, R.C.; Konkol, J.L.; Ploetz, J.N.; Carrillo, D.; Campbell, A.; et al. Invasive asian Fusarium—Euwallacea ambrosia beetle mutualists pose a serious threat to forests, urban landscapes and the avocado industry. Phytoparasitica 2016, 44, 435–442. [Google Scholar] [CrossRef]
- Carrillo, D.; Cruz, L.F.; Kendra, P.E.; Narvaez, T.I.; Montgomery, W.S.; Monterroso, A.; De Grave, C.; Cooperband, M.F. Distribution, pest status and fungal associates of Euwallacea nr. fornicatus in florida avocado groves. Insects 2016, 7, 55. [Google Scholar]
- Carrillo, D.; Duncan, R.E.; Peña, J.E. Ambrosia beetles (Coleoptera: Curculionidae: Scolytinae) that breed in avocado wood in florida. Fla. Entomol. 2012, 95, 573–579. [Google Scholar] [CrossRef]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [PubMed]
- Grigoriev, I.V.; Cullen, D.; Goodwin, S.B.; Hibbett, D.; Jeffries, T.W.; Kubicek, C.P.; Kuske, C.; Magnuson, J.K.; Martin, F.; Spatafora, J.W.; et al. Fueling the future with fungal genomics. Mycology 2011, 2, 192–209. [Google Scholar]
- Grigoriev, I.V.; Nikitin, R.; Haridas, S.; Kuo, A.; Ohm, R.; Otillar, R.; Riley, R.; Salamov, A.; Zhao, X.; Korzeniewski, F.; et al. Mycocosm portal: Gearing up for 1000 fungal genomes. Nucleic Acids Res. 2014, 42, D699–D704. [Google Scholar] [CrossRef] [PubMed]
- Ibarra-Laclette, E.; Sánchez-Rangel, D.; Hernández-Domínguez, E.; Pérez-Torres, C.A.; Ortiz-Castro, R.; Villafán, E.; Alonso-Sánchez, A.; Rodríguez-Hass, B.; López-Buenfil, A.; García-Avila, C.; et al. Draft genome sequence of the phytopathogenic fungus Fusarium euwallaceae, the causal agent of fusarium dieback. Genome Announc. 2017, 5, e00881–e00817. [Google Scholar] [CrossRef] [PubMed]
- Vanderpool, D.; Bracewell, R.R.; McCutcheon, J.P. Know your farmer: Ancient origins and multiple independent domestications of ambrosia beetle fungal cultivars. Mol. Ecol. 2018, 27, 2077–2094. [Google Scholar] [CrossRef] [PubMed]
- Wingfield, B.D.; Barnes, I.; Wilhelm de Beer, Z.; De Vos, L.; Duong, T.A.; Kanzi, A.M.; Naidoo, K.; Nguyen, H.D.T.; Santana, Q.C.; Sayari, M.; et al. IMA genome-f 5: Draft genome sequences of Ceratocystis eucalypticola, Chrysoporthe cubensis, C. deuterocubensis, Davidsoniella virescens, Fusarium temperatum, Graphilbum fragrans, Penicillium nordicum, and Thielaviopsis musarum. IMA Fungus 2015, 6, 493–506. [Google Scholar] [CrossRef] [PubMed]
- Wingfield, B.D.; Ades, P.K.; Al-Naemi, F.A.; Beirn, L.A.; Bihon, W.; Crouch, J.A.; de Beer, Z.W.; De Vos, L.; Duong, T.A.; Fields, C.J.; et al. IMA genome-f 4: Draft genome sequences of Chrysoporthe austroafricana, Diplodia scrobiculata, Fusarium nygamai, Leptographium lundbergii, Limonomyces culmigenus, Stagonosporopsis tanaceti, and Thielaviopsis punctulata. IMA Fungus 2015, 6, 233–248. [Google Scholar] [CrossRef]
- Endoh, R.; Suzuki, M.; Okada, G.; Takeuchi, Y.; Futai, K. Fungus symbionts colonizing the galleries of the ambrosia beetle Platypus quercivorus. Microb. Ecol. 2011, 62, 106–120. [Google Scholar] [CrossRef]
- De Marco, L.; Epis, S.; Capone, A.; Martin, E.; Bozic, J.; Crotti, E.; Ricci, I.; Sassera, D. The genomes of four Meyerozyma caribbica isolates and novel insights into the Meyerozyma guilliermondii species complex. Genes Genomes Genet. 2018, 8, 755–759. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef]
- Shu, S.; Chen, B.; Zhou, M.; Zhao, X.; Xia, H.; Wang, M. De novo sequencing and transcriptome analysis of Wolfiporia cocos to reveal genes related to biosynthesis of triterpenoids. PLoS ONE 2013, 8, e71350. [Google Scholar] [CrossRef] [PubMed]
- Evans, T.; Loose, M. ALIGNWISE: A tool for identifying protein-coding sequence and correcting frame-shifts. BMC Bioinform. 2015, 16, 376. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed]
- Birney, E.; Clamp, M.; Durbin, R. Genewise and genomewise. Genome Res. 2004, 14, 988–995. [Google Scholar] [CrossRef] [PubMed]
- Poelchau, M.; Childers, C.; Moore, G.; Tsavatapalli, V.; Evans, J.; Lee, C.-Y.; Lin, H.; Lin, J.-W.; Hackett, K. The i5k workspace@NAL—Enabling genomic data access, visualization and curation of arthropod genomes. Nucleic Acids Res. 2015, 43, D714–D719. [Google Scholar] [CrossRef] [PubMed]
- Meyer, J.M.; Markov, G.V.; Baskaran, P.; Herrmann, M.; Sommer, R.J.; Rödelsperger, C. Draft genome of the scarab beetle Oryctes borbonicus on La Réunion Island. Genome Biol. Evol. 2016, 8, 2093–2105. [Google Scholar] [CrossRef]
- Fallon, T.R.; Lower, S.E.; Chang, C.-H.; Bessho-Uehara, M.; Martin, G.J.; Bewick, A.J.; Behringer, M.; Debat, H.J.; Wong, I.; Day, J.C.; et al. Firefly genomes illuminate parallel origins of bioluminescence in beetles. eLife 2018, 7, e36495. [Google Scholar] [CrossRef]
- McKenna, D.D.; Scully, E.D.; Pauchet, Y.; Hoover, K.; Kirsch, R.; Geib, S.M.; Mitchell, R.F.; Waterhouse, R.M.; Ahn, S.-J.; Arsala, D.; et al. Genome of the asian longhorned beetle (Anoplophora glabripennis), a globally significant invasive species, reveals key functional and evolutionary innovations at the beetle-plant interface. Genome Biol. 2016, 17, 227. [Google Scholar] [CrossRef]
- Schoville, S.D.; Chen, Y.H.; Andersson, M.N.; Benoit, J.B.; Bhandari, A.; Bowsher, J.H.; Brevik, K.; Cappelle, K.; Chen, M.-J.M.; Childers, A.K.; et al. A model species for agricultural pest genomics: The genome of the colorado potato beetle, Leptinotarsa decemlineata (Coleoptera: Chrysomelidae). Sci. Rep. 2018, 8, 1931. [Google Scholar] [CrossRef]
- Keeling, C.I.; Yuen, M.M.; Liao, N.Y.; Roderick Docking, T.; Chan, S.K.; Taylor, G.A.; Palmquist, D.L.; Jackman, S.D.; Nguyen, A.; Li, M.; et al. Draft genome of the mountain pine beetle, Dendroctonus ponderosae hopkins, a major forest pest. Genome Biol. 2013, 14, R27. [Google Scholar] [CrossRef] [PubMed]
- Tribolium Genome Sequencing Consortium. The genome of the model beetle and pest Tribolium castaneum. Nature 2008, 452, 949. [Google Scholar] [CrossRef] [PubMed]
- The Honeybee Genome Sequencing Consortium. Insights into social insects from the genome of the honeybee Apis mellifera. Nature 2006, 443, 931. [Google Scholar] [CrossRef]
- Suen, G.; Teiling, C.; Li, L.; Holt, C.; Abouheif, E.; Bornberg-Bauer, E.; Bouffard, P.; Caldera, E.J.; Cash, E.; Cavanaugh, A.; et al. The genome sequence of the leaf-cutter ant Atta cephalotes reveals insights into its obligate symbiotic lifestyle. PLoS Genet. 2011, 7, e1002007. [Google Scholar] [CrossRef]
- Oxley, P.R.; Ji, L.; Fetter-Pruneda, I.; McKenzie, S.K.; Li, C.; Hu, H.; Zhang, G.; Kronauer, D.J. The genome of the clonal raider ant Cerapachys biroi. Curr. Biol. 2014, 24, 451–458. [Google Scholar] [CrossRef] [PubMed]
- Standage, D.S.; Berens, A.J.; Glastad, K.M.; Severin, A.J.; Brendel, V.P.; Toth, A.L. Genome, transcriptome and methylome sequencing of a primitively eusocial wasp reveal a greatly reduced DNA methylation system in a social insect. Mol. Ecol. 2016, 25, 1769–1784. [Google Scholar] [CrossRef]
- Nygaard, S.; Zhang, G.; Schiott, M.; Li, C.; Wurm, Y.; Hu, H.; Zhou, J.; Ji, L.; Qiu, F.; Rasmussen, M.; et al. The genome of the leaf-cutting ant Acromyrmex echinatior suggests key adaptations to advanced social life and fungus farming. Genome Res. 2011, 21, 1339–1348. [Google Scholar] [CrossRef]
- Terrapon, N.; Li, C.; Robertson, H.M.; Ji, L.; Meng, X.; Booth, W.; Chen, Z.; Childers, C.P.; Glastad, K.M.; Gokhale, K.; et al. Molecular traces of alternative social organization in a termite genome. Nat. Commun. 2014, 5, 3636. [Google Scholar] [CrossRef]
- Adams, M.D.; Celniker, S.E.; Holt, R.A.; Evans, C.A.; Gocayne, J.D.; Amanatides, P.G.; Scherer, S.E.; Li, P.W.; Hoskins, R.A.; Galle, R.F.; et al. The genome sequence of Drosophila melanogaster. Science 2000, 287, 2185. [Google Scholar] [CrossRef]
- Jones, P.; Binns, D.; Chang, H.Y.; Fraser, M.; Li, W.; McAnulla, C.; McWilliam, H.; Maslen, J.; Mitchell, A.; Nuka, G.; et al. Interproscan 5: Genome-scale protein function classification. Bioinformatics 2014, 30, 1236–1240. [Google Scholar] [CrossRef]
- Finn, R.D.; Attwood, T.K.; Babbitt, P.C.; Bateman, A.; Bork, P.; Bridge, A.J.; Chang, H.-Y.; Dosztányi, Z.; El-Gebali, S.; Fraser, M.; et al. Interpro in 2017—beyond protein family and domain annotations. Nucleic Acids Res. 2017, 45, D190–D199. [Google Scholar] [CrossRef] [PubMed]
- Finn, R.D.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Mistry, J.; Mitchell, A.L.; Potter, S.C.; Punta, M.; Qureshi, M.; Sangrador-Vegas, A.; et al. The pFam protein families database: Towards a more sustainable future. Nucleic Acids Res. 2016, 44, D279–D285. [Google Scholar] [CrossRef] [PubMed]
- Punta, M.; Coggill, P.C.; Eberhardt, R.Y.; Mistry, J.; Tate, J.; Boursnell, C.; Pang, N.; Forslund, K.; Ceric, G.; Clements, J.; et al. The pFam protein families database. Nucleic Acids Res. 2012, 40, D290–D301. [Google Scholar] [CrossRef] [PubMed]
- Haft, D.H.; Selengut, J.D.; Richter, R.A.; Harkins, D.; Basu, M.K.; Beck, E. TIGRFAMS and genome properties in 2013. Nucleic Acids Res. 2013, 41, D387–D395. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.H.; Nikolskaya, A.; Huang, H.; Yeh, L.S.; Natale, D.A.; Vinayaka, C.R.; Hu, Z.Z.; Mazumder, R.; Kumar, S.; Kourtesis, P.; et al. PIRSF: Family classification system at the protein information resource. Nucleic Acids Res. 2004, 32, D112–D114. [Google Scholar] [CrossRef] [PubMed]
- de Lima Morais, D.A.; Fang, H.; Rackham, O.J.L.; Wilson, D.; Pethica, R.; Chothia, C.; Gough, J. SUPERFAMILY 1.75 including a domain-centric gene ontology method. Nucleic Acids Res. 2011, 39, D427–D434. [Google Scholar] [CrossRef] [PubMed]
- Stanke, M.; Tzvetkova, A.; Morgenstern, B. AUGUSTUS at EGASP: Using est, protein and genomic alignments for improved gene prediction in the human genome. Genome Biol. 2006, 7. [Google Scholar] [CrossRef]
- Stanke, M.; Diekhans, M.; Baertsch, R.; Haussler, D. Using native and syntenically mapped cDNA alignments to improve de novo gene finding. Bioinformatics 2008, 24, 637–644. [Google Scholar] [CrossRef]
- Cantarel, B.L.; Korf, I.; Robb, S.M.C.; Parra, G.; Ross, E.; Moore, B.; Holt, C.; Alvarado, A.S.; Yandell, M. MAKER: An easy-to-use annotation pipeline designed for emerging model organism genomes. Genome Res. 2008, 18. [Google Scholar] [CrossRef]
- Campbell, M.S.; Holt, C.; Moore, B.; Yandell, M. Genome annotation and curation using MAKER and MAKER-P. Curr. Protoc. Bioinform. 2014, 48. [Google Scholar] [CrossRef]
- Sánchez-Rangel, D.; Hernández-Domínguez, E.E.; Pérez-Torres, C.; Ortiz-Castro, R.; Villafán, E.; Rodríguez-Haas, B.; Alonso-Sánchez, A.; López-Buenfil, A.; Carrillo-Ortiz, N.; Hernández-Ramos, L.; et al. Environmental pH modulates genetic responses related to pathogenicity in the fungus Fusarium sp. associated with KSHB Euwallaceae sp. Near fornicatus. BMC Genom. 2018, 19, 721. [Google Scholar]
- Contreras-Moreira, B.; Vinuesa, P. GET_HOMOLOGUES, a versatile software package for scalable and robust microbial pangenome analysis. Appl. Environ. Microbiol. 2013, 79, 7696–7701. [Google Scholar] [CrossRef] [PubMed]
- Contreras-Moreira, B.; Cantalapiedra, C.P.; Garcia-Pereira, M.J.; Gordon, S.P.; Vogel, J.P.; Igartua, E.; Casas, A.M.; Vinuesa, P. Analysis of plant pan-genomes and transcriptomes with GET_HOMOLOGUES-EST, a clustering solution for sequences of the same species. Front. Plant Sci. 2017, 8, 184. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Stoeckert, C.J., Jr.; Roos, D.S. ORTHOMCL: Identification of ortholog groups for Eukaryotic Genomes. Genome Res. 2003, 13, 2178–2189. [Google Scholar] [CrossRef] [PubMed]
- Enright, A.J.; Van Dongen, S.; Ouzounis, C.A. An efficient algorithm for large-scale detection of protein families. Nucleic Acids Res. 2002, 30, 1575–1584. [Google Scholar] [CrossRef] [PubMed]
- Yamada, K.; Katoh, K.; Tomii, K. 1P267 application of novel amino acid substitution matrix, MIQS, to the mafft multiple sequence aligner (22c. Bioinformatics: Comparative genomics, poster, the 52nd annual meeting of the biophysical society of japan(bsj2014)). Seibutsu Butsuri 2014, 54, S185. [Google Scholar] [CrossRef]
- Suyama, M.; Torrents, D.; Bork, P. PAL2NAL: Robust conversion of protein sequence alignments into the corresponding codon alignments. Nucleic Acids Res. 2006, 34, W609–W612. [Google Scholar] [CrossRef] [PubMed]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FASTTREE 2—Approximately maximum-likelihood trees for large alignments. PLoS ONE 2010, 5, e9490. [Google Scholar] [CrossRef]
- Pond, S.L.; Frost, S.D.; Grossman, Z.; Gravenor, M.B.; Richman, D.D.; Brown, A.J. Adaptation to different human populations by HIV-1 revealed by codon-based analyses. PLoS Comput. Biol. 2006, 2, e62. [Google Scholar]
- Smith, M.D.; Wertheim, J.O.; Weaver, S.; Murrell, B.; Scheffler, K.; Kosakovsky Pond, S.L. Less is more: An adaptive branch-site random effects model for efficient detection of episodic diversifying selection. Mol. Biol. Evol. 2015, 32, 1342–1353. [Google Scholar] [CrossRef]
- Guindon, S.; Dufayard, J.F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef]
- Lefort, V.; Longueville, J.E.; Gascuel, O. SMS: Smart model selection in PhyML. Mol. Biol. Evol. 2017, 34, 2422–2424. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z. PAML 4: Phylogenetic analysis by maximum likelihood. Mol. Biol. Evol. 2007, 24, 1586–1591. [Google Scholar] [CrossRef] [PubMed]
- Misof, B.; Liu, S.; Meusemann, K.; Peters, R.S.; Donath, A.; Mayer, C.; Frandsen, P.B.; Ware, J.; Flouri, T.; Beutel, R.G.; et al. Phylogenomics resolves the timing and pattern of insect evolution. Science 2014, 346, 763–767. [Google Scholar] [CrossRef] [PubMed]
- Rambaut, A.; Suchard, M.A.; Xie, D.; Drummond, A.J. TRACER v1.6. Available online: http://tree.bio.ed.ac.uk/software/tracer/ (accessed on 14 October 2018).
- Levy Karin, E.; Wicke, S.; Pupko, T.; Mayrose, I. An integrated model of phenotypic trait changes and site-specific sequence evolution. Syst. Biol. 2017, 66, 917–933. [Google Scholar] [CrossRef] [PubMed]
- Levy Karin, E.; Ashkenazy, H.; Wicke, S.; Pupko, T.; Mayrose, I. TraitRateProp: A web server for the detection of trait-dependent evolutionary rate shifts in sequence sites. Nucleic Acids Res. 2017, 45, W260–W264. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [PubMed]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. EDGER: A bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B Methodol. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Li, Y.; Dharkar, P.; Han, T.H.; Serpe, M.; Lee, C.H.; Mayer, M.L. Novel functional properties of Drosophila CNS glutamate receptors. Neuron 2016, 92, 1036–1048. [Google Scholar] [CrossRef] [PubMed]
- Funada, M.; Hara, H.; Sasagawa, H.; Kitagawa, Y.; Kadowaki, T. A honey bee dsCAM family member, abscam, is a brain-specific cell adhesion molecule with the neurite outgrowth activity which influences neuronal wiring during development. Eur. J. Neurosci. 2007, 25, 168–180. [Google Scholar] [CrossRef] [PubMed]
- Kanamori, Y.; Saito, A.; Hagiwara-Komoda, Y.; Tanaka, D.; Mitsumasu, K.; Kikuta, S.; Watanabe, M.; Cornette, R.; Kikawada, T.; Okuda, T. The trehalose transporter 1 gene sequence is conserved in insects and encodes proteins with different kinetic properties involved in trehalose import into peripheral tissues. Insect Biochem. Mol. Biol. 2010, 40, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.W.; Zhang, X.X.; Chen, J.Y.; Lu, M.X.; Gong, W.R.; Du, Y.Z. Characterization of three heat shock protein 70 genes from Liriomyza trifolii and expression during thermal stress and insect development. Bull. Entomol. Res. 2018. [Google Scholar] [CrossRef] [PubMed]
- Iwaki, D.D.; Johansen, K.A.; Singer, J.B.; Lengyel, J.A. drumstick, bowl, and lines are required for patterning and cell rearrangement in the Drosophila embryonic hindgut. Dev. Biol. 2001, 240, 611–626. [Google Scholar] [CrossRef] [PubMed]
- Unschuld, P.G.; Dächsel, J.; Darios, F.; Kohlmann, A.; Casademunt, E.; Lehmann-Horn, K.; Dichgans, M.; Ruberg, M.; Brice, A.; Gasser, T.; et al. Parkin modulates gene expression in control and ceramide-treated PC12 cells. Mol. Biol. Rep. 2006, 33, 13–32. [Google Scholar] [CrossRef] [PubMed]
- Mi, W.; Zhang, Y.; Lyu, J.; Wang, X.; Tong, Q.; Peng, D.; Xue, Y.; Tencer, A.H.; Wen, H.; Li, W.; et al. The ZZ-type zinc finger of ZZZ3 modulates the atac complex-mediated histone acetylation and gene activation. Nat. Commun. 2018, 9, 3759. [Google Scholar] [CrossRef]
- Seachrist, J.L.; Ferguson, S.S.G. Regulation of G protein-coupled receptor endocytosis and trafficking by rab gtpases. Life Sci. 2003, 74, 225–235. [Google Scholar] [CrossRef]
- Miller, K.G.; Emerson, M.D.; McManus, J.R.; Rand, J.B. RIC-8 (synembryn): A novel conserved protein that is required for G(α)alpha signaling in the C. elegans nervous system. Neuron 2000, 27, 289–299. [Google Scholar] [CrossRef]
- Sopko, R.; Perrimon, N. Receptor tyrosine kinases in Drosophila development. Cold Spring Harb. Perspect. Biol. 2013, 5. [Google Scholar] [CrossRef]
- Krymskaya, V.P. Tumour suppressors hamartin and tuberin: Intracellular signalling. Cell. Signal. 2003, 15, 729–739. [Google Scholar] [CrossRef]
- Nowling, R.J.; Abrudan, J.L.; Shoue, D.A.; Abdul-Wahid, B.; Wadsworth, M.; Stayback, G.; Collins, F.H.; McDowell, M.A.; Izaguirre, J.A. Identification of novel arthropod vector G protein-coupled receptors. Parasites Vectors 2013, 6, 150. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, W.; Nakanishi, H.; Miyoshi, J.; Mandai, K.; Ishizaki, H.; Tanaka, M.; Togawa, A.; Takahashi, K.; Nishioka, H.; Yoshida, H.; et al. Afadin: A key molecule essential for structural organization of cell-cell junctions of polarized epithelia during embryogenesis. J. Cell Biol. 1999, 146, 1117. [Google Scholar] [CrossRef] [PubMed]
- Ito, Y.; Yasuda, A.; Sonobe, H. Synthesis and phosphorylation of ecdysteroids during ovarian development in the silkworm, Bombyx mori. Zool. Sci. 2008, 25, 721–727. [Google Scholar] [CrossRef] [PubMed]
- Katzemich, A.; West, R.J.H.; Fukuzawa, A.; Sweeney, S.T.; Gautel, M.; Sparrow, J.; Bullard, B. Binding partners of the kinase domains in Drosophila obscurin and their effect on the structure of the flight muscle. J. Cell Sci. 2015, 128, 3386–3397. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, A.; Hasselbach, L.; Echtermeyer, F.; Jyotsana, N.; Theilmeier, G.; Herzog, C. Fibulin-6 regulates pro-fibrotic TGF-β responses in neonatal mouse ventricular cardiac fibroblasts. Sci. Rep. 2017, 7, 42725. [Google Scholar] [CrossRef]
- Morel, V.; Lepicard, S.; Rey, A.N.; Parmentier, M.L.; Schaeffer, L. Drosophila nesprin-1 controls glutamate receptor density at neuromuscular junctions. Cell. Mol. Life Sci. 2014, 71, 3363–3379. [Google Scholar] [CrossRef]
- Brose, K.; Bland, K.S.; Wang, K.H.; Arnott, D.; Henzel, W.; Goodman, C.S.; Tessier-Lavigne, M.; Kidd, T. Slit proteins bind robo receptors and have an evolutionarily conserved role in repulsive axon guidance. Cell 1999, 96, 795–806. [Google Scholar] [CrossRef]
- Xing, G.; Li, M.; Sun, Y.; Rui, M.; Zhuang, Y.; Lv, H.; Han, J.; Jia, Z.; Xie, W. Neurexin–neuroligin 1 regulates synaptic morphology and functions via the WAVE regulatory complex in Drosophila neuromuscular junction. eLife 2018, 7, e30457. [Google Scholar] [CrossRef]
- Klaric, T.; Lardelli, M.; Key, B.; Koblar, S.; Lewis, M. Activity-dependent expression of neuronal PAS domain-containing protein 4 (npas4a) in the developing zebrafish brain. Front. Neuroanat. 2014, 8, 148. [Google Scholar]
- Kubli, E. Sexual behaviour: A receptor for sex control in Drosophila females. Curr. Biol. 2008, 18, R210–R212. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Yapici, N.; Kim, Y.-J.; Ribeiro, C.; Dickson, B.J. A receptor that mediates the post-mating switch in Drosophila reproductive behaviour. Nature 2007, 451, 33. [Google Scholar] [CrossRef] [PubMed]
- Barchuk, A.R.; dos Santos, G.D.; Dias Caneschi, R.; de Paula Junior, D.E.; Moda, L.M.R. The ontogenetic saga of a social brain. Apidologie 2018, 49, 32–48. [Google Scholar] [CrossRef]
- Delaux, P.M.; Varala, K.; Edger, P.P.; Coruzzi, G.M.; Pires, J.C.; Ane, J.M. Comparative phylogenomics uncovers the impact of symbiotic associations on host genome evolution. PLoS Genet. 2014, 10, e1004487. [Google Scholar] [CrossRef] [PubMed]
- De Fine Licht, H.H.; Boomsma, J.J.; Tunlid, A. Symbiotic adaptations in the fungal cultivar of leaf-cutting ants. Nat. Commun. 2014, 5, 5675. [Google Scholar] [CrossRef] [PubMed]
- Sudakaran, S.; Kost, C.; Kaltenpoth, M. Symbiont acquisition and replacement as a source of ecological innovation. Trends Microbiol. 2017, 25, 375–390. [Google Scholar] [CrossRef] [PubMed]
- Ayres, M.P.; Wilkens, R.T.; Ruel, J.J.; Lombardero, M.J.; Vallery, E. Nitrogen budgets of phloem-feeding bark beetles with and without symbiotic fungi. Ecology 2000, 81, 2198–2210. [Google Scholar] [CrossRef]
- Norris, D.M.; Baker, J.M.; Chu, H.M. Symbiontic interrelationships between microbes and ambrosia beetles. III. Ergosterol as the source of sterol to the insect. Ann. Entomol. Soc. Am. 1969, 62, 413–414. [Google Scholar] [CrossRef]
- Rider, M.H.; Bertrand, L.; Vertommen, D.; Michels, Paul A.; Rousseau, Guy G.; Hue, L. 6-phosphofructo-2-kinase/fructose-2,6-bisphosphatase: Head-to-head with a bifunctional enzyme that controls glycolysis. Biochem. J. 2004, 381, 561. [Google Scholar] [CrossRef] [PubMed]
- Yasugi, T.; Yamada, T.; Nishimura, T. Adaptation to dietary conditions by trehalose metabolism in Drosophila. Sci. Rep. 2017, 7, 1619. [Google Scholar] [CrossRef]
- Du, X.H.; Gray, P.M. Evolutionary Gem: Evolution of lactase persistence. West. Undergr. Res. J. 2017, 8, 1–3. [Google Scholar] [CrossRef][Green Version]
- Liebert, A.; Lopez, S.; Jones, B.L.; Montalva, N.; Gerbault, P.; Lau, W.; Thomas, M.G.; Bradman, N.; Maniatis, N.; Swallow, D.M. World-wide distributions of lactase persistence alleles and the complex effects of recombination and selection. Hum. Genet. 2017, 136, 1445–1453. [Google Scholar] [CrossRef] [PubMed]
- Segurel, L.; Bon, C. On the evolution of lactase persistence in humans. Annu. Rev. Genom. Hum. Genet. 2017, 18, 297–319. [Google Scholar] [CrossRef] [PubMed]
- Luca, F.; Perry, G.H.; Rienzo, A.D. Evolutionary adaptations to dietary changes. Annu. Rev. Nutr. 2010, 30, 291–314. [Google Scholar] [CrossRef] [PubMed]
- Kapheim, K.M.; Pan, H.; Li, C.; Salzberg, S.L.; Puiu, D.; Magoc, T.; Robertson, H.M.; Hudson, M.E.; Venkat, A.; Fischman, B.J.; et al. Social evolution. Genomic signatures of evolutionary transitions from solitary to group living. Science 2015, 348, 1139–1143. [Google Scholar] [CrossRef] [PubMed]
- Shell, W.A.; Rehan, S.M. Behavioral and genetic mechanisms of social evolution: Insights from incipiently and facultatively social bees. Apidologie 2018, 49, 13–30. [Google Scholar] [CrossRef]
- Rehan, S.M.; Berens, A.J.; Toth, A.L. At the brink of eusociality: Transcriptomic correlates of worker behaviour in a small carpenter bee. BMC Evol. Biol. 2014, 14, 260. [Google Scholar] [CrossRef]
- Cande, J.; Prud’homme, B.; Gompel, N. Smells like evolution: The role of chemoreceptor evolution in behavioral change. Curr. Opin. Neurobiol. 2013, 23, 152–158. [Google Scholar] [CrossRef]
- Kreher, S.A.; Mathew, D.; Kim, J.; Carlson, J.R. Translation of sensory input into behavioral output via an olfactory system. Neuron 2008, 59, 110–124. [Google Scholar] [CrossRef]
- Avila, F.W.; Mattei, A.L.; Wolfner, M.F. Sex peptide receptor is required for the release of stored sperm by mated Drosophila melanogaster females. J. Insect Physiol. 2015, 76, 1–6. [Google Scholar] [CrossRef]
- Oldroyd, B.P.; Beekman, M. Effects of selection for honey bee worker reproduction on foraging traits. PLoS Biol. 2008, 6, e56. [Google Scholar] [CrossRef] [PubMed]
- Marinissen, M.J.; Gutkind, J.S. G-protein-coupled receptors and signaling networks: Emerging paradigms. Trends Pharmacol. Sci. 2001, 22, 368–376. [Google Scholar] [CrossRef]
- Simola, D.F.; Wissler, L.; Donahue, G.; Waterhouse, R.M.; Helmkampf, M.; Roux, J.; Nygaard, S.; Glastad, K.M.; Hagen, D.E.; Viljakainen, L.; et al. Social insect genomes exhibit dramatic evolution in gene composition and regulation while preserving regulatory features linked to sociality. Genome Res. 2013, 23, 1235–1247. [Google Scholar] [CrossRef] [PubMed]
- Cooperband, M.F.; Cossé, A.A.; Jones, T.H.; Carrillo, D.; Cleary, K.; Canlas, I.; Stouthamer, R. Pheromones of three ambrosia beetles in the Euwallacea fornicatus species complex: Ratios and preferences. PeerJ 2017, 5, e3957. [Google Scholar] [CrossRef] [PubMed]
- Beroza, M. Chemicals Controlling Insect Behavior; Elsevier Science: Amsterdam, The Netherlands, 2012. [Google Scholar]
- Funaro, C.F.; Böröczky, K.; Vargo, E.L.; Schal, C. Identification of a queen and king recognition pheromone in the subterranean termite Reticulitermes flavipes. Proc. Natl. Acad. Sci. USA 2018, 115, 3888–3893. [Google Scholar] [CrossRef] [PubMed]
- Yanay, C.; Morpurgo, N.; Linial, M. Evolution of insect proteomes: Insights into synapse organization and synaptic vesicle life cycle. Genome Biol. 2008, 9, R27. [Google Scholar] [CrossRef] [PubMed]
- Spangler, S.A.; Hoogenraad, C.C. Liprin-alpha proteins: Scaffold molecules for synapse maturation. Biochem. Soc. Trans. 2007, 35, 1278–1282. [Google Scholar] [CrossRef]
- Beaubien, F.; Raja, R.; Kennedy, T.E.; Fournier, A.E.; Cloutier, J.-F. Slitrk1 is localized to excitatory synapses and promotes their development. Sci. Rep. 2016, 6, 27343. [Google Scholar] [CrossRef]
- Garbe, J.C.; Yang, E.; Fristrom, J.W. Imp-l2: An essential secreted immunoglobulin family member implicated in neural and ectodermal development in Drosophila. Development 1993, 119, 1237–1250. [Google Scholar]
- Lichte, B.; Veh, R.W.; Meyer, H.E.; Kilimann, M.W. Amphiphysin, a novel protein associated with synaptic vesicles. EMBO J. 1992, 11, 2521–2530. [Google Scholar] [CrossRef]
- Shi, L.; Quick, M.; Zhao, Y.; Weinstein, H.; Javitch, J.A. The mechanism of a neurotransmitter: Sodium symporter--inward release of Na+ and substrate is triggered by substrate in a second binding site. Mol. Cell 2008, 30, 667–677. [Google Scholar] [CrossRef] [PubMed]
- Goulding, S.E.; zur Lage, P.; Jarman, A.P. Amos, a proneural gene for Drosophila olfactory sense organs that is regulated by lozenge. Neuron 2000, 25, 69–78. [Google Scholar] [CrossRef]
- Huang, M.-L.; Hsu, C.-H.; Chien, C.-T. The proneural gene amos promotes multiple dendritic neuron formation in the Drosophila peripheral nervous system. Neuron 2000, 25, 57–67. [Google Scholar] [CrossRef]
- Schweisguth, F.; Posakony, J.W. Suppressor of hairless, the Drosophila homolog of the mouse recombination signal-binding protein gene, controls sensory organ cell fates. Cell 1992, 69, 1199–1212. [Google Scholar] [CrossRef]
- Corthals, K.; Heukamp, A.S.; Kossen, R.; Grosshennig, I.; Hahn, N.; Gras, H.; Gopfert, M.C.; Heinrich, R.; Geurten, B.R.H. Neuroligins Nlg2 and Nlg4 affect social behavior in Drosophila melanogaster. Front. Psychiatry 2017, 8, 113. [Google Scholar] [CrossRef] [PubMed]
- Bracewell, R.R.; Six, D.L. Broadscale specificity in a bark beetle-fungal symbiosis: A spatio-temporal analysis of the mycangial fungi of the western pine beetle. Microb. Ecol. 2014, 68, 859–870. [Google Scholar] [CrossRef] [PubMed]
- Kostovcik, M.; Bateman, C.C.; Kolarik, M.; Stelinski, L.L.; Jordal, B.H.; Hulcr, J. The ambrosia symbiosis is specific in some species and promiscuous in others: Evidence from community pyrosequencing. ISME J. 2014, 9, 126. [Google Scholar] [CrossRef]
- Six, D.L.; Bentz, B.J. Temperature determines symbiont abundance in a multipartite bark beetle-fungus ectosymbiosis. Microb. Ecol. 2007, 54, 112–118. [Google Scholar] [CrossRef]
- Addison, A.L.; Powell, J.A.; Six, D.L.; Moore, M.; Bentz, B.J. The role of temperature variability in stabilizing the mountain pine beetle-fungus mutualism. J. Theor. Biol. 2013, 335, 40–50. [Google Scholar] [CrossRef]
- Freeman, S.; Sharon, M.; Dori-Bachash, M.; Maymon, M.; Belausov, E.; Maoz, Y.; Margalit, O.; Protasov, A.; Mendel, Z. Symbiotic association of three fungal species throughout the life cycle of the ambrosia beetle Euwallacea nr. fornicatus. Symbiosis 2016, 68, 115–128. [Google Scholar] [CrossRef]
- Seybold, S.J.; Tittiger, C. Biochemistry and molecular biology of de novo isoprenoid pheromone production in the scolytidae. Annu. Rev. Entomol. 2003, 48, 425–453. [Google Scholar] [CrossRef] [PubMed]
- Seybold, S.J.; Bohlmann, J.; Raffa, K.F. Biosynthesis of coniferophagous bark beetle pheromones and conifer isoprenoids: Evolutionary perspective and synthesis. Can. Entomol. 2012, 132, 697–753. [Google Scholar] [CrossRef]
- Blomquist, G.J.; Figueroa-Teran, R.; Aw, M.; Song, M.; Gorzalski, A.; Abbott, N.L.; Chang, E.; Tittiger, C. Pheromone production in bark beetles. Insect Biochem. Mol. Biol. 2010, 40, 699–712. [Google Scholar] [CrossRef] [PubMed]
- Kashiwagi, T.; Nakashima, T.; Tebayashi, S.-I.; Kim, C.-S. Determination of the absolute configuration of quercivorol, (1S, 4R)-p-menth-2-en-1-ol, an aggregation pheromone of the ambrosia beetle Platypus quercivorus (Coleoptera: Platypodidae). Biosci. Biotechnol. Biochem. 2006, 70, 2544–2546. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
| Euwallacea sp. near fornicatus | Xyleborus glabratus | |
|---|---|---|
| Total high-quality paired reads | 61,919,467 | 68,243,721 |
| Total number of assembled transcripts | 248,739 | 150,163 |
| N50 (bp) | 1636 | 1261 |
| Total assembled bases | 221,342,598 | 110,621,067 |
| Average contig length (bp) | 889.86 | 736.67 |
| Total number of predicted unigenes | 68,490 | 46,814 |
| InterproScan5 annotated unigenes | 56,315 | 34,612 |
| BLASTp annotated unigenes | 55,115 | 37,188 |
| Cluster ID | Association with | BLASTp Annotation | Ortholog Group |
|---|---|---|---|
| 199572_KRT80184 | Mutualism | methylosome subunit pICln | α |
| 199849_KRT80461 | Mutualism | Rab-protein 6 (Rab6) | α |
| 200376_KRT80988 | Mutualism | ribosomal RNA small subunit methyltransferase NEP1 | α |
| 203911_KRT84523 | Mutualism | phosphatidylethanolamine-binding protein homolog F40A3.3 | α |
| 198969_KRT79581 | Both | ribosome biogenesis regulatory protein homolog | α |
| 201287_KRT81899 | Sociality | ubiquitin carboxyl-terminal hydrolase 30 | α |
| 203022_KRT83634 | Sociality | phosphoacetylglucosamine mutase-like | α |
| 180851_KRT82102 | Both | NADH dehydrogenase (ubiquinone) 1-alpha subcomplex 9, 39kDa (Ndufa9) | β |
| 181368_KRT82619 | Both | Bax inhibitor 1 (BaxI1) | β |
| 182416_KRT83667 | Both | vesicle transport through interaction with t-SNAREs homolog 1A | β |
| 182600_KRT83851 | Both | UDP-N-acetylglucosamine-dolichyl-phosphate N-acetylglucosaminephosphotransferase | β |
| 183558_KRT84809 | Sociality | ribosome-recycling factor, mitochondrial | β |
| 185607_KRT86858 | Sociality | malate dehydrogenase, mitochondrial | β |
| 176873_KRT78124 | Mutualism | poly (ADP-ribose) glycohydrolase ARH3 | β |
| 177777_KRT79028 | Mutualism | DNA replication complex GINS protein SLD5 | β |
| 181622_KRT82873 | Mutualism | ubiquitin domain-containing protein 1/2 | β |
| 185046_KRT86297 | Mutualism | MFS-type transporter C6orf192 homolog | β |
| 100143_KRT79886 | Both | cysteine-rich with EGF-like domain protein 2 | γ |
| 101937_KRT81680 | Both | Cornichon protein | γ |
| 103130_KRT82873 | Both | ubiquitin domain-containing protein 1 | γ |
| 99990_KRT79733 | Both | suppressor of hairless protein | γ |
| 100907_KRT80650 | Both | transmembrane protein 127 | γ |
| 99195_KRT78938 | Both | voltage-dependent anion-selective channel protein 2 | γ |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Blaz, J.; Barrera-Redondo, J.; Vázquez-Rosas-Landa, M.; Canedo-Téxon, A.; Aguirre von Wobeser, E.; Carrillo, D.; Stouthamer, R.; Eskalen, A.; Villafán, E.; Alonso-Sánchez, A.; et al. Genomic Signals of Adaptation towards Mutualism and Sociality in Two Ambrosia Beetle Complexes. Life 2019, 9, 2. https://doi.org/10.3390/life9010002
Blaz J, Barrera-Redondo J, Vázquez-Rosas-Landa M, Canedo-Téxon A, Aguirre von Wobeser E, Carrillo D, Stouthamer R, Eskalen A, Villafán E, Alonso-Sánchez A, et al. Genomic Signals of Adaptation towards Mutualism and Sociality in Two Ambrosia Beetle Complexes. Life. 2019; 9(1):2. https://doi.org/10.3390/life9010002
Chicago/Turabian StyleBlaz, Jazmín, Josué Barrera-Redondo, Mirna Vázquez-Rosas-Landa, Anahí Canedo-Téxon, Eneas Aguirre von Wobeser, Daniel Carrillo, Richard Stouthamer, Akif Eskalen, Emanuel Villafán, Alexandro Alonso-Sánchez, and et al. 2019. "Genomic Signals of Adaptation towards Mutualism and Sociality in Two Ambrosia Beetle Complexes" Life 9, no. 1: 2. https://doi.org/10.3390/life9010002
APA StyleBlaz, J., Barrera-Redondo, J., Vázquez-Rosas-Landa, M., Canedo-Téxon, A., Aguirre von Wobeser, E., Carrillo, D., Stouthamer, R., Eskalen, A., Villafán, E., Alonso-Sánchez, A., Lamelas, A., Ibarra-Juarez, L. A., Pérez-Torres, C. A., & Ibarra-Laclette, E. (2019). Genomic Signals of Adaptation towards Mutualism and Sociality in Two Ambrosia Beetle Complexes. Life, 9(1), 2. https://doi.org/10.3390/life9010002