
Halophilic Archaea: Life with Desiccation, Radiation and Oligotrophy over Geological Times


{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Specific Strategies of Haloarchaea
2.1. Low Water Activity and Desiccation
2.2. Oligotrophic Environments and Starvation
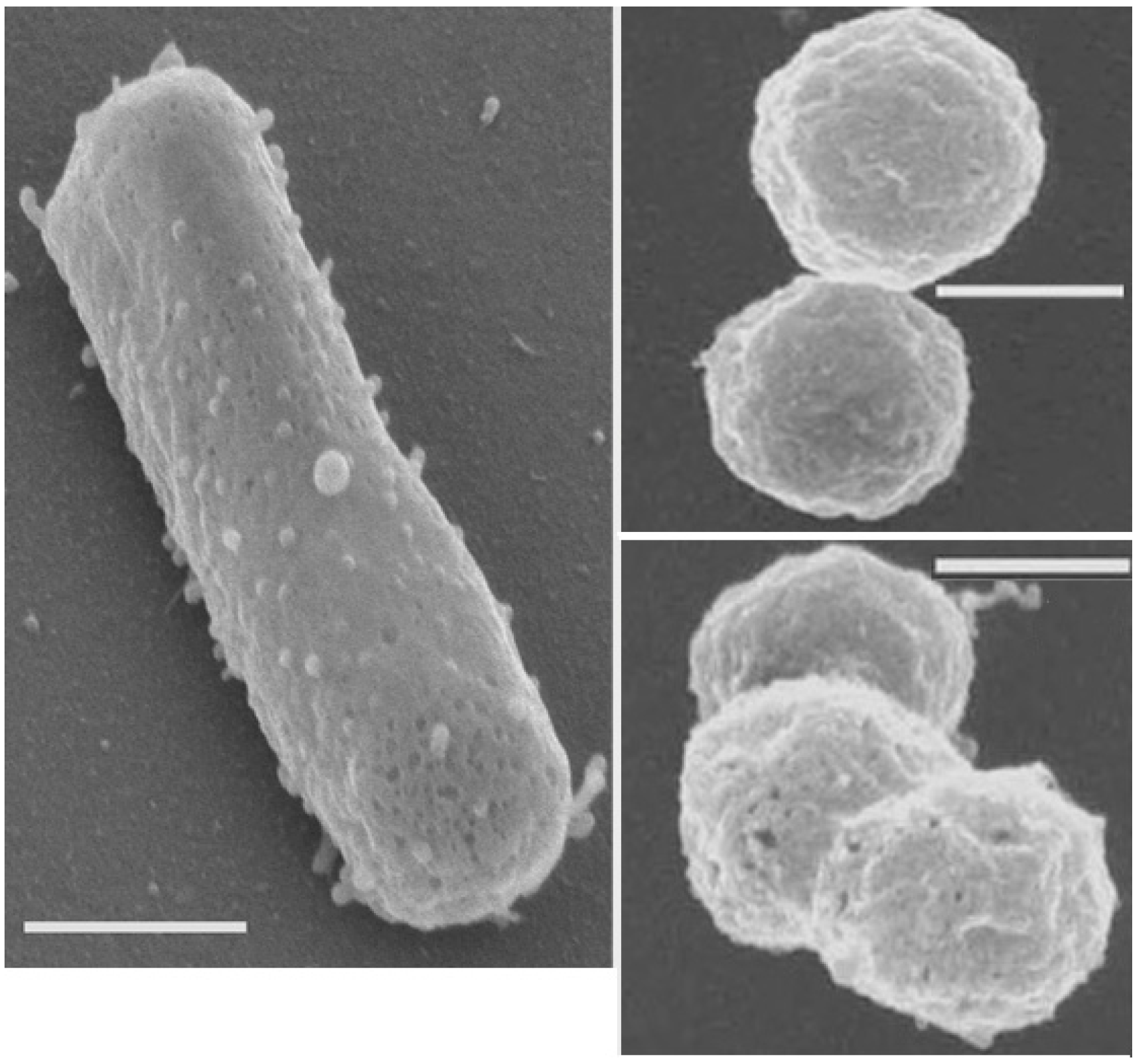
2.3. Sphere Formation in Fluid Inclusions

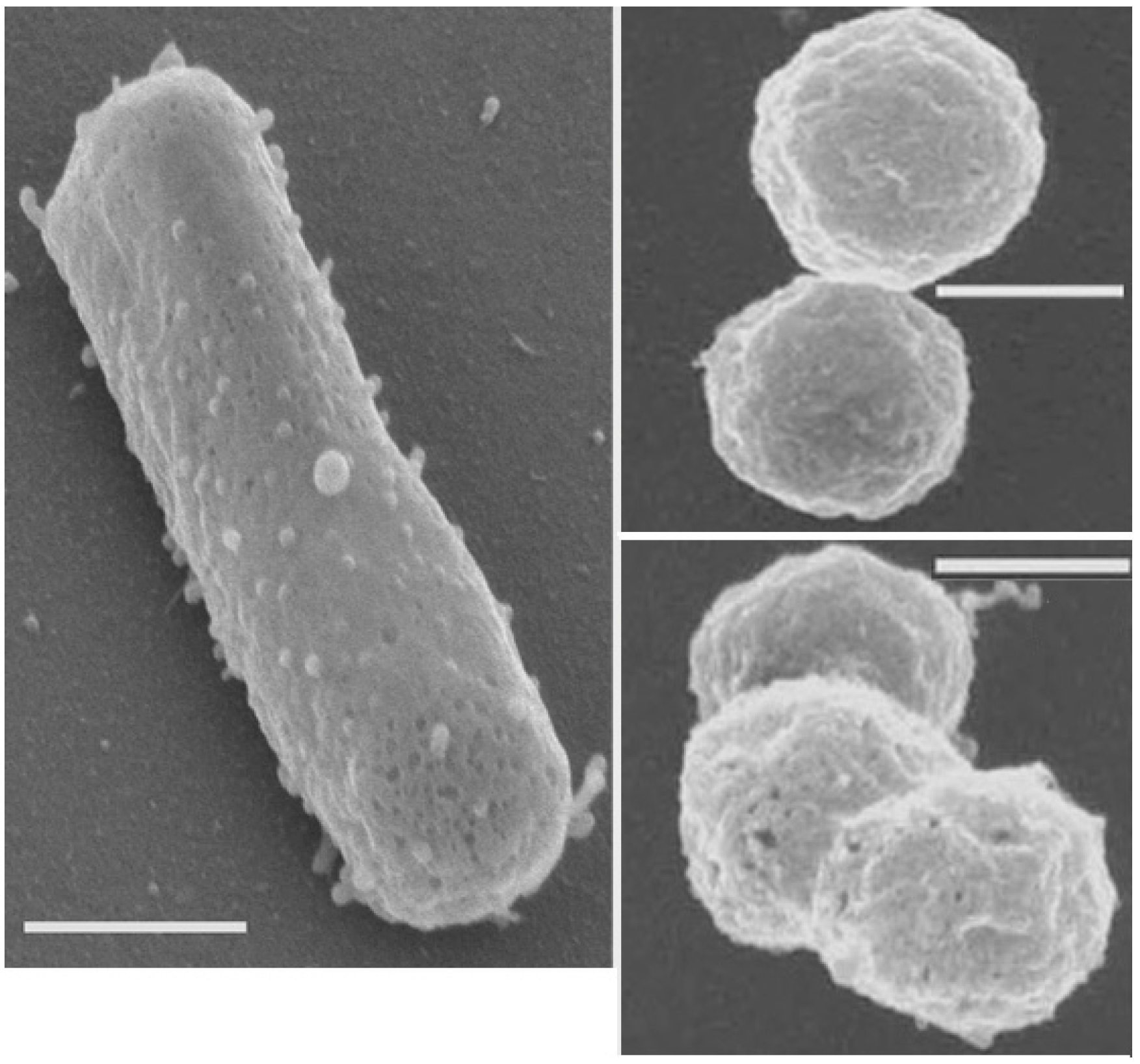
2.4. Resistance to Radiation
3. Extraterrestrial Halite
4. Conclusions
Acknowledgements
Conflicts of Interest
References
- Gramain, A.; Chong Díaz, G.C.; Demergasso, C.; Lowenstein, T.K.; McGenity, T.J. Archaeal diversity along a subterranean salt core from the Salar Grande. Environ. Microbiol. 2011, 13, 2105–2121. [Google Scholar] [CrossRef] [PubMed]
- Reiser, R.; Tasch, P. Investigation of the viability of osmophile bacteria of great geological age. Trans. Kans. Acad. Sci. 1960, 63, 31–34. [Google Scholar] [CrossRef] [PubMed]
- Dombrowski, H. Bacteria from Paleozoic salt deposits. Ann. N. Y. Acad. Sci. 1963, 108, 453–460. [Google Scholar] [CrossRef] [PubMed]
- Norton, C.F.; McGenity, T.J.; Grant, W.D. Archaeal halophiles (halobacteria) from two British salt mines. J. Gen. Microbiol. 1993, 139, 1077–1081. [Google Scholar] [CrossRef]
- Denner, E.B.M.; McGenity, T.J.; Busse, H.-J.; Wanner, G.; Grant, W.D.; Stan-Lotter, H. Halococcus salifodinae sp. nov., an archaeal isolate from an Austrian salt mine. Int. J. Syst. Bacteriol. 1994, 44, 774–780. [Google Scholar] [CrossRef]
- Stan-Lotter, H.; McGenity, T.J.; Legat, A.; Denner, E.B.M.; Glaser, K.; Stetter, K.O.; Wanner, G. Very similar strains of Halococcus salifodinae are found in geographically separated Permo-Triassic salt deposits. Microbiology 1999, 145, 3565–3574. [Google Scholar] [PubMed]
- Radax, C.; Gruber, C.; Stan-Lotter, H. Novel haloarchaeal 16S rRNA gene sequences from Alpine Permo-Triassic rock salt. Extremophiles 2001, 5, 221–228. [Google Scholar] [CrossRef] [PubMed]
- Stan-Lotter, H.; Pfaffenhuemer, M.; Legat, A.; Busse, H.-J.; Radax, C.; Gruber, C. Halococcus dombrowskii sp. nov., an archaeal isolate from a Permian alpine salt deposit. Int. J. Syst. Evol. Microbiol. 2002, 52, 1807–1814. [Google Scholar] [CrossRef] [PubMed]
- Mormile, M.R.; Biesen, M.A.; Gutierrez, M.C.; Ventosa, A.; Pavlovich, J.B.; Onstott, T.C.; Fredrickson, J.K. Isolation of Halobacterium salinarum retrieved directly from halite brine inclusions. Environ. Microbiol. 2003, 5, 1094–1102. [Google Scholar] [CrossRef] [PubMed]
- Gruber, C.; Legat, A.; Pfaffenhuemer, M.; Radax, C.; Weidler, G.; Busse, H.-J.; Stan-Lotter, H. Halobacterium noricense sp. nov., an archaeal isolate from a bore core of an alpine Permian salt deposit, classification of Halobacterium sp. NRC-1 as a strain of H. salinarum and emended description of H. salinarum. Extremophiles 2004, 8, 431–439. [Google Scholar]
- Vreeland, R.H.; Jones, J.; Monson, A.; Rosenzweig, W.D.; Lowenstein, T.K.; Timofeeff, M.; Satterfield, C.; Cho, B.C.; Park, J.S.; Wallace, A.; Grant, W.D. Isolation of live Cretaceous (121–112 million years old) halophilic Archaea from primary salt crystals. Geomicrobiol. J. 2007, 24, 275–282. [Google Scholar] [CrossRef]
- Park, J.S.; Vreeland, R.H.; Cho, B.C.; Lowenstein, T.K.; Timofeeff, M.N.; Rosenzweig, W.D. Haloarchaeal diversity in 23, 121 and 419 MYA salts. Geobiology 2009, 7, 515–523. [Google Scholar] [CrossRef] [PubMed]
- Schubert, B.A.; Lowenstein, T.K.; Timofeeff, M.N.; Parker, M.A. Halophilic Archaea cultured from ancient halite, Death Valley, California. Environ. Microbiol. 2010, 12, 440–454. [Google Scholar] [CrossRef] [PubMed]
- Saralov, A.I.; Baslerov, R.V.; Kuznetsov, B.B. Haloferax chudinovii sp. nov., a halophilic archaeon from Permian potassium salt deposits. Extremophiles 2013, 17, 499–504. [Google Scholar] [CrossRef] [PubMed]
- Jaakkola, S.T.; Zerulla, K.; Guo, Q.; Liu, Y.; Ma, H.; Yang, C.; Bamford, D.B.; Chen, X.; Soppa, J.; Oksanen, H.M. Halophilic archaea cultivated from surface sterilized middle-late Eocene rock salt are polyploid. PLoS ONE 2014, 9, e110533,. [Google Scholar] [CrossRef] [PubMed]
- Schubert, B.A.; Lowenstein, T.K.; Timofeeff, M.N. Microscopic identification of prokaryotes in modern and ancient halite, Saline Valley and Death Valley, California. Astrobiology 2009, 9, 467–482. [Google Scholar] [CrossRef] [PubMed]
- Schubert, B.A.; Timofeeff, M.N.; Lowenstein, T.K.; Polle, J.E.W. Dunaliella cells in fluid inclusions in halite: Significance for long-term survival of prokaryotes. Geomicrobiol. J. 2010, 27, 61–75,. [Google Scholar] [CrossRef]
- Lowenstein, T.K.; Schubert, B.; Timofeeff, M. Microbial communities in fluid inclusions and long-term survival in halite. GSA Today 2011, 21, 4–9. [Google Scholar] [CrossRef]
- Onyenwoke, R.U.; Brill, J.A.; Farahi, K.; Wiegel, J. Sporulation genes in members of the low G + C Gram-type-positive phylogenetic branch (Firmicutes). Arch. Microbiol. 2004, 182, 182–192. [Google Scholar] [CrossRef] [PubMed]
- Grant, W.D.; Gemmell, R.T.; McGenity, T.J. Halobacteria: The evidence for longevity. Extremophiles 1998, 2, 279–287. [Google Scholar] [CrossRef] [PubMed]
- Breuert, S.; Allers, T.; Spohn, G.; Soppa, J. Regulated polyploidy in halophilic archaea. PLoS ONE 2006, 1, e92. [Google Scholar] [CrossRef] [PubMed]
- Soppa, J. Evolutionary advantages of polyploidy in halophilic archaea. Biochem. Soc. Trans. 2013, 41, 339–343,. [Google Scholar] [CrossRef] [PubMed]
- Soppa, J. Polyploidy in archaea and bacteria: About desiccation resistance, giant cell size, long-term survival, enforcement by a eukaryotic host and additional aspects. J. Mol. Microbiol. Biotechnol. 2014, 24, 409–419,. [Google Scholar] [CrossRef] [PubMed]
- Zerulla, K.; Soppa, J. Polyploidy in haloarchaea: Advantages for growth and survival. Front. Microbiol. 2014, 5. [Google Scholar] [CrossRef] [PubMed]
- Zerulla, K.; Chimileski, S.; Näther, D.; Gophna, U.; Papke, R.T.; Soppa, J. DNA as a phosphate storage polymer and the alternative advantages of polyploidy for growth or survival. PLoS ONE 2014, 9, e94819. [Google Scholar] [CrossRef] [PubMed]
- Oren, A. Halophilic Microorganisms and Their Environments; Springer: Dordrecht, The Netherlands, 2002. [Google Scholar]
- Grant, W.D. Life at low water activity. Phil. Trans. R. Soc. B Biol. Sci. 2004, 359, 1249–1267. [Google Scholar] [CrossRef] [PubMed]
- Parkes, K.; Walsby, A.E. Ultrastructure of a gas-vacuolate square bacterium. J. Gen. Microbiol. 1981, 126, 503–506. [Google Scholar] [CrossRef]
- Bolhuis, H.; Palm, P.; Wende, A.; Falb, M.; Rampp, M.; Rodriguez-Valera, F.; Pfeiffer, F.; Oesterhelt, D. The genome of the square archaeon Haloquadratum walsbyi: Life at the limits of water activity. BMC Genomics 2006, 7. [Google Scholar] [CrossRef] [PubMed]
- Dyall-Smith, M.L.; Pfeiffer, F.; Klee, K.; Palm, P.; Gross, K.; Schuster, S.C.; Rampp, M.; Oesterhelt, D. Haloquadratum walsbyi: Limited diversity in a global pond. PLoS ONE 2011, 6, e20968. [Google Scholar] [CrossRef] [PubMed]
- Zenke, R.; von Gronau, S.; Bolhuis, H.; Gruska, M.; Pfeiffer, F.; Oesterhelt, D. Fluorescence microscopy visualization of halomucin, a secreted 927 kDa protein surrounding Haloquadratum walsbyi cells. Front. Microbiol. 2015, 6. [Google Scholar] [CrossRef] [PubMed]
- Oh, D.; Porter, K.; Russ, B.; Burns, D.; Dyall-Smith, M. Diversity of Haloquadratum and other haloarchaea in three, geographically distant, Australian saltern crystallizer ponds. Extremophiles 2010, 14, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Egli, T. How to live at very low substrate concentration. Water Res. 2010, 44, 4826–4837. [Google Scholar] [CrossRef] [PubMed]
- Morita, R.Y. Starvation-survival of heterotrophs in the marine environment. Adv. Microbiol. Ecol. 1982, 6, 171–198. [Google Scholar]
- Bolhuis, H. Walsby’s square archaeon. In Adaptation to Life at High Salt Concentrations in Archaea, Bacteria, and Eukarya; Gunde-Cimerman, N., Oren, A., Plemenitaš, A., Eds.; Springer: Dordrecht, The Netherlands, 2005; pp. 187–199. [Google Scholar]
- Fendrihan, S.; Dornmayr-Pfaffenhuemer, M.; Gerbl, F.W.; Holzinger, A.; Grösbacher, M.; Briza, P.; Erler, A.; Gruber, C.; Plätzer, K.; Stan-Lotter, H. Spherical particles of halophilic archaea correlate with exposure to low water activity—implications for microbial survival in fluid inclusions of ancient halite. Geobiology 2012, 10, 424–433. [Google Scholar] [CrossRef] [PubMed]
- Lennon, J.T. Diversity and metabolism of marine bacteria cultivated on dissolved DNA. Appl. Environ. Microbiol. 2007, 73, 2799–2805. [Google Scholar] [CrossRef] [PubMed]
- Roedder, E. The fluids in salt. Am. Mineral. 1984, 69, 413–439. [Google Scholar]
- Norton, C.F.; Grant, W.D. Survival of halobacteria within fluid inclusions in salt crystals. J. Gen. Microbiol. 1988, 134, 1365–1373. [Google Scholar] [CrossRef]
- Adamski, J.C.; Roberts, J.A.; Goldstein, R.H. Entrapment of bacteria in fluid inclusions in laboratory-grown halite. Astrobiology 2006, 6, 552–562. [Google Scholar] [CrossRef] [PubMed]
- Fendrihan, S.; Stan-Lotter, H. Survival of halobacteria in fluid inclusions as a model of possible biotic survival in martian halite. In Mars and Planetary Science and Technology; Teodorescu, H.N., Griebel, H.S., Eds.; Performantica Press: Iasi, Romania, 2004; pp. 9–18. [Google Scholar]
- Fendrihan, S.; Legat, A.; Gruber, C.; Pfaffenhuemer, M.; Weidler, G.; Gerbl, F.; Stan-Lotter, H. Extremely halophilic archaea and the issue of long term microbial survival. Rev. Environ. Sci. Biotechnol. 2006, 5, 203–218. [Google Scholar] [CrossRef] [PubMed]
- Dornmayr-Pfaffenhuemer, M.; Legat, A.; Schwimbersky, K.; Fendrihan, S.; Stan-Lotter, H. Responses of haloarchaea to simulated microgravity. Astrobiology 2011, 11, 199–205. [Google Scholar] [CrossRef] [PubMed]
- Kottemann, M.; Kish, A.; Iloanusi, C.; Bjork, S.; DiRuggiero, J. Physiological responses of the halophilic archaeon Halobacterium sp. strain NRC-1 to desiccation and gamma irradiation. Extremophiles 2005, 9, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Webb, K.M.; DiRuggiero, J. Radiation resistance in extremophiles: Fending off multiple attacks. In Polyextremophiles; Series: Cellular Origins, Life in Extreme Habitats and Astrobiology; Seckbach, J., Oren, A., Stan-Lotter, H., Eds.; Springer: Dordrecht, The Netherlands, 2013; pp. 249–267. [Google Scholar]
- DeVeaux, L.C.; Müller, J.A.; Smith, J.R.; Petrisko, J.; Wells, D.P.; DasSarma, S. Extremely radiation-resistant mutants of a halophilic archaeon with increased single-stranded DNA-binding protein (RPA) gene expression. Radiat. Res. 2007, 168, 507–514. [Google Scholar] [CrossRef] [PubMed]
- Daly, M.J. A new perspective on radiation resistance based on Deinococcus radiodurans. Nat. Rev. Microbiol. 2009, 7, 237–245. [Google Scholar] [CrossRef] [PubMed]
- Capes, M.D.; DasSarma, P.; DasSarma, S. The core and unique proteins of haloarchaea. BMC Genomics 2012, 13. [Google Scholar] [CrossRef] [PubMed]
- Kminek, G.; Bada, J.L.; Pogliano, K.; Ward, J.F. Radiation-dependent limit for the viability of bacterial spores in halite fluid inclusions and on Mars. Radiat. Res. 2003, 159, 722–729. [Google Scholar] [CrossRef]
- Westall, F.; Loizeau, D.; Foucher, F.; Bost, N.; Betrand, M.; Vago, J.; Kminek, G. Habitability on Mars from a microbial point of view. Astrobiology 2013, 13, 887–897. [Google Scholar] [CrossRef] [PubMed]
- Treiman, A.H.; Gleason, J.D.; Bogard, D.D. The SNC meteorites are from Mars. Planet. Space Sci. 2000, 48, 1213–1230. [Google Scholar] [CrossRef]
- Barber, D.J. Matrix phyllosilicates and associated minerals in C2M carbonaceous chondrites. Geochim. Cosmochim. Acta 1981, 45, 945–970. [Google Scholar] [CrossRef]
- Zolensky, M.E.; Bodnar, R.J.; Gibson, E.K.; Nyquist, L.E.; Reese, Y.; Shih, C.Y.; Wiesman, H. Asteroidal water within fluid inclusion-bearing halite in an H5 chondrite, Monahans (1998). Science 1999, 285, 1377–1379. [Google Scholar] [CrossRef] [PubMed]
- Postberg, F.; Kempf, S.; Schmidt, J.; Beinsen, A.; Abel, B.; Buck, U.; Srama, R. Sodium salts in E-ring ice grains from an ocean below the surface of Enceladus. Nature 2009, 459, 1098–1101. [Google Scholar] [CrossRef] [PubMed]
- McEwen, A.S.; Ojha, L.; Dundas, C.M.; Mattson, S.S.; Byrne, S.; Wray, J.J.; Cull, S.C.; Murchie, S.L.; Thomas, N.; Gulick, V.C. Seasonal flows on warm Martian slopes. Science 2011, 333, 740–743. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Iglesias, V.; Bonales, L.J.; Prieto-Ballesteros, O. pH and salinity evolution of Europa’s brines: Raman spectroscopy study of fractional precipitation at 1 and 300 bar. Astrobiology 2013, 13, 693–702. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stan-Lotter, H.; Fendrihan, S. Halophilic Archaea: Life with Desiccation, Radiation and Oligotrophy over Geological Times. Life 2015, 5, 1487-1496. https://doi.org/10.3390/life5031487
Stan-Lotter H, Fendrihan S. Halophilic Archaea: Life with Desiccation, Radiation and Oligotrophy over Geological Times. Life. 2015; 5(3):1487-1496. https://doi.org/10.3390/life5031487
Chicago/Turabian StyleStan-Lotter, Helga, and Sergiu Fendrihan. 2015. "Halophilic Archaea: Life with Desiccation, Radiation and Oligotrophy over Geological Times" Life 5, no. 3: 1487-1496. https://doi.org/10.3390/life5031487
APA StyleStan-Lotter, H., & Fendrihan, S. (2015). Halophilic Archaea: Life with Desiccation, Radiation and Oligotrophy over Geological Times. Life, 5(3), 1487-1496. https://doi.org/10.3390/life5031487