1. Introduction
The moderately to extremely halophilic Archaea (Haloarchaea) are adapted to salty environments with sodium chloride concentrations ranging from 1 M up to saturation at 5.3 M. They are heterotrophic, often facultatively anaerobic microorganisms living on amino acids. Bioinformatic analyses of 10 haloarchaeal and more than 1000 bacterial reference genomes imply that a common haloarchaeal ancestor, the “founder” haloarchaeon, evolved from an anaerobic methanogen by acquiring 1047 bacterial genes, thus expanding the physiological properties to an aerobic organotroph [
1]. The transferred genes mainly derive from Actinobacteria and are responsible for catabolic metabolism, membrane transporters and components required for oxygen respiration [
1,
2]. Lateral gene transfer is still a special feature of the Haloarchaea, but their aerobic lifestyle appears to be a consequence of a single mass acquisition of bacterial genes.
The first haloarchaeon characterized in the laboratory was
Halobacterium (
Hbt.)
salinarum (formerly
Hbt. halobium), isolated as a contaminant from salted fish in 1919 [
3]. All other Haloarchaea derive from salt lakes, salt flats or solar salterns.
Hbt. salinarum uses light-driven ion pumps (proton pump: bacteriorhodopsin; chloride pump: halorhodopsin; plus sensory rhodopsins) as special energy conversion and sensing systems, and similar light-driven proton pumps are also present in marine proteobacteria [
4,
5]. Bacteriorhodopsin is produced under microaerobic conditions and forms almost crystalline purple patches in the cytoplasmic membrane of
Hbt. salinarum (purple membrane, Pum)
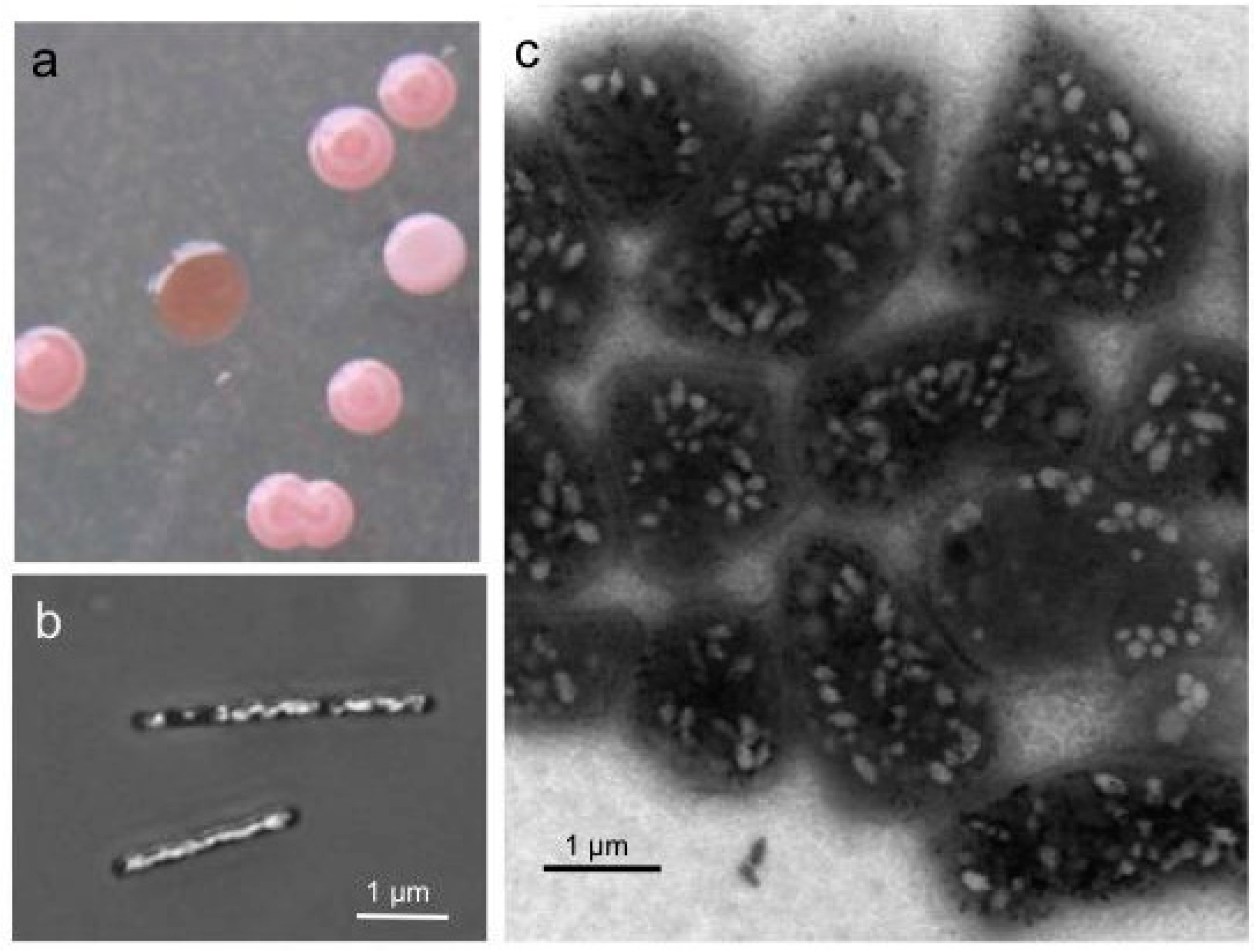
. Haloarchaea also contain C50 carotenoids (bacterioruberins, Rub) leading to red colonies on agar plates. The possession of gas vesicles (Vac) turns the colony color into pink and opaque, as opposed to the red transparent colonies of Vac
− mutants (
Figure 1a). Blooms and biofilms of Haloarchaea color brines and salt lakes red, as also seen for solar salterns at the coastline. Since Haloarchaea captured in salt crystals survive for a very long time, the use of natural sea salt was the reason why
Hbt. salinarum grew on salted fish.
Figure 1.
Colonies (a) and cells of Halobacterium (Hbt.) salinarum producing gas vesicles (b,c). (a) Colonies on solid media grown for one week at 40 °C and three weeks at room temperature. Vesicle (Vac+) cells form pink white colonies, whereas colonies of Vac− mutants are red and transparent. (b) Cells grown in liquid media observed by phase-contrast light microscopy. (c) Cells of a Vac+ colony investigated by transmission electron microscopy. The pleomorphic shape of the cells grown for three months on solid media differs from the rod-shaped cells seen in liquid culture.
Figure 1.
Colonies (a) and cells of Halobacterium (Hbt.) salinarum producing gas vesicles (b,c). (a) Colonies on solid media grown for one week at 40 °C and three weeks at room temperature. Vesicle (Vac+) cells form pink white colonies, whereas colonies of Vac− mutants are red and transparent. (b) Cells grown in liquid media observed by phase-contrast light microscopy. (c) Cells of a Vac+ colony investigated by transmission electron microscopy. The pleomorphic shape of the cells grown for three months on solid media differs from the rod-shaped cells seen in liquid culture.
Hbt. salinarum,
Haloferax (
Hfx.)
mediterranei,
Halorubrum vacuolatum and
Haloquadratum (
Hqr.)
walsbyi contain up to 70 spindle- or cylinder-shaped gas vesicles per cell, easily seen as white bodies by phase-contrast light microscopy or transmission electron microscopy (
Figure 1b,c). Fourteen
gvp genes clustered in the genomic vac region are involved in gas vesicle formation in
Hbt. salinarum and
Hfx. mediterranei [
6], whereas only 12
gvp genes (
gvpDE are lacking
) are found in the Vac
+ species
Hrr. vacuolatum and
Hqr. walsbyi [
7,
8,
9]. In contrast,
Halopiger xanaduensis,
Haladaptatus pauchihalophilus and the haloalkaliphiles,
Natrinema pellirubrum,
Natrialba magadii and
Natronobacterium gregoryi, all contain a vac region lacking
gvpC and
gvpDE [
10]. These 11
gvp genes are sufficient for gas vesicle genesis, but these species are all Vac
−. It is not known whether their
gvp gene clusters are generally not expressed or only transcribed under special conditions.
The vac regions have been characterized for the
Hbt. salinarum strains PHH1, NRC-1, SB3, GN101 and GRB [
6,
11,
12,
13]. The latter three strains are more recent natural isolates and genetically stable, whereas PHH1 (DSM 670) and NRC-1 (ATC 700922) are derived from strain collections and exhibit high mutation rates of Vac, Rub and Pum, due to the action of insertion (ISH) elements. The vac regions of PHH1 and NRC-1 were identified using the
gvpA gene probe of the cyanobacterium,
Calothrix [
11,
14]. This gene encodes the major gas vesicle structural protein, GvpA, and is highly conserved between bacterial and archaeal gas vesicle producers. Transformation experiments using the Vac
− species,
Hfx. volcanii, uncovered that
gvpA is not sufficient for gas vesicle formation and that 12 additional
gvp genes are required [
13,
15]. Furthermore, Vac
− mutants of PHH1 (and NRC-1) are usually due to the integration of an ISH-element in a 9-kbp region surrounding
gvpA [
13]. The Vac
− mutant strain, R1, whose genome sequence has been determined by the Oesterhelt Lab, incurred an ISH3 insertion upstream of p-
gvpA, preventing the transcription [
16].
Hbt. salinarum PHH1, R1 and NRC-1 all contain two different
gvp gene clusters, p-vac (
gvp1 in NRC-1) and c-vac (
gvp2 in NRC-1), whereas SB3, GN101 and GRB contain the c-vac region only [
6,
12,
17]. Expression of p-vac leads to spindle-shaped gas vesicles throughout growth, whereas the cylinder-shaped c-vac gas vesicles occur in the stationary growth phase only. The p-vac region is located on a plasmid and often incurs an ISH element. The 14
gvp genes constituting each vac region are arranged in the two oppositely oriented gene clusters,
gvpACNO and
gvpDEFGHIJKLM (
Figure 2a)
. This arrangement is typical for p-vac and c-vac, but also for the mc-vac region of
Hfx. mediterranei [
6].
Hfx. mediterranei produces cylinder-shaped gas vesicles in the stationary growth phase when grown in 25% salt-media [
18]. In contrast, twelve
gvp genes in
Hrr. vacuolatum and
Hqr. walsbyi are arranged as the
gvpACNO-FGHIJKLM gene cluster with additional DNA inserted between
gvpO and
gvpF in
Hqr. walsbyi.In this review, I will discuss the expression of the vac regions in Hbt. salinarum and Hfx. mediterranei, with emphasis on the regulation at the transcriptional and translational level. In addition, experiments to deduce the early steps in the formation of the gas vesicle wall will be presented. Furthermore, I will discuss the applications of gas vesicles in biotechnology and biomedicine.
2. Differences Between the Hbt. salinarum Strains and the Action of ISH Elements
Hbt. salinarum PHH1, R1 and NRC-1 are very closely related, but differ in the arrangement of their plasmids, due to the frequent action of various ISH elements. The nucleotide sequences determined for the chromosomes of NRC-1 and strain R1 differ by twelve point mutations only, but the plasmid sequences are completely rearranged [
16]. All three strains contain many ISH-elements, first observed with Vac, Rub or Pum mutants [
19,
20]. A 1% mutation frequency is found with Vac, but also mutants in Rub or Pum occur at frequencies of 10
−4 [
19]. Rub and Pum are chromosomally encoded, but mutants of these phenotypes always carry insertions in the respective gene regions and also incurred additional alterations in the plasmid DNA, including insertions, duplications, inversions and/or deletions [
19,
21].
Hbt. salinarum NRC-1 contains the
gvp1 gene cluster on the 190-kbp plasmid, pNRC100, whereas the almost identical p-vac region is located on plasmid pHH1 in
Hbt. salinarum PHH1 or on pHS1 in strain R1 (
Table 1) [
6,
16,
22]. The
gvp2 gene cluster resides on the mini-chromosome, pNRC200, in NRC-1, which contains a 145-kbp duplication of pNRC100, including a copy of
gvp1, whereas c-vac in R1 is located on plasmid pHS3, similar to c-vac in
Hbt. salinarum PHH1 (
Table 1). Gas vesicles due to the expression of c-vac are only found in
Hbt. salinarum PHH4, which harbors the 35-kbp pHH4, a small derivative of pHH1 lacking p-vac [
23,
24].
Table 1.
Characteristics of Hbt. salinarum strains.
Table 1.
Characteristics of Hbt. salinarum strains.
| Strains | Plasmids and vac regions | Vac phenotype | Presence of tfb and tbp | Reference |
|---|
| PHH1 Vac+ | pHH1, 150 kbp; p-vac mini-chromosome *; c-vac | spindle-shaped gas vesicles from p-vac in all stages of growth | tbp: A, B, C::ISH, D, E, F | [12,19] |
| tfb: A, B, D, F, G | [25,26] |
| PHH4 Vac+ | pHH4, 35 kbp; mini-chromosome*; c-vac | cylinder-shaped gas vesicles from c-vac in the stationary growth phase | tbp: E | [24,27] |
| tfb: A,B,D,F,G | [25,26] |
| NRC-1 Vac+ | pNRC100, 190 kbp; gvp1 pNRC200, 354 kbp; contains gvp1 and gvp2 | spindle-shaped gas vesicles from gvp1 in all stages of growth | tbp: A, B, C, D, E, F | [22] |
| tfb: A, B, C, D, E, F, G; C + E on pNRC200 | [22] |
| R1 Vac− | pHS1, 147 kbp; p-vac::ISH3
pHS3, 284 kbp; c-vac
pHS2, 195 kbp; pHS4, 40 kbp | Vac− phenotype due to an ISH3 insertion upstream of gvpA in p-vac | tbp: A, B::ISH, C, D, E, F::ISH | [16] |
| tfb: A, B, C, D, E, F, G, H | [16] |
These plasmid alterations also affect the number of genes encoding the general transcription factors, TFB and TBP (
Table 1). TFB binds to the BRE sequence adjacent to the TATA-box in the archaeal promoter and recruits, together with the TATA-box binding protein TBP, the RNA polymerase for transcription initiation. Many of the
tfb and
tbp genes are plasmid-encoded in
Hbt. salinarum [
16,
22]. Six different
tbp are present, but
tbpB,
tbpF or
tbpC in R1 and PHH1 are inactivated by insertion elements [
25] (
Table 1). Strain PHH4 even lacks all of the
tbp genes, except for the chromosomal
tbpE, which is the only essential
tbp under standard conditions. Differences are also seen with respect to TFB (
Table 1) [
26]. All of these variations in
tfb and
tbp genes might explain the differences in the amount of gas vesicles in
Hbt. salinarum PHH1 and NRC-1 when both are grown under non-standard conditions. A systems analysis of the expression of the seven TFBs under different conditions in
Hbt. salinarum NRC-1 reveals that both the promoter and protein-coding sequences of
tfb are important in encoding environment-dependent regulatory programs for rapid adaptation to environmental niches [
28].
The
Hbt. salinarum strains, GN101, GRB and SB3, are derived from salt flats in Mexico (GN101), France (GRB) or the USA (SB3) [
29]. Their 16S rRNA genes exhibit >99% sequence identity with the 16S rRNA gene of
Hbt. salinarum PHH1 and NRC-1. All three natural isolates produce cylinder-shaped gas vesicles in late exponential growth due to the expression of c-vac, similar to
Hbt. salinarum PHH4. The mutation rate with respect to the Vac phenotype is low, since all three contain much less ISH elements compared to PHH1 and NRC-1. The plasmids found here differ from each other, except for the small, multi-copy plasmids, pGN1, pHSB1, and pGRB1, which are homologous (
Table 2). All three strains lack plasmids related to the pHH1/pNRC100/pHS1 family and, thus, also p-vac. The lack of the pHH1-type plasmids implies that p-vac might be derived from a lateral gene transfer event.
Table 2.
Plasmids of Hbt. salinarum GRB, SB3 and GN101.
Table 2.
Plasmids of Hbt. salinarum GRB, SB3 and GN101.
| Strains | Plasmids and vac regions | Comments | Reference |
|---|
| GN101 Vac+ | Plasmids: 39, 43 and 1.7 kbp (pGN1) mini-chromosome; c-vac | Plasmids not related to pHH1/pNRC100/pHS1;1.7-kbp plasmids are homologous | [12,29] |
| SB3 Vac± | Plasmids: 34, 52, and 1.7 kbp (pHSB1) mini-chromosome; c-vac | Plasmids not related to pHH1/pNRC100/pHS1;1.7-kbp plasmids are homologous | [12,29] |
| GRB Vac+ | Plasmids: 35, 65, and 1.7 kbp (pGRB1) mini-chromosome; c-vac | Plasmids not related to pHH1/pNRC100/pHS1;1.7-kbp plasmids are homologous | [12,29] |
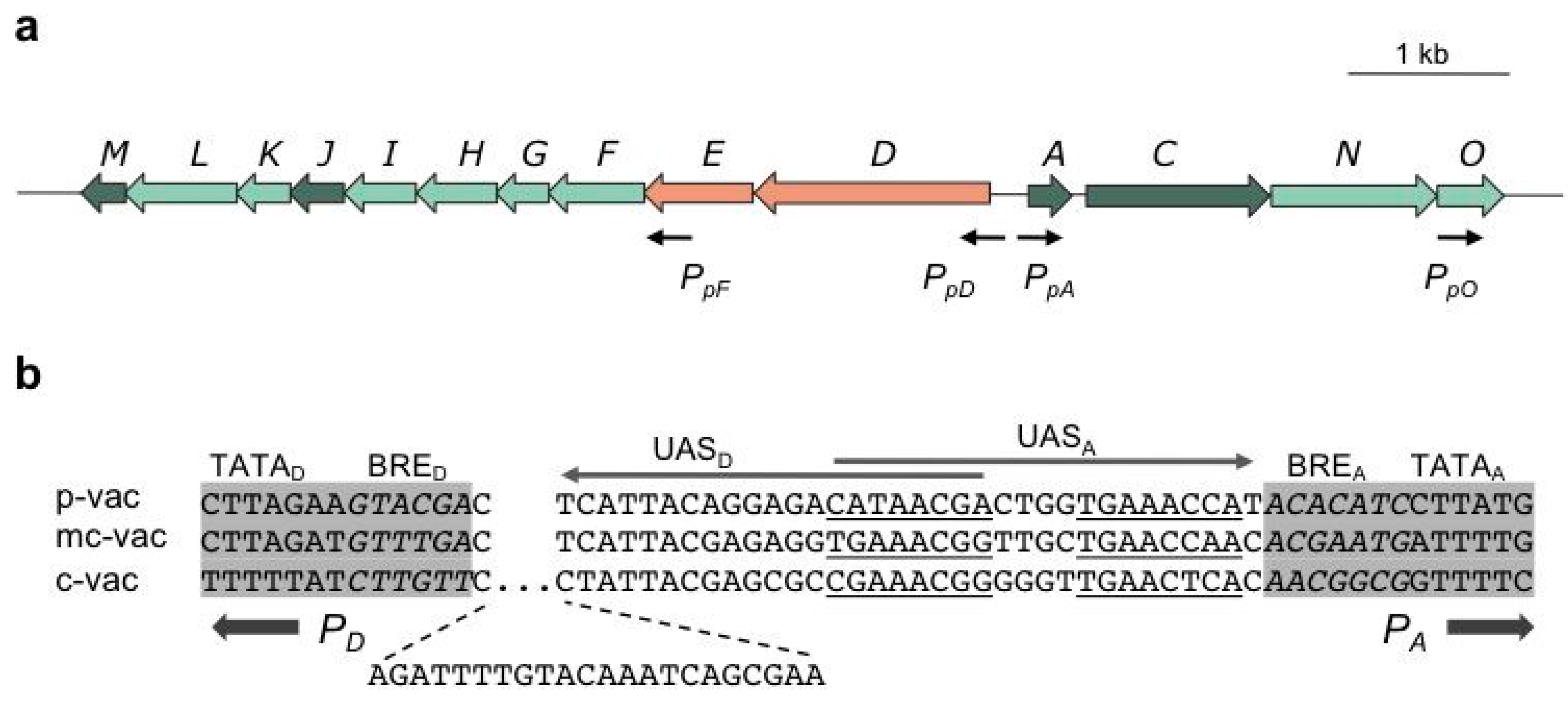
3. Regulation at the Level of Transcription Involves GvpE and GvpD
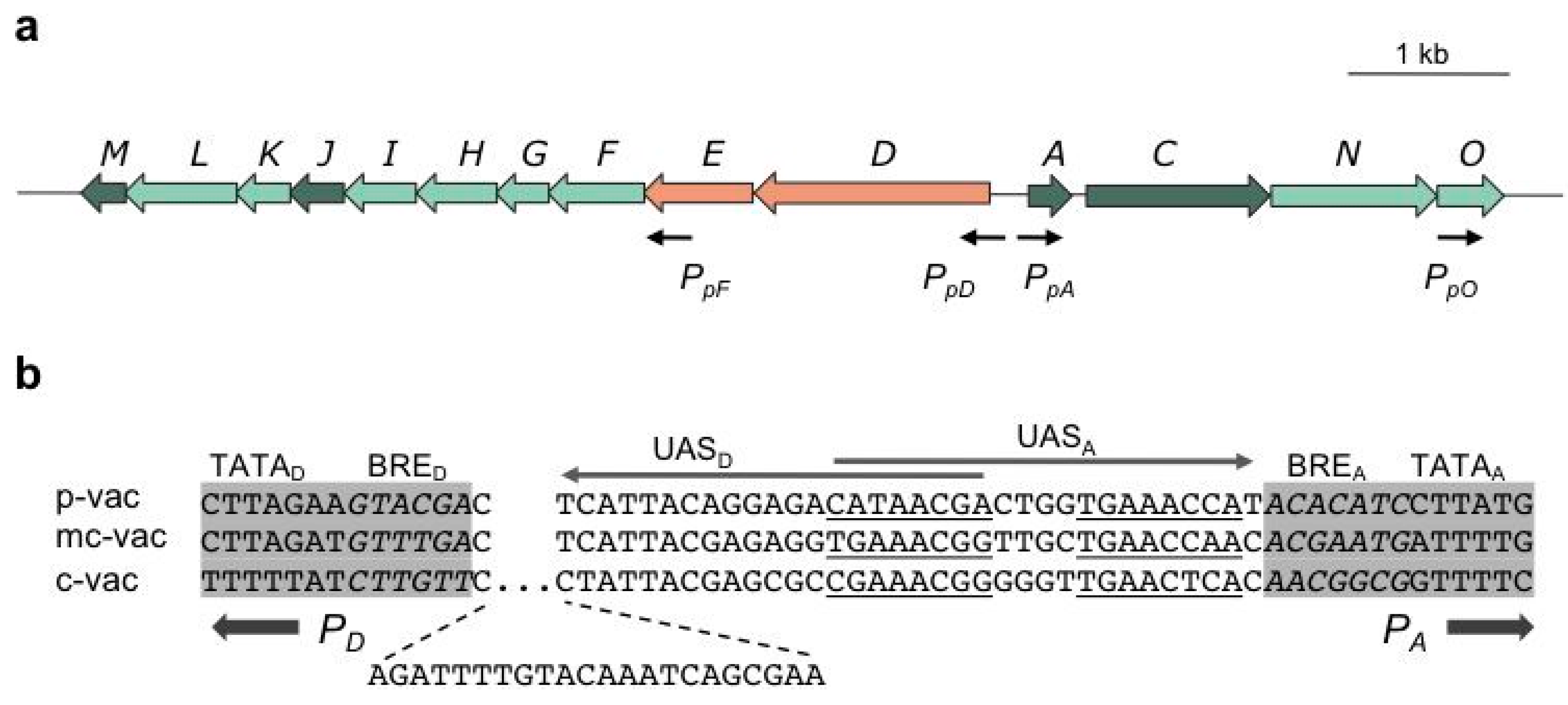
The p-vac region of
Hbt. salinarum PHH1 contains the 14
gvp genes in two oppositely-arranged gene clusters, p-
gvpACNO and p-
gvpDEFGHIJKLM (
Figure 2a). The intervening sequence between the mRNA start sites of p-
gvpA and p-
gvpD is 108 base pairs (bp) in size, with the TATA-BRE elements of the
PpA and
PpD promoters separated by 35 bp (
Figure 2b).
PpA, driving the expression of p-
gvpACNO, is the strongest promoter of p-vac, due to a 70- to 100-fold induction by the endogenous transcriptional activator, GvpE [
30]. The co-transcript contains a 20-nt 5'-untranslated region (5'-UTR), and the major 320-nt p-
gvpA transcript supports the massive GvpA production required to form the gas vesicle wall, which mainly consists of GvpA (>95%). A stem-loop structure occurs between p-
gvpA and p-
gvpC, and read-through leads to the minor amounts of the p-
gvpAC, p-
gvpACN and p-
gvpACNO co-transcripts. The stem-loop was thought to be a transcription terminator, but a deletion of this structure results in similarly low amounts of read-through transcripts, as with the wild-type arrangement, implying that the stem-loop stabilizes the p-
gvpA mRNA, rather than being a termination signal. In case of mc-vac, the half-life of the mc-
gvpA transcript is almost 90 min, whereas the half-life of the 3-kb mc-
gvpACNO transcript is 10 min, which is significantly shorter [
31]. The p-
gvpC gene downstream of p-
gvpA encodes the second structural protein, GvpC, which attaches to the outer surface of the gas vesicles to stabilize the gas vesicle wall. The functions of GvpN and GvpO are not yet known.
The oppositely-oriented
PpD promoter has a weak basal activity that is 20- to 40-fold activated by GvpE. The resulting p-
gvpDE transcript encodes the two regulatory proteins, GvpD and GvpE. The GvpE protein is one of the few transcriptional activators characterized in Archaea. It dimerizes in solution and resembles a leucine-zipper protein [
32]. The GvpE-responsive elements, UAS
A and UAS
D, occur adjacent to the respective BRE and are 20-nucleotides (nt) in size (
Figure 2b). Each element consists of two 8-nt sequences separated by 4 nt that are not important for the activation [
33,
34]. Since
PpA and
PpD initiate transcription in opposite directions, the distal portions of UAS
D and UAS
A overlap by 7 nt in the center (
Figure 2b).
In vitro studies by protein affinity chromatography yield that GvpE interacts with any TFB and TBP tested [
26,
35]. Binding constants between GvpE and the various general transcription factors have not been determined. Putative differences in the GvpE-TFB and/or GvpE-TBP affinities might cause alterations in the environment-dependent regulation of
gvp gene expression. GvpE could stimulate their binding at
PpA and
PpD to enhance the transcription initiation.
Figure 2.
Arrangement of gvp genes in p-vac of Hbt. salinarum PHH1 (a) and a comparison of the intergenic regions separating PD and PA (b). (a) The arrows depicting genes are colored as follows: dark green, encoding structural proteins of the A-J-M family and GvpC; light green, encoding accessory Gvp; red, encoding regulator proteins. Black arrows mark the start sites of transcription. (b) Comparison of the intergenic regions separating PA and PD in p-, mc- and c-vac. A 22-nt insertion occurs in c-vac adjacent to BRED. The 20-nt sequence required for GvpE-mediated activation is underlined (8-nt elements separated by 4 nt of unimportant sequences) in the case of UASA and marked by an arrow. Similar activation elements are found with UASD, pointing in the opposite direction. The TATA-box and BRE sequences (italics) are shaded in grey.
Figure 2.
Arrangement of gvp genes in p-vac of Hbt. salinarum PHH1 (a) and a comparison of the intergenic regions separating PD and PA (b). (a) The arrows depicting genes are colored as follows: dark green, encoding structural proteins of the A-J-M family and GvpC; light green, encoding accessory Gvp; red, encoding regulator proteins. Black arrows mark the start sites of transcription. (b) Comparison of the intergenic regions separating PA and PD in p-, mc- and c-vac. A 22-nt insertion occurs in c-vac adjacent to BRED. The 20-nt sequence required for GvpE-mediated activation is underlined (8-nt elements separated by 4 nt of unimportant sequences) in the case of UASA and marked by an arrow. Similar activation elements are found with UASD, pointing in the opposite direction. The TATA-box and BRE sequences (italics) are shaded in grey.
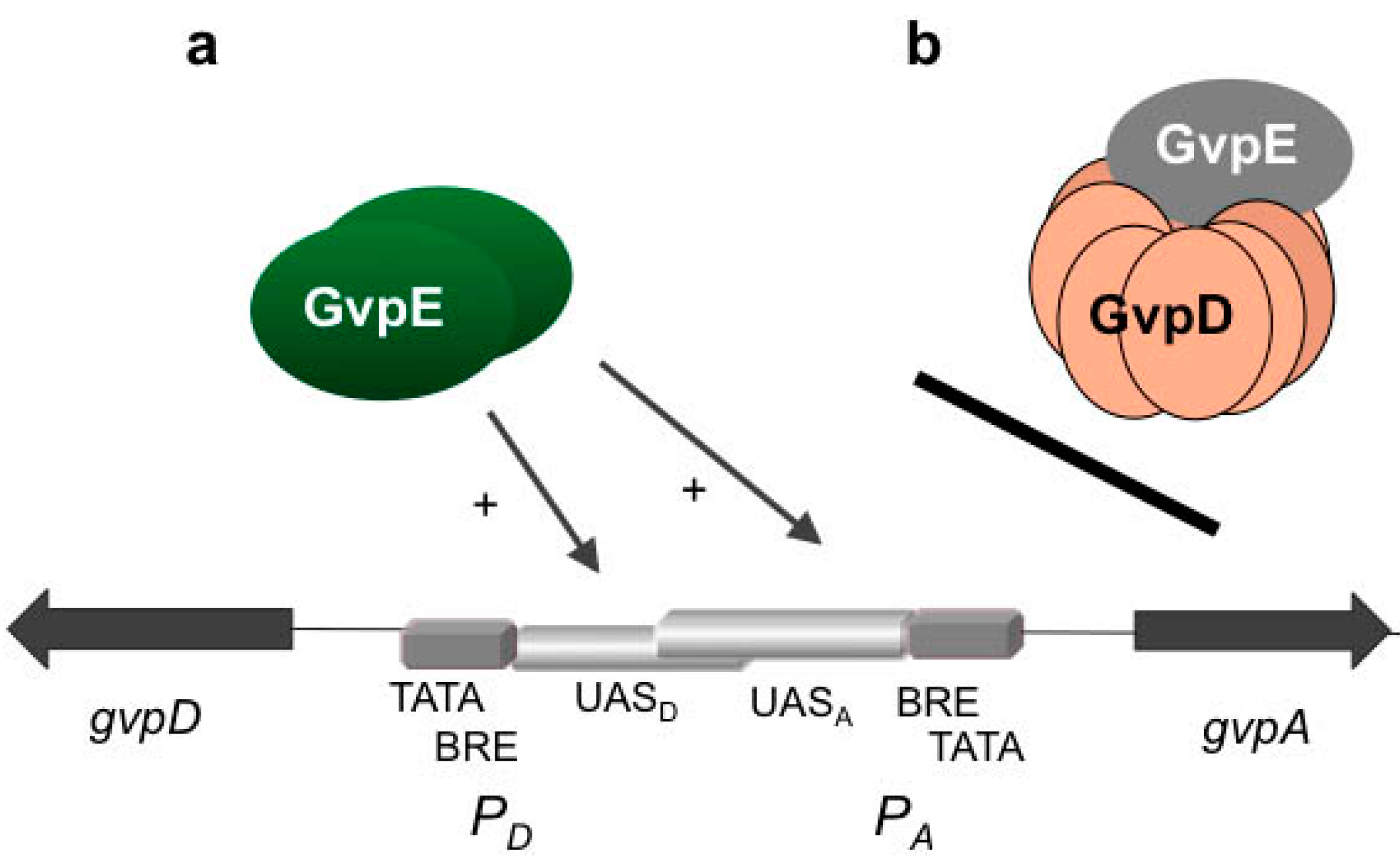
The presence of GvpE leads to a fast and strong activation of
PpA and
PpD under standard growth conditions, but as soon as GvpD appears, the activation is reduced (
Figure 3). GvpD and GvpE are able to interact, and the presence of GvpD leads to an almost undetectable amount of GvpE, as determined by western analysis [
36]. The reduction in the amount of GvpE was quantified using the green fluorescent protein smGFP fused to the N-terminus of GvpE [
37]. The amount of GFP-GvpE is more than 60% reduced in the presence of GvpD. The fact that a reduction in the amount of GvpE is not observed in the presence of the repression defective GvpD
Mut6 mutant underlines that a functional GvpD is required for this process [
37].
The repressing function of GvpD was initially observed in ∆D transformants carrying the mc-vac region of
Hfx. mediterranei with a 918-bp deletion in the
gvpD reading frame (∆D construct) [
38].
Hfx. volcanii ∆D transformants are filled with numerous gas vesicles, turning the flat and disc-shaped cells into spheres, and the presence of GvpD in ∆D + D transformants reduces the amount of gas vesicles to the wild-type level. GvpD oligomerizes in solution and exhibits similarities to the large family of AAA+ ATPase proteins by possessing a Walker I and II motif to bind ATP and an arginine-rich region [
39]. Both motifs are required for the GvpD-induced breakdown of GvpE. The repression defective GvpD
Mut6 mentioned above carries mutations in the Walker motif, underlining that nucleotide binding is important for the GvpD function [
40]. A second arginine-rich region close to the C-terminus of GvpD is also involved in repression, since an alteration of this 494RRR496 sequence to three alanine residues yields the super-repressor, GvpD
3-AAA [
40]. In presence of GvpD
3-AAA, the fluorescence of GFP-GvpE is reduced to 20% of the original fluorescence in
Hfx. volcanii transformants [
37].
Figure 3.
Activation of PA-PD by GvpE (a) and repression by GvpD (b). Schematic representation of the region between gvpA and gvpD and the two oppositely-oriented promoters PA and PD. TATA-box and BRE are shown in grey, and the two UAS elements are partly overlapping in the center in light grey. The reading frames gvpD and gvpA are represented by dark arrows. (a) Activation of transcription by GvpE should involve binding of GvpE, presumably as a dimer, at the respective UAS element and recruitment of TFB, TBP and of the RNA polymerase. (b) In the presence of GvpD, the interaction of GvpE-GvpD leads to a strong reduction in the amount of GvpE and the repression of gas vesicle formation.
Figure 3.
Activation of PA-PD by GvpE (a) and repression by GvpD (b). Schematic representation of the region between gvpA and gvpD and the two oppositely-oriented promoters PA and PD. TATA-box and BRE are shown in grey, and the two UAS elements are partly overlapping in the center in light grey. The reading frames gvpD and gvpA are represented by dark arrows. (a) Activation of transcription by GvpE should involve binding of GvpE, presumably as a dimer, at the respective UAS element and recruitment of TFB, TBP and of the RNA polymerase. (b) In the presence of GvpD, the interaction of GvpE-GvpD leads to a strong reduction in the amount of GvpE and the repression of gas vesicle formation.
The additional promoters,
PpF and
PpO, of the p-vac region are independent of GvpE and drive the synthesis of accessory Gvp proteins [
33].
PpO yields the essential GvpO throughout growth, but the function of this protein is not yet known.
PpF is active during the exponential and early stationary growth phase and leads to minor amounts of the p-
gvpFGHIJKLM transcript [
41]. The transcript start occurs 177-nt within the
gvpE reading frame. Since the AUG start codon of
gvpF occurs 8-nt within
gvpE, the p-
gvpF-M transcript contains a 169-nt 5'-UTR. The amounts of the accessory proteins, GvpF through GvpM, are much lower compared to GvpA and GvpC, and except for GvpI and GvpH, all of them are required for the formation of the gas vesicle wall [
42].
A very similar GvpE-mediated activation of
PA and
PD occurs in the mc-vac region, whereas in the case of c-vac,
PcA is the only promoter induced by GvpE.
PcD incurred a 22-bp insertion adjacent to BRE, enlarging the distance to UAS
D and, thus, abolishing GvpE-activation (
Figure 2b) [
6,
43]. The close distance of UAS-BRE is essential for the activation [
34]. The basal activity of
PcD is very weak and yields minor amounts of c-
gvpDEFGHIJKLM transcripts in late exponential growth. Additional promoters are neither found in mc-vac nor in c-vac.
All of these analyses demonstrate the importance of PA and PD for the formation of gas vesicles. The transcriptional activation by GvpE leads to the large amounts of the major gas vesicle structural protein, GvpA. The amount of the activator, GvpE, is reduced in the presence of GvpD and, thus, also, the production of gas vesicles. To investigate the function of the GvpE/GvpD regulatory system in more detail, crystal structures of both proteins are required. Furthermore, additional mutants inactive in regulation will be useful to pinpoint other regions important for the regulatory functions.
4. Environmental Factors Influencing Transcription
Oxygen availability, temperature, salt concentration and carbon sources all influence the formation of gas vesicles in Haloarchaea [
18,
26,
44,
45,
46]. Growth under anoxic conditions is supported by nitrate respiration (
Hfx. mediterranei) or arginine fermentation in the case of
Hbt. salinarum. The amounts of the
gvpACNO and
gvpD-M transcripts are 10-fold reduced in both species under anoxic conditions, and only a few small gas vesicles are formed [
45] (
Table 3).
Hfx. volcanii mc-vac transformants grown by nitrate respiration also show a 10-fold reduced expression. It is interesting to note that aerobic ∆D transformants (
i.e., mc-vac with a 918-bp deletion in mc-
gvpD) are filled with numerous gas vesicles, but lack these structures completely when grown under anoxic conditions. Thus, the absence of gas vesicles under anoxic conditions is independent of the repressing protein, GvpD. The extremely large amount of mc-
gvpA transcripts in ΔD transformants is 10-fold reduced under these conditions, but still large compared to the wild-type. Furthermore, GvpA is present in relatively large amounts in ∆D transformants, implying that the gas vesicle formation is inhibited at the level of assembly [
45]. The lower amount of ATP produced under anoxic conditions and/or the smaller amounts of accessory proteins might restrict the formation of gas vesicles under these conditions. However,
Hbt. salinarum PHH1 and NRC-1 grown by anaerobic respiration with dimethylsulfoxide (DMSO) or trimethylamine-
N-oxide (TMAO) as the terminal electron acceptor produce many gas vesicles, demonstrating that a lack of oxygen does not always lead to a reduced number of gas vesicles [
45,
46].
The effect of temperature on gas vesicle formation was studied with
Hbt. salinarum PHH1 and NRC-1 [
26,
47]. Heat shock conditions (1 h at 49 °C followed by 3 h at 56 °C and subsequent growth at 37 °C) lead to an eight-fold reduced amount of p-
gvpA transcripts in PHH1, and the gas vesicle formation stops completely (
Table 3). The p-
gvpA mRNA is rapidly lost during heat shock treatment [
26]. A reduction in the
gvpE mRNA encoding the transcriptional activator, GvpE, might also contribute to the interruption of gas vesicle formation, as determined for NRC-1 [
47]. In contrast, PHH4 is able to cope very well with heat shock conditions. A reduction in growth is not observed; nevertheless, the amount of c-
gvpA drops significantly, and gas vesicle formation stops [
26]. It is possible that the secondary structure of the
gvpA mRNA contributing to the stability of the transcript is destroyed under heat shock conditions. PHH1 and NRC-1 grown at the relatively cold temperature of 15 °C contain very large amounts of gas vesicles. A two-fold increase (PHH1) in the expression of p-
gvpACNO was determined by quantitative RT-PCR [
26], and a 1.8- to 8-fold increase of
gvpACNO and a three-fold increase in
gvpDE expression were determined for NRC-1 by microarray analyses [
47]. The higher transcription of the
gvp gene clusters under cold conditions is exceptional, since most housekeeping genes of
Hbt. salinarum are downregulated at 15 °C. GvpE might activate transcription continuously, since the amount is not reduced. The interaction of GvpD and GvpE (resulting in the degradation of GvpE) might be disturbed or the proteases involved in the breakdown of GvpE are less functional at 15 °C [
26].
Table 3.
Environmental factors influencing gas vesicle formation.
Table 3.
Environmental factors influencing gas vesicle formation.
| Environmental Factors | Strains and Conditions | Effect on Vac | Effect on gvp Transcription |
|---|
| Anaerobiosis | Hbt. salinarum p-vac | | |
| +arginine | Vac− | 10-fold reduced compared to oxic growth |
| +TMAO | Vac+ | not altered compared to oxic growth |
| +DMSO | Vac+ | not altered compared to oxic growth |
| Temperature | Hbt. salinarum p-vac | | |
| 15 °C | Vac++ | 2-fold increase compared to 37 °C |
| heat shock | Vac− | 8-fold reduced compared to 37 °C |
| Salt concentration | Hfx. mediterranei mc-vac | | |
| 15% salt | Vac− | - |
| 25% salt | Vac+ | 7-fold increase compared to 15% salt |
| Glucose | Hfx. mediterranei mc-vac | | |
| 50 mM | Vac± | - |
| 100 mM | Vac− | 10-fold reduced compared to standard |
| 200 mM | Vac− | - |
The sodium chloride concentration of the medium influences the gas vesicle formation of the moderately halophilic
Hfx. mediterranei. Gas vesicles are only formed when this species is grown in 17%–30% (w/v) salt media, whereas cells grown in 15% salt media are Vac
− (
Table 3). The amount of the mc-
gvpA transcripts is seven-fold enhanced in cells grown in 25%-salt compared to the 15%-salt culture [
18]. Glucose also influences the
PmcA promoter activity, since the addition of 50, 100 or 200 mM glucose results in a 10-fold reduction of the basal activity of
PmcA, whereas the activity of the promoter of the housekeeping ferredoxin gene,
Pfdx, is almost unaffected [
44]. A complete inhibition of gas vesicle formation is observed at glucose concentrations above 100 mM. Maltose and sucrose impose a similar effect, whereas xylose, arabinose, lactose, pyruvate and 2-deoxy-glucose have no influence [
44]. Also in this case, ∆D transformants lack gas vesicles in the presence of glucose, demonstrating that the repressing protein, GvpD, is not involved. The function of GvpD as a glucose sensor is thus excluded.
In summary, the GvpE/GvpD regulatory system is often not involved in the transfer of environmental signals to the respective vac region. It appears that both regulatory proteins are mainly required to enhance the gas vesicle formation under standard conditions in
Hbt. salinarum and
Hfx. mediterranei. Since most gas vesicle gene clusters of other Haloarchaea lack
gvpDE, the impact of GvpD and GvpE on
gvp gene expression is confined to a few species [
10]
. 5. Relevance of 5'-UTR and Shine-Dalgarno Sequence for Translation
The transcription of p-vac starts with the p-
gvpFGHIJKLM in the early exponential growth phase, followed by p-
gvpDE, p-
gvpACNO and
p-gvpO. Except for p-
gvpO, all transcripts contain a 5'-untranslated region (5'-UTR), with sizes of 20-nt (p-
gvpACNO), 72-nt (p-
gvpDE) and 169-nt (p-
gvpFGHIJKLM). With the exception of p-
gvpA and p-
gvpO, all reading frames are preceded by a putative Shine-Dalgarno (SD) sequence (GGAGGUCA) that appears at a distance of 5 to 13 nt of the AUG start codon [
48]. The SD element is complementary to a sequence found near the 3' end of the small ribosomal subunit rRNA of
Hbt. salinarum. In bacteria, SD sequences facilitate the transcript recognition at the 30S ribosomal subunit, whereas in Archaea, their importance is under debate.
An almost perfect SD sequence is found 7 nt upstream of the AUG of the p-
gvpH reading frame. A 4-nt scanning mutagenesis of this region uncovered that mutations in the SD sequence lead to a 5%–50% reduction in translation, as quantified using
bgaH encoding a haloarchaeal β-galactosidase as the reporter [
48,
49]. However, even a complete alteration of this element leads to a residual BgaH activity of 20% compared to the wild-type. Altering the spacing of the SD element of p-
gvpH with respect to the AUG start codon yields a good translation with distances of 4 to 10 nt, but a spacing of only 1 nt results in the loss of translation. These observations suggest that the SD sequence supports translation, but does not strictly control the expression of a reading frame [
48]. These results differ from the analysis of SD sequences in the hyperthermophilic archaeon,
Sulfolobus solfataricus, where single nucleotide alterations in the SD sequence completely prevent translation
in vitro [
50]. Another investigation of a putative SD sequence in the 5'-UTR of the single
sod gene of
Hfx. volcanii shows that partial mutations or even a complete removal of the SD sequence has no effect on the translation efficiency, implying that this sequence has no function [
51]. Taking the massive acquisition of bacterial genes in the haloarchaeal founder organism into account [
1,
2], the SD sequences might be remnants of bacterial origin with little or no influence on the expression of single genes. However, it is remarkable that genes organized in operons, such as in the vac regions, usually contain SD sequences that influence translation, as shown for p-
gvpH and p-
gvpG [
48].
A much larger influence on translation is even achieved when the 5'-UTR is lacking. Bioinformatic analyses predict that 30% of the haloarchaeal genes are transcribed as leaderless mRNA, including the ferredoxin transcript starting 1 nt upstream of the AUG start codon [
52]. Ferredoxin constitutes 2% of the total proteins in
Hbt. salinarum, and the
fdx gene is strongly transcribed and efficiently translated. The haloarchaeal expression vector pJAS35 is based on the strong expression of the leaderless transcripts under
Pfdx promoter control [
53]. In the case of the p-vac region, a deletion of the entire 5'-UTR of p-
gvpH (
i.e., creating a leaderless transcript) results in a 15-fold enhanced translation [
48]. Moreover, all leaderless transcripts constructed for p-vac yield a stronger expression compared to their leader-containing counterparts. These results imply that 5'-UTRs regulate gene expression by reducing the translation efficiency. The 5'- or 3'-UTRs of the haloarchaeal transcripts often form secondary structures that stabilize the mRNA (as seen with the p-
gvpA transcript) or these regions offer binding sites of proteins or small regulatory RNAs, influencing translation initiation. The exploration of the regulatory potential of 5'- and 3'-UTRs is still ongoing and will certainly yield further information.
6. Effect of Deletions and Overexpression of Single gvp Genes on Gas Vesicle Formation
The importance of the various Gvp proteins for gas vesicle formation was deduced from
Hfx. volcanii transformants carrying p-vac constructs that incurred a deletion of a single
gvp gene (∆X constructs) and also from transformants containing p-vac plus an additional
gvp gene overexpressed in pJAS35. The deletion analyses determined the eight genes,
gvpA,
F,
G,
H,
J,
K,
L,
M and
O, as essential for the synthesis of gas-filled compartments [
42]. Deletion of p-
gvpDE results in minor amounts of gas vesicles, whereas a deletion of
gvpI,
gvpH,
gvpN or
gvpC leads to extremely long (∆I), fragile (∆H, ∆C), small (∆N) or large and egg-shaped (∆C) gas vesicles. Complementation of such ∆X transformants by the missing
gvp gene again results in gas vesicles similar to
Hbt. salinarum PHH1 [
42]. An additional study has been performed with the
gvp1 gene cluster of
Hbt. salinarum NRC-1 using insertions of a kanamycin-resistance cassette (κ-element) in various
gvp genes generated in
Escherichia coli [
54]. The effect of this linker-scanning mutagenesis on gas vesicle formation was studied in a pNRC100-deficient
Hbt. salinarum NRC-1 variant still containing a partially expressed
gvp2 gene cluster. The results of the two studies (deletions
versus insertions) differ in six out of 14 cases, and possible reasons have been discussed earlier [
55]. They include also polar effects of the κ-insertion on the expression of
gvp genes located downstream of the integration site.
An overexpression of
gvpF,
G,
H,
I,
J,
K,
L or
M in the presence of p-vac does not affect gas vesicle formation, but the presence of large amounts of GvpG, GvpH or GvpM results in Vac
− transformants [
56]. Since p-vac + GHIJKLM
ex transformants (
gvpG-M inserted in the expression vector pJAS35) produce gas vesicles similar to the wild-type, a balanced amount of the accessory proteins is obviously required for gas vesicle assembly. It is possible that GvpG, GvpH or GvpM interact with other essential Gvp proteins produced in minor amounts by p-vac, thus diminishing the amount(s) of the essential partner protein(s) required for gas vesicle assembly. GvpG and GvpH are soluble, but the hydrophobic GvpM forms aggregate when produced in large amounts, easily seen with GvpM fusions to the green fluorescent smGFP. Such GvpM-GFP (M
GFP) transformants containing one to two large fluorescent foci per cell. In contrast, cells containing H
GFP or L
GFP are fully fluorescent [
57]. Together with GvpJ and GvpA, GvpM belongs to the A-J-M family of hydrophobic Gvp proteins. P-vac + MX
ex transformants (X = G, H, J or L) demonstrate that GvpH, GvpJ or GvpL are able to neutralize the inhibitor effect of GvpM, whereas GvpG is unable to do so. Protein affinity chromatography confirms that GvpM interacts with GvpH, GvpJ or GvpL, but not with GvpG, supporting the hypothesis that M-H, M-J and M-L complexes are formed. L
GFP transformants are fully fluorescent, but stable M-L
GFP aggregates occur in the presence of GvpM. In contrast, M-H aggregates are neither observed in H
GFP-M nor in H-M
GFP transformants carrying GFP fused to the C-terminus of GvpH (H
GFP) or GvpM (M
GFP) [
57]. The product could be too small to be visualized as fluorescence foci in the cells. GvpH might prevent the unspecific aggregation of GvpM by keeping this protein “soluble” and, thus, useful for incorporation in the gas vesicle wall. Similar analyses with other accessory Gvp proteins will unravel additional complexes that might occur during gas vesicle assembly.
7. Gas Vesicle Wall and Mutations Affecting the Shape
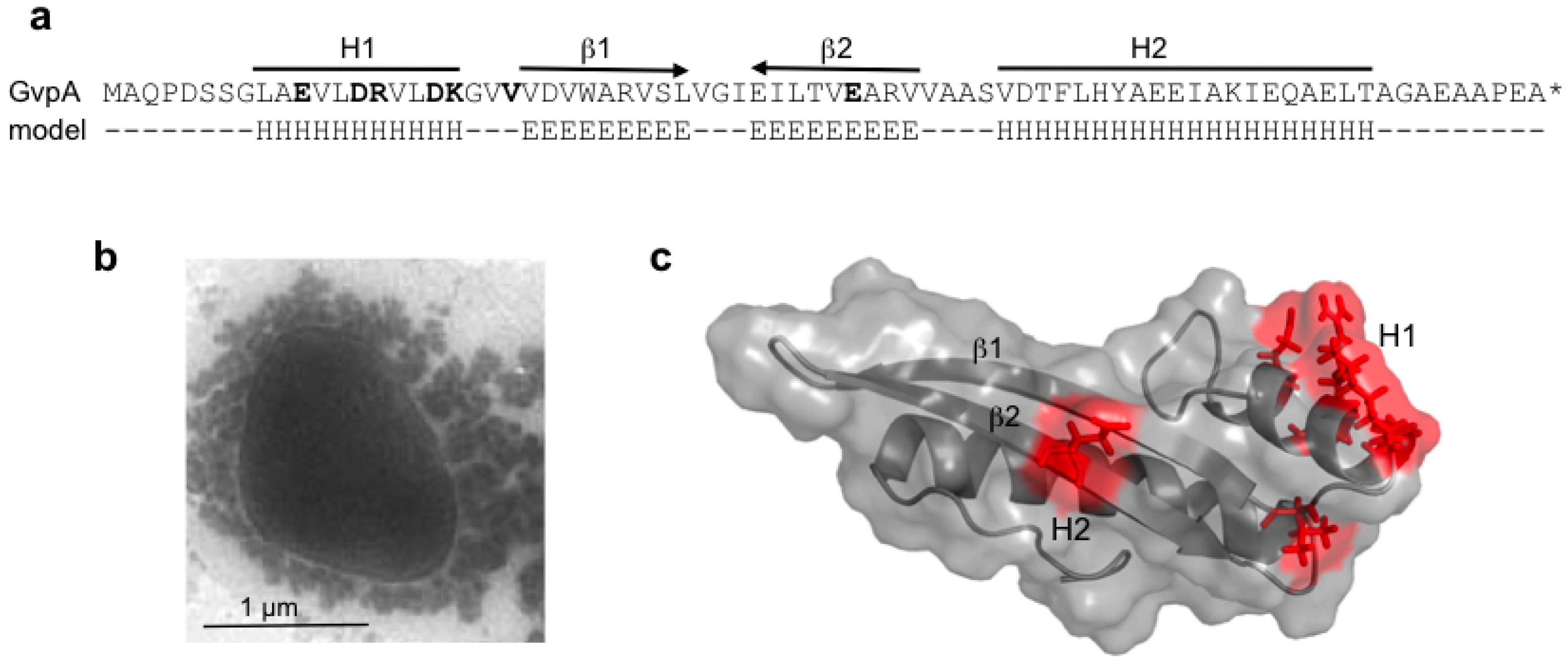
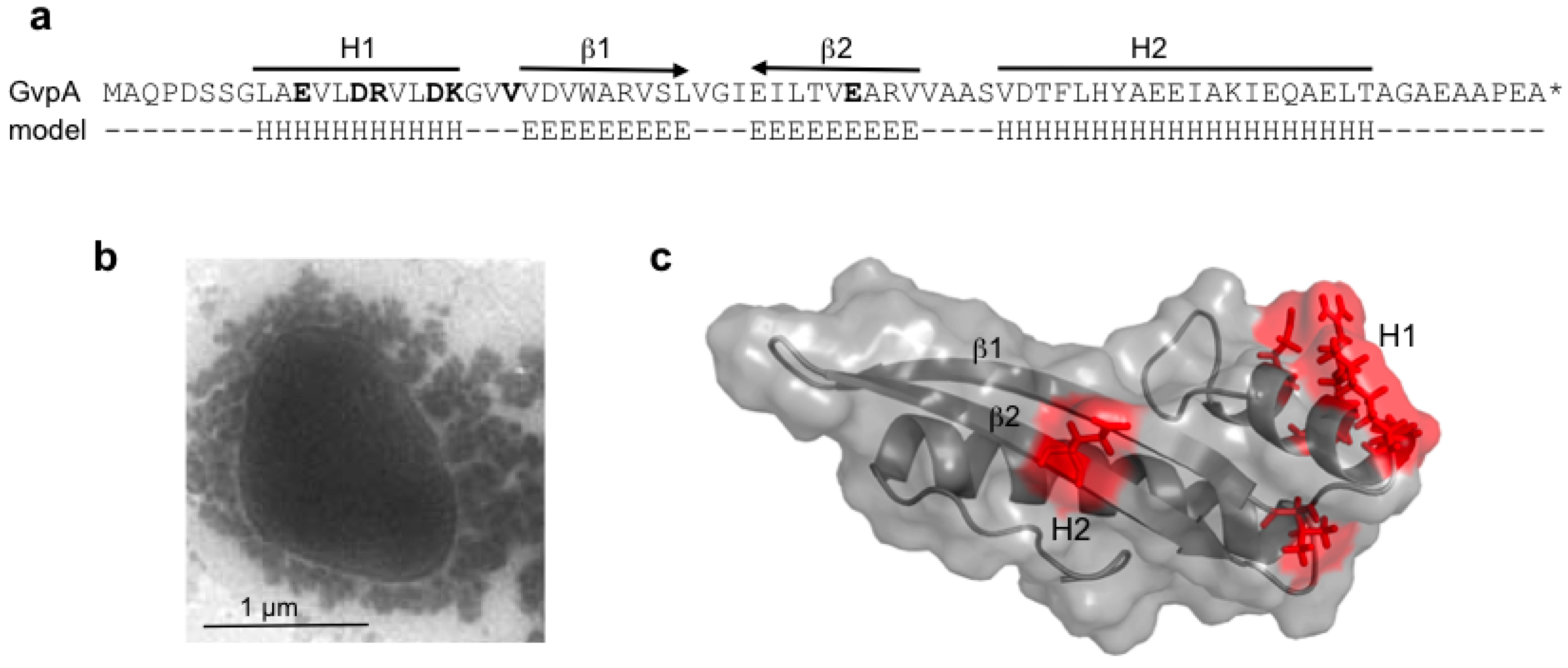
GvpA constitutes the almost crystalline gas vesicle wall, forming a helix of low pitch seen as ribs running perpendicular to the long axis by transmission electron microscopy [
58,
59]. The gas vesicle wall is difficult to disaggregate, and the protein constituents are difficult to analyze. Immunological methods and MALDI-TOF mass spectrometry indicate that with the exception of GvpK, all Gvp proteins are present [
60,
61]. The sequence of the 8-kDa GvpA is highly conserved between archaeal and also bacterial gas vesicle producers (see
Figure 4), and differences occur mainly near the N- and C-terminus. GvpA is not post-translationally modified, as demonstrated by protein sequencing and MALDI-TOF mass spectrometry [
59,
61]. A crystal structure of GvpA is not available due to its hydrophobic nature and high tendency to aggregate. The secondary structure prediction of GvpA suggests a coil-α-β-β-α-coil fold (
Figure 5a) [
62,
63]. Solid-state NMR and Fourier transform infrared spectroscopy (FTIR) with isolated gas vesicles indicates anti-parallel β-sheets, and X-ray analyses and atomic force microscopy imply that the β-strands of GvpA are tilted in the ribs at an angle of 54° [
59,
62,
64]. The C-terminal portion of GvpA is exposed to the outside of the gas vesicles, since a trypsin site and several endopeptidase GluC sites are accessible here, whereas other portions of GvpA are protected [
61].
Figure 4.
Sequence alignment of GvpA proteins derived from Haloarchaea and methanogens. Identical amino acid residues are highlighted in the following colors: red, negatively charged; blue, positively charged; green, polar, uncharged; yellow, small and variable; grey, aromatic; white, non-polar residues. The bar on top marks the highly conserved central 51 amino acids. The difference in the C-terminus of GvpA (gvp1, NRC-1) to the sequence of pGvpA (p-vac, PHH1) is due to a missing G nucleotide close to the 3'-terminus of gvpA in gvp1. pGvpA and cGvpA are derived from Hbt. salinarum PHH1 and mcGvpA from Hfx. mediterranei.
Figure 4.
Sequence alignment of GvpA proteins derived from Haloarchaea and methanogens. Identical amino acid residues are highlighted in the following colors: red, negatively charged; blue, positively charged; green, polar, uncharged; yellow, small and variable; grey, aromatic; white, non-polar residues. The bar on top marks the highly conserved central 51 amino acids. The difference in the C-terminus of GvpA (gvp1, NRC-1) to the sequence of pGvpA (p-vac, PHH1) is due to a missing G nucleotide close to the 3'-terminus of gvpA in gvp1. pGvpA and cGvpA are derived from Hbt. salinarum PHH1 and mcGvpA from Hfx. mediterranei.
A 3D model was derived from a high-performance
de novo modelling of mcGvpA [
63], and the model was confirmed for the almost identical GvpA of
Hbt. salinarum NRC-1 [
65]. The hydrophobic β-sheet portion most likely faces the gas phase and prevents the precipitation of water molecules entering the gas-filled core. A more hydrophilic portion, including the α-helix H2, constitutes the outer surface facing the cytoplasm. Single amino acid alterations in GvpA, studied in ∆A + A
mut transformants for their ability to form gas vesicles, pinpoint essential amino acid positions. Construct ∆A contains all p-vac
gvp genes, except for
gvpA, and can be complemented by
gvpA wild-type or mutant genes [
42,
63]. Some of the GvpA mutations abolish the formation of gas vesicles,
i.e., single alanine substitutions of the polar amino acids in α-helix H1 (
Figure 5b,c), whereas other mutations have no effect on gas vesicle formation [
63]. Polar residues at the surface might form salt bridges to connect GvpA-GvpA molecules or GvpA with accessory GvpX in the wall. Some GvpA mutants yield extremely long and thin gas vesicles in transformants (GvpA
I34M and others) or mini gas vesicles (GvpA
K60A and others) that are never or only slowly enlarged [
63]. The diversity of gas vesicle structures shows the dependency of the shape on the respective GvpA mutant. Such shape variants are useful to investigate the architecture of this nanocompartment.
Figure 5.
Amino acid sequence of GvpA and the structure derived from the
in silico modelling. (
a) Amino acid sequence of pGvpA and the proposed 2D structure (model). H denotes α-helices and E β-strands. The α-helices, H1 and H2, and the anti-parallel β-strands are indicated on top. (
b) Electron micrograph of a Vac
− ∆A + A
mut transformant, showing that gas-filled compartments are indeed lacking in such mutants. (
c) Structure of GvpA, highlighting some positions of ala-substitutions leading to gas vesicle negative ∆A + A
mut transformants [
63]. The respective single amino acids that are altered are shown in bold in (a).
Figure 5.
Amino acid sequence of GvpA and the structure derived from the
in silico modelling. (
a) Amino acid sequence of pGvpA and the proposed 2D structure (model). H denotes α-helices and E β-strands. The α-helices, H1 and H2, and the anti-parallel β-strands are indicated on top. (
b) Electron micrograph of a Vac
− ∆A + A
mut transformant, showing that gas-filled compartments are indeed lacking in such mutants. (
c) Structure of GvpA, highlighting some positions of ala-substitutions leading to gas vesicle negative ∆A + A
mut transformants [
63]. The respective single amino acids that are altered are shown in bold in (a).
8. Application of Gas Vesicles
Gas vesicles nanostructures are objects for special applications in biomedical research. The almost crystalline gas vesicle wall provides the structural rigidity to maintain a hollow gas-filled core that is useful as an ultrasonic contrast agent in biomedical research and clinical diagnostics [
66]. In contrast to the lipid- or protein-stabilized micro-bubbles, gas vesicles produce a stable ultrasound contrast and provide a much larger mechanical response. The acoustic scattering signal can be also silenced using ultrasound amplitudes exceeding the critical collapse pressure of the gas vesicles.
Gas vesicles are also applied as reporters for hyperpolarized xenon magnetic resonance imaging (MRI) and enable non-invasive observation of the anatomy and function of organisms [
67]. They are capable of chemical exchange saturation transfer interactions. Filled with hyperpolarized
129Xe, they allow chemically-amplified gas vesicle detection at picomolar concentrations. Functionalized gas vesicles carrying surface-attached streptavidin antibodies against the HER2 receptor are able to label an HER2-expressing breast cancer cell line, so that these cells are easily distinguishable from others by MRI [
67].
In addition, gas vesicles are an effective antigen display system based on a fusion of the peptide of interest to the surface protein, GvpC. The expression of such
gvpC-fusions in
Hbt. salinarum yields recombinant gas vesicles displaying these peptides on the surface. The recombinant gas vesicles are easy to isolate by lysis of the haloarchaeal cells and repeated centrifugally accelerated flotation. They are self-adjuvanting, and the antigenic epitopes displayed on the surface stimulate the immune system. Gas vesicles have been used to expose different SIV peptides (Tat, Rev, NefI) [
68,
69], outer membrane proteins of the pathogen,
Chlamydia trachomatis [
70], or portions of the
Salmonella SopB antigen [
71]. The antiserum raised against the
Chlamydia proteins is useful to detect the pathogen in sera of
Chlamydia-positive patients. The
Salmonella protein, SopB, is injected during the pathogenesis of typhoid, and mice boosted with SopB gas vesicles elicit a protective response to
Salmonella infection. Such recombinant gas vesicles will be further developed as improved vaccines [
71].



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}