
Influence of Low-Level Red Laser Irradiation on the Proliferation, Viability, and Differentiation of Human Embryonic Stem Cell-Derived Mesenchymal Stem Cells



Abstract
1. Introduction
2. Materials and Methods
2.1. Human Embryonic Stem Cell-Derived Mesenchymal Stem Cell Culture
2.2. Low-Level Red Laser Irradiation
2.3. Cell Proliferation Determination
2.4. Assessment of the Cell Viability Rate
2.5. Cell Migration Assay
2.6. Osteoblast Differentiation (OB) Analysis
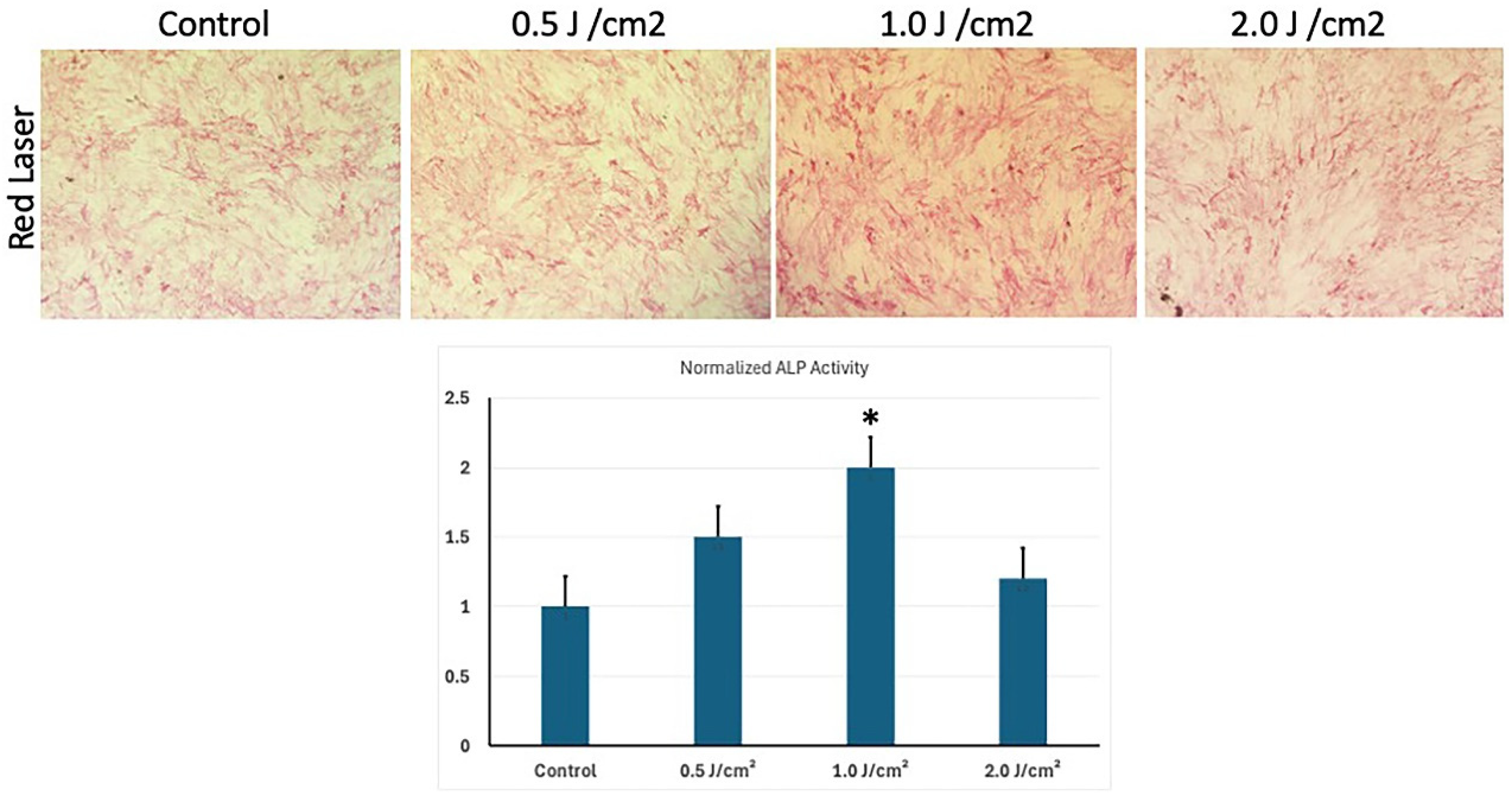
2.7. Quantitative Alkaline Phosphatase Activity
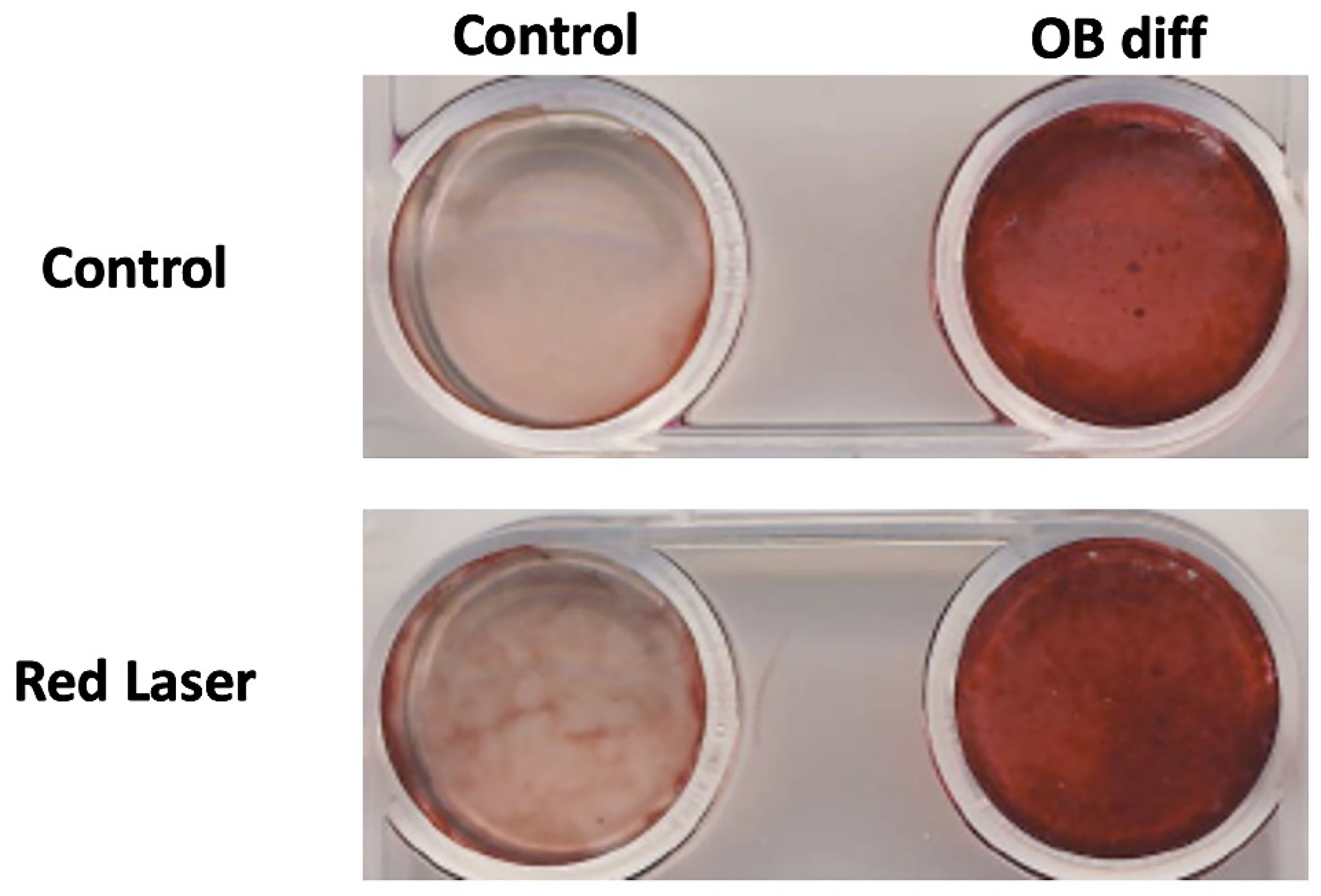
- Alizarin Red Staining:
- Image Analysis:
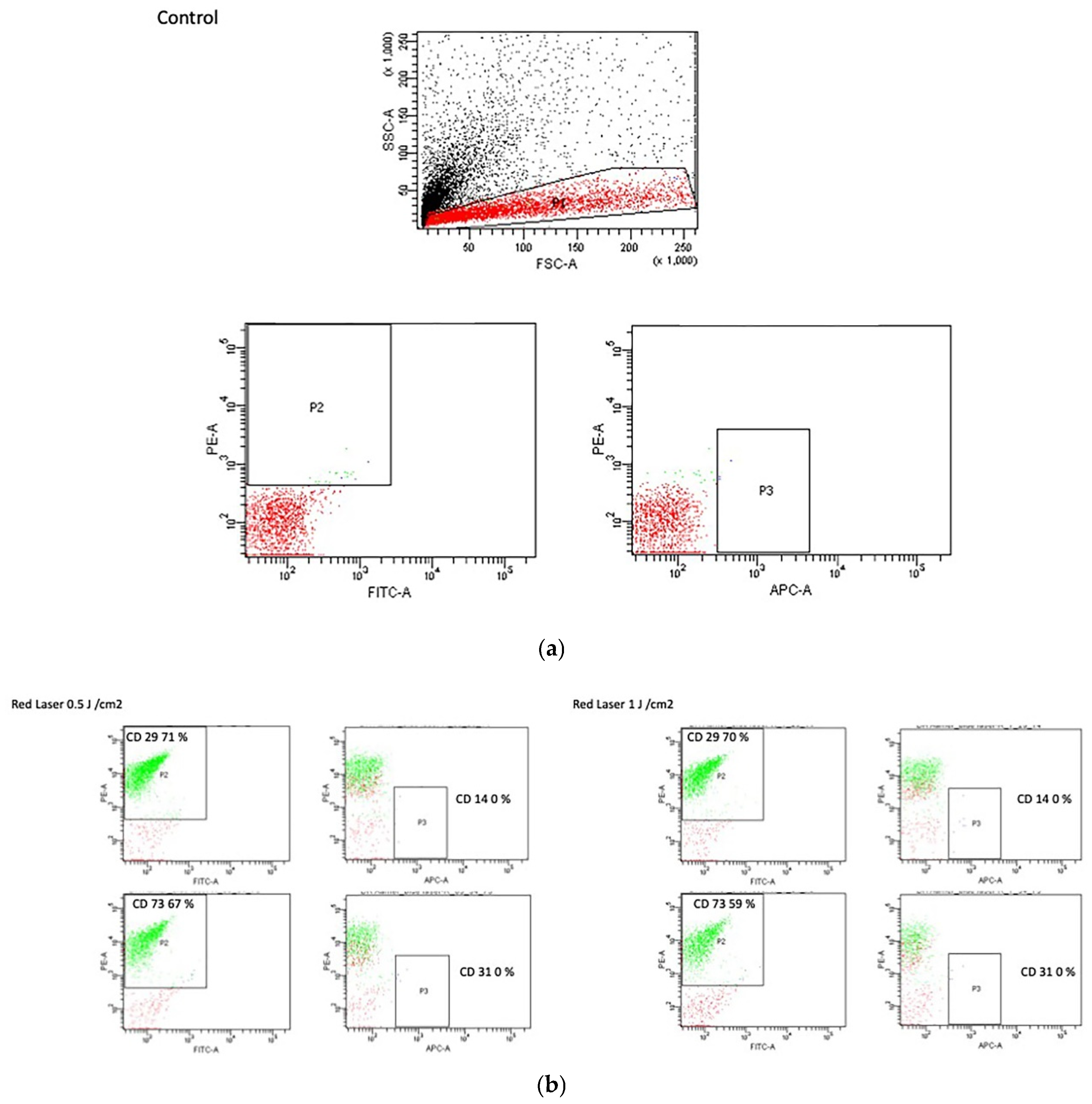
2.8. Flow Cytometry Analysis of Treated and Untreated Cells
2.9. Quantitative Real-Time PCR (qRT-PCR) Procedure
2.10. Statistical Analysis
3. Results
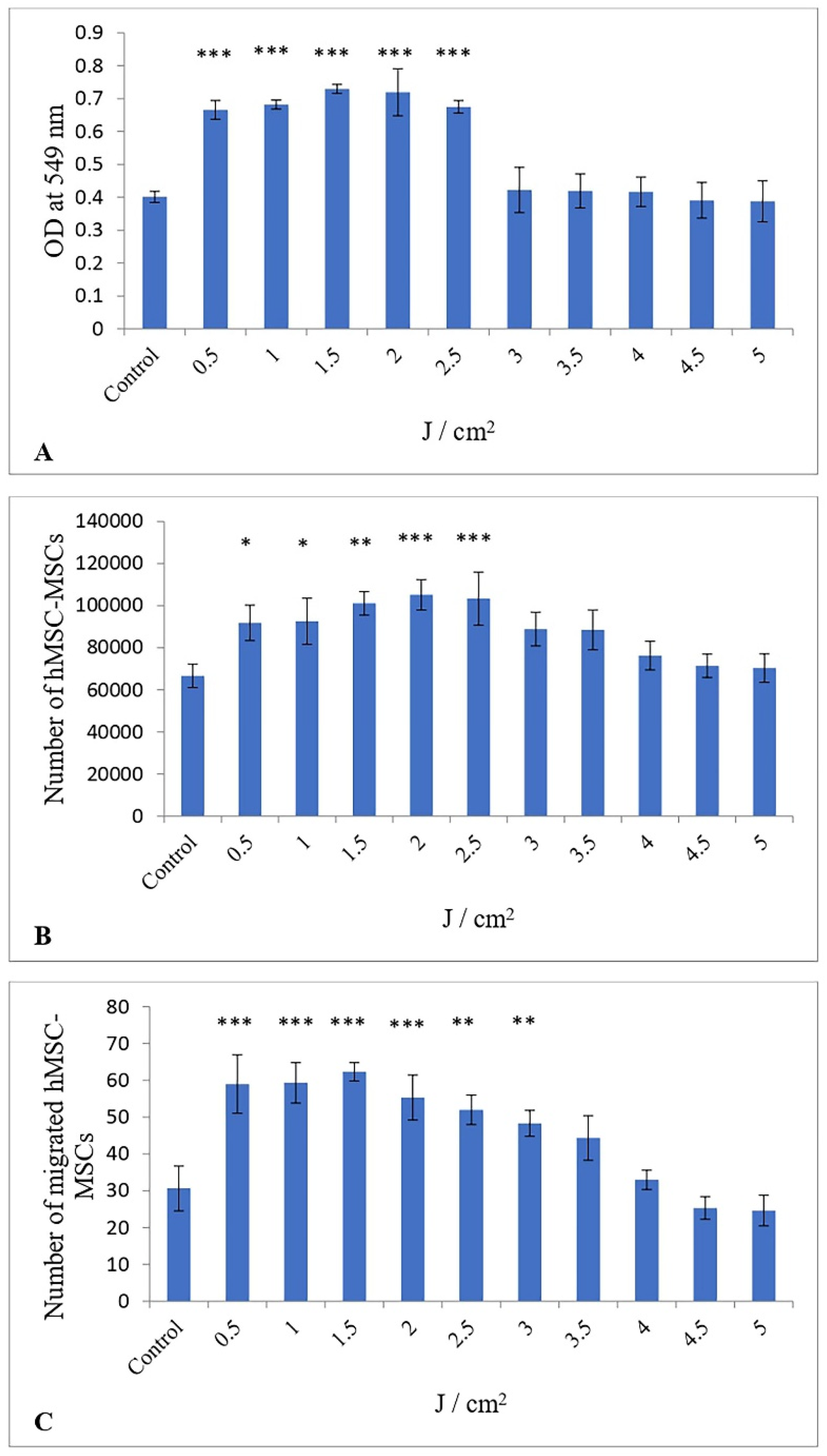
3.1. Impact of Low-Level Red Laser Treatment on the Proliferation of hESC-MSCs
3.2. Impact of Low-Level Red Laser Treatment on the Viability of hESC-MSCs
3.3. Influence of a Low-Level Red Laser on the Migration of hESC-MSCs
3.4. Low-Level Red Laser Stimulation Enhances the Osteoblast Differentiation of hESC-MSCs
3.5. Flow Cytometry Analysis
3.6. Gene Expression Analysis of Key Osteoblastic Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Woo, S.H. Advances in laser and stem cell treatment: Current technologies, limitations, and future prospects. Med. Lasers 2024, 13, 75–81. [Google Scholar] [CrossRef]
- Weiskirchen, S.; Schröder, S.K.; Buhl, E.M.; Weiskirchen, R. A Beginner’s Guide to Cell Culture: Practical Advice for Preventing Needless Problems. Cells 2023, 12, 682. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.G.; Mallon, B.S.; McKay, R.D.G.; Robey, P.G. Human Pluripotent Stem Cell Culture: Considerations for Maintenance, Expansion, and Therapeutics. Cell Stem Cell 2014, 14, 13–26. [Google Scholar] [CrossRef] [PubMed]
- Alghamdi, K.M.; Kumar, A.; Moussa, N.A. Low-level laser therapy: A useful technique for enhancing the proliferation of various cultured cells. Lasers Med. Sci. 2012, 27, 237–249. [Google Scholar] [CrossRef] [PubMed]
- Barboza, C.A.; Ginani, F.; Soares, D.M.; Henriques, A.C.G.; Freitas, R.d.A. Low-level laser irradiation induces in vitro proliferation of mesenchymal stem cells. Einstein 2014, 12, 75–81. [Google Scholar] [CrossRef]
- Liao, X.; Xie, G.H.; Liu, H.W.; Cheng, B.; Li, S.H.; Xie, S.; Xiao, L.L.; Fu, X.B. Helium-neon laser irradiation promotes the proliferation and migration of human epidermal stem cells in vitro: Proposed mechanism for enhanced wound re-epithelialization. Photomed. Laser Surg. 2014, 32, 219–225. [Google Scholar] [CrossRef]
- Cárdenas-Sandoval, R.P.; Bernal-Bernal, L.D.; Cabrera-Salazar, S.; Gómez-Ramírez, D.M.; González-Ballesteros, L.M.; Hooker-Mendoza, K.M.; Ospina-Piedrahíta, L.N.; Hernández-Charry, C.X.; Ardila-Rojas, G.; Velásquez-Durán, A.M.; et al. In-vitro study on type I collagen synthesis in low-level laser therapy on the early ligament fibroblasts’ healing process. Lasers Med. Sci. 2024, 39, 225. [Google Scholar] [CrossRef]
- Marques, N.C.T.; Neto, N.L.; Prado, M.T.O.; Vitor, L.L.R.; Oliveira, R.C.; Sakai, V.T.; Santos, C.F.; Machado, M.A.A.M.; Oliveira, T.M. Effects of PBM in different energy densities and irradiance on maintaining cell viability and proliferation of pulp fibroblasts from human primary teeth. Lasers Med. Sci. 2017, 32, 1621–1628. [Google Scholar] [CrossRef]
- Pandeshwar, P.; Roa, M.D.; Das, R.; Shastry, S.P.; Kaul, R.; Srinivasreddy, M.B. Photobiomodulation in oral medicine: A review. J. Investig. Clin. Dent. 2016, 7, 114–126. [Google Scholar] [CrossRef]
- Peng, F.; Wu, H.; Zheng, Y.; Xu, X.; Yu, J. The effect of noncoherent red light irradiation on proliferation and osteogenic differentiation of bone marrow mesenchymal stem cells. Lasers Med. Sci. 2012, 27, 645–653. [Google Scholar] [CrossRef]
- Zha, K.; Tian, Y.; Panayi, A.C.; Mi, B.; Liu, G. Recent Advances in Enhancement Strategies for Osteogenic Differentiation of Mesenchymal Stem Cells in Bone Tissue Engineering. Front. Cell Dev. Biol. 2022, 10, 824812. [Google Scholar] [CrossRef]
- Grotheer, V.; Skrynecki, N.; Oezel, L.; Windolf, J.; Grassmann, J. Osteogenic differentiation of human mesenchymal stromal cells and fibroblasts differs depending on tissue origin and replicative senescence. Sci. Rep. 2021, 11, 11968. [Google Scholar] [CrossRef]
- Sterczała, B.; Grzech-Leśniak, K.; Michel, O.; Trzeciakowski, W.; Dominiak, M.; Jurczyszyn, K. Assessment of Human Gingival Fibroblast Proliferation after Laser Stimulation In Vitro Using Different Laser Types and Wavelengths (1064, 980, 635, 450, and 405 nm)—Preliminary Report. J. Pers. Med. 2021, 11, 98. [Google Scholar] [CrossRef] [PubMed]
- Carreira, A.C.; Lojudice, F.H.; Halcsik, E.; Navarro, R.D.; Sogayar, M.C.; Granjeiro, J.M. Bone morphogenetic proteins: Facts, challenges, and future perspectives. J. Dent. Res. 2014, 93, 335–345. [Google Scholar] [CrossRef] [PubMed]
- Mahmood, A.; Harkness, L.; Schrøder, H.D.; Abdallah, B.M.; Kassem, M. Enhanced differentiation of human embryonic stem cells to multipotential mesodermal stem cells by inhibition of TGF-b/Activin/Nodal signaling using SB-431542. J. Bone Miner. Res. 2010, 25, 1216–1233. [Google Scholar] [CrossRef]
- Tuby, H.; Maltz, L.; Oron, U. Low-level laser irradiation (LLLI) promotes proliferation of mesenchymal and cardiac stem cells in culture. Lasers Surg. Med. 2007, 39, 373–378. [Google Scholar] [CrossRef]
- Hou, J.F.; Zhang, H.; Yuan, X.; Li, J.; Wei, Y.J.; Hu, S.S. In vitro effects of low-level laser irradiation for bone marrow mesenchymal stem cells: Proliferation, growth factors secretion and myogenic differentiation. Lasers Surg. Med. 2008, 40, 726–733. [Google Scholar] [CrossRef]
- Kim, H.K.; Kim, J.H.; Abbas, A.A.; Kim, D.-O.; Park, S.-J.; Chung, J.Y.; Song, E.K.; Yoon, T.R. Red light of 647 nm enhances osteogenic differentiation in mesenchymal stem cells. Lasers Med. Sci. 2009, 24, 214–222. [Google Scholar] [CrossRef]
- Alghamdi, K.M.; Kumar, A.; AlGhamdi, A.A.; Ashour, A. A comparative study of the effects of different low-level lasers on the proliferation, viability and migration of human melanocyte in vitro. Laser Med. Sci. 2015, 30, 1541–1551. [Google Scholar] [CrossRef]
- Alghamdi, K.M.; Kumar, A.; AlGhamdi, A.; Rikabi, A.; Mubarek, M.; Ashour, A. Ultra-structural effects of different low-level lasers on the normal cultured human melanocytes: In vitro comparative study. Laser Med. Sci. 2016, 31, 1819–1825. [Google Scholar] [CrossRef]
- Kumar, A.; Mohammad, A.A.; Ahmed, B.; Ennaji, H.; Ahamed, M.; Bourhia, M.; Chebaibi, M.; Benbacer, L.; Ghneim, H.K.; Abudawood, M.; et al. Apoptosis-mediated anti-proliferative activity of Calligonum comosum against human breast cancer cells, and molecular docking of its major polyphenolics to Caspase-3. Front. Cell Dev. Biol. 2022, 10, 972111. [Google Scholar] [CrossRef] [PubMed]
- AlTimimi, Z. The impact of 980 nm diode laser irradiation on the proliferation of mesenchymal stem cells derived from the umbilical cord’s. Tissue Cell 2024, 91, 102568. [Google Scholar] [CrossRef]
- Agha, R.; Beeson, W.; Beeson, W.H. In vitro comparison of light-emitting diodes and carnosic acid effects on keratinocyte proliferation and wound healing. Am. J. Cosmet. Surg. 2012, 29, 30–37. [Google Scholar] [CrossRef]
- Ma, H.; Yang, J.P.; Tan, R.K.; Lee, H.W.; Han, S.K. Effect of Low-Level Laser Therapy on Proliferation and Collagen Synthesis of Human Fibroblasts in Vitro. J. Wound Manag. Res. 2018, 14, 1–6. [Google Scholar] [CrossRef]
- Aleksic, V.; Aoki, A.; Iwasaki, K.; Takasaki, A.A.; Wang, C.Y.; Abiko, Y. Low-level Er: YAG laser irradiation enhances osteoblast proliferation through activation of MAPK/ERK. Lasers Med. Sci. 2010, 25, 559–569. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Zhu, T.; Wu, Y.; Shu, C.; Chen, Q.; Yang, J.; Luo, X.; Wang, Y. Irradiation with blue light-emitting diode enhances osteogenic differentiation of stem cells from the apical papilla. Lasers Med. Sci. 2020, 35, 1981–1988. [Google Scholar] [CrossRef]
- Rahmati, A.; Roshanak, A.; Najafi, R.; Rezaei-soufi, L.; Karkehabadi, H. Effect of diode low level laser and red-light emitting diode irradiation on cell proliferation and osteogenic/odontogenic differentiation of stem cells from the apical papilla. BMC Oral Health 2022, 22, 543. [Google Scholar] [CrossRef]
- Yamauchi, N.; Minagawa, E.; Imai, K.; Kobuchi, K.; Li, R.; Taguchi, Y.; Umeda, M. High-Intensity Red Light-Emitting Diode Irradiation Suppresses the Inflammatory Response of Human Periodontal Ligament Stem Cells by Promoting Intracellular ATP Synthesis. Life 2022, 12, 736. [Google Scholar] [CrossRef]
- Chung, H.; Dai, T.; Sharma, S.K.; Huang, Y.Y.; Carroll, J.D.; Hamblin, M.R. The nuts and bolts of low-level laser (light) therapy. Ann. Biomed. Eng. 2012, 40, 516–533. [Google Scholar] [CrossRef]
- Wang, Y.; Huang, Y.Y.; Wang, Y. Red (660 nm) or near-infrared (810 nm) photobiomodulation stimulates, while blue (415 nm), green (540 nm) light inhibits proliferation in human adipose-derived stem cells. Sci. Rep. 2017, 7, 7781. [Google Scholar] [CrossRef]
- Wang, L.; Wu, F.; Liu, C.; Song, Y.; Guo, J.; Yang, Y.; Qiu, Y. Low-level laser irradiation modulates the proliferation and the osteogenic differentiation of bone marrow mesenchymal stem cells under healthy and inflammatory condition. Lasers Med. Sci. 2019, 34, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Stein, A.; Benayahu, D.; Shainberg, A. Osteogenic differentiation in bone marrow-derived stem cells following exposure to pulsed low-intensity ultrasound and low-energy laser irradiation. J. Orthop. Res. 2005, 23, 944–952. [Google Scholar]
- Pereira, A.N.; Eduardo, C.P.; Matson, E.; Marques, M.M. Effect of low-power laser irradiation on cell growth and procollagen synthesis of cultured fibroblasts. Lasers Surg. Med. 2002, 31, 263–267. [Google Scholar] [CrossRef]
- Kovacic, J.C.; Moreno, P.; Hachinski, V.; Nabel, E.G.; Fuster, V. Cellular senescence, vascular disease, and aging: Part 2 of a 2-part review: Clinical vascular disease in the elderly. Circulation 2008, 117, 2429–2439. [Google Scholar] [CrossRef]
- Berni, M.; Brancato, A.M.; Torriani, C.; Bina, V.; Annunziata, S.; Cornella, E.; Trucchi, M.; Jannelli, E.; Mosconi, M.; Gastaldi, G.; et al. The Role of Low-Level Laser Therapy in Bone Healing: Systematic Review. Int. J. Mol. Sci. 2023, 24, 7094. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | 0.5 J/cm2 | 1.0 J/cm2 | 1.5 J/cm2 | 2.0 J/cm2 | 2.5 J/cm2 | 3.0 J/cm2 | 3.5 J/cm2 | 4.0 J/cm2 | 4.5 J/cm2 | 5.0 J/cm2 | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Proliferation (Optical Density) | |||||||||||
| Mean | 0.4 | 0.66 | 0.68 | 0.73 | 0.72 | 0.67 | 0.42 | 0.42 | 0.41 | 0.39 | 0.38 |
| Standard Deviation | 0.01 | 0.02 | 0.01 | 0.01 | 0.06 | 0.01 | 0.05 | 0.04 | 0.03 | 0.04 | 0.05 |
| Confidence Intervals | 0.39–0.4 | 0.64–0.68 | 0.67–0.69 | 0.72–0.74 | 0.65–0.78 | 0.64–0.68 | 0.36–0.48 | 0.37–0.46 | 0.37–0.46 | 0.34–0.43 | 0.32–0.43 |
| Variance | 0.0001 | 0.0005 | 0.0001 | 0.0001 | 0.004 | 0.0002 | 0.003 | 0.001 | 0.001 | 0.001 | 0.002 |
| Cohen’s d Effect size | 16.1 | ||||||||||
| Viability (no. of cells) | |||||||||||
| Mean | 66,666 | 91,851 | 92,592 | 101,110 | 105,184 | 103,332 | 88,888 | 88,517 | 76,295 | 71,481 | 70,369 |
| Standard Deviation | 4536 | 6869.1 | 8950.2 | 4536 | 5902.9 | 10,304.2 | 6541.8 | 7715.8 | 5543.4 | 4566.2 | 5543.1 |
| 95% Confidence Intervals | 61,533.4–71,799.2 | 84,078.3–99,624.2 | 82,464.3–102,720.2 | 95,977.7–106,243.5 | 98,504.9–111,864.2 | 91,672.5–114,992.6 | 81,485.7–96,290.9 | 79,786.5–97,248.7 | 70,022.7–82,568.4 | 66,313.9–76,648 | 64,096.5–76,641.5 |
| Variance | 20,575,720.2 | 47,184,910.9 | 80,107,050.9 | 20,575,720.2 | 34,844,910.8 | 106,176,790.2 | 42,795,720.2 | 59,534,293 | 30,729,766.8 | 20,850,260.6 | 30,725,816.2 |
| Cohen’s d Effect size | 4.3 | ||||||||||
| Migration | |||||||||||
| Mean | 30.6 | 59 | 59.3 | 62.3 | 55.3 | 52 | 48.3 | 44.3 | 33 | 25.3 | 24.6 |
| Standard Deviation | 4.9 | 6.4 | 4.4 | 2 | 5 | 3.2 | 2.8 | 4.9 | 2.2 | 2.5 | 3.4 |
| 95% Confidence Intervals | 25–36.1 | 51.3–66.7 | 54.3–64.3 | 60–64.5 | 49.6–60.9 | 48.4–55.6 | 45.1–51.4 | 38.7–9.8 | 30.5–35.5 | 22.4–28.1 | 20.7–28.4 |
| Variance | 24.8 | 42 | 20.2 | 4.2 | 24.8 | 10.6 | 8.2 | 24.2 | 4.6 | 6.2 | 11.5 |
| Cohen’s d Effect size | 4.8 | ||||||||||
| Control | Percentage of Total Cells |
|---|---|
| CD29 | 81.70% |
| CD14 | 0% |
| CD73 | 75.40% |
| CD34 | 0% |
| CD146 | 55.80% |
| CD31 | 0% |
| CD44 | 5.30% |
| Red Laser | 0.5 J/cm2 | 1.0 J/cm2 | 2.0 J/cm2 |
|---|---|---|---|
| CD29 | 71% | 69.70% | 65% |
| CD14 | 0% | 0% | 0% |
| CD73 | 67.20% | 59.10% | 56.40% |
| CD34 | 0% | 0% | 0% |
| CD146 | 25.60% | 31.40% | 34.50% |
| CD31 | 0% | 0% | 0% |
| CD44 | 7.50% | 5.90% | 5% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
AlGhamdi, K.M.; Kumar, A.; Alfayez, M.; Mahmood, A. Influence of Low-Level Red Laser Irradiation on the Proliferation, Viability, and Differentiation of Human Embryonic Stem Cell-Derived Mesenchymal Stem Cells. Life 2025, 15, 1125. https://doi.org/10.3390/life15071125
AlGhamdi KM, Kumar A, Alfayez M, Mahmood A. Influence of Low-Level Red Laser Irradiation on the Proliferation, Viability, and Differentiation of Human Embryonic Stem Cell-Derived Mesenchymal Stem Cells. Life. 2025; 15(7):1125. https://doi.org/10.3390/life15071125
Chicago/Turabian StyleAlGhamdi, Khalid M., Ashok Kumar, Musaad Alfayez, and Amer Mahmood. 2025. "Influence of Low-Level Red Laser Irradiation on the Proliferation, Viability, and Differentiation of Human Embryonic Stem Cell-Derived Mesenchymal Stem Cells" Life 15, no. 7: 1125. https://doi.org/10.3390/life15071125
APA StyleAlGhamdi, K. M., Kumar, A., Alfayez, M., & Mahmood, A. (2025). Influence of Low-Level Red Laser Irradiation on the Proliferation, Viability, and Differentiation of Human Embryonic Stem Cell-Derived Mesenchymal Stem Cells. Life, 15(7), 1125. https://doi.org/10.3390/life15071125