
Alterations in the Morphology of the Testis and Epididymis Caused by the Consumption of Hyperlipidic Diets in Wistar Rats

 , , ,
, , ,
Abstract
1. Introduction
2. Results
2.1. Lee Index
2.2. Scrotal Fat
2.3. Gonadosomatic Index (GSI) and Epididymal Somatic Index (ESI)
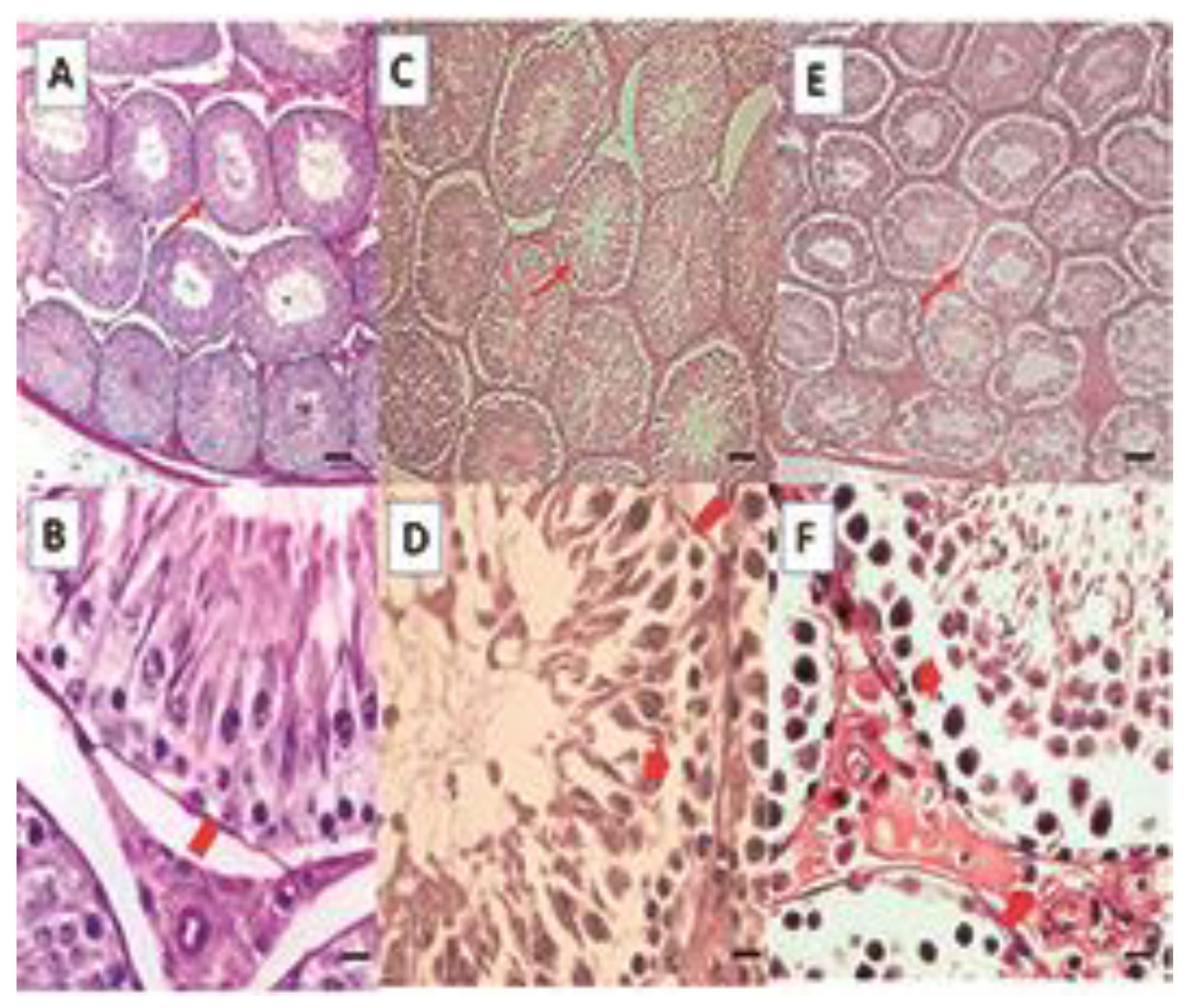
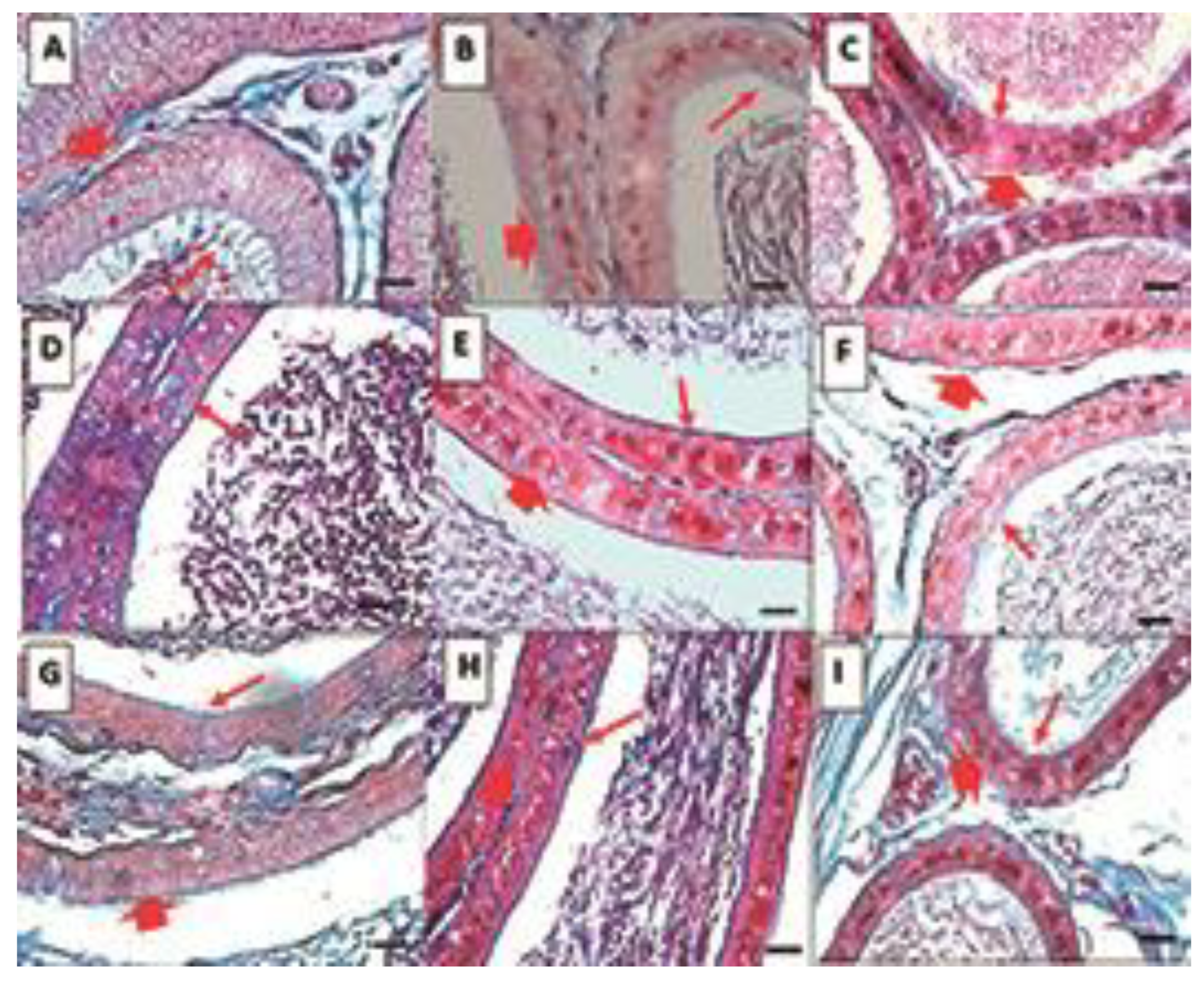
2.4. Histological Analysis
Testicle
- Epididymis
2.5. Histomorphometric Analysis
- Duct Area
- Percentage of duct lumen
- Percentage of Ductal Area Occupied by Sperm
- Epithelial Height
- Cilia length
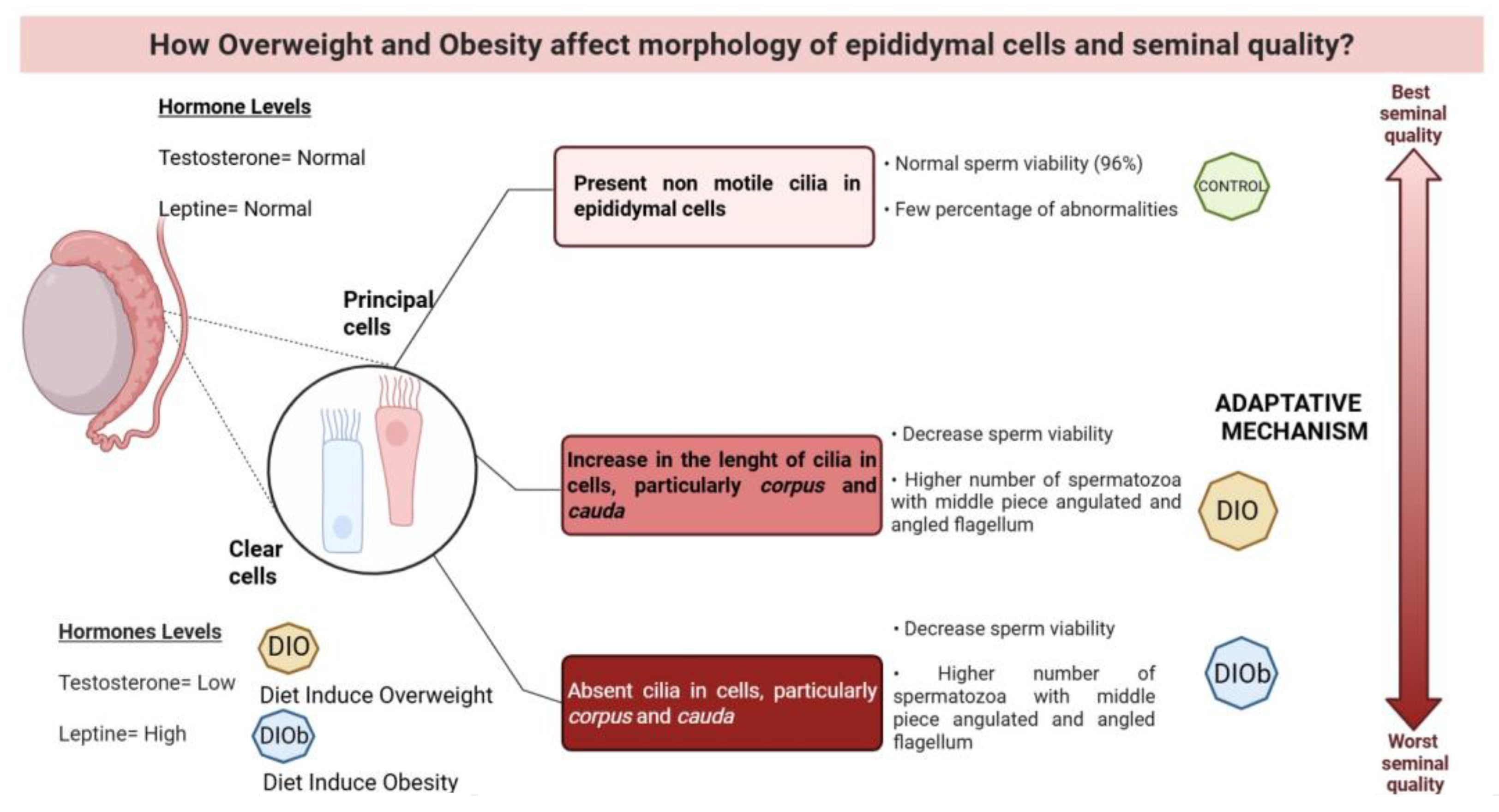
3. Discussion
4. Materials and Methods
4.1. Animals and Treatment
4.2. Lee Index
4.3. Gonadosomatic Index (GSI) and Epididymal Somatic Index (ESI)
4.4. Sample Collection
4.5. Histological Analysis
- Testicle
- Epididymis
4.6. Sperm Analysis
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| DNA | Desoxyribonucleic Acid |
| E2 | Estradiol |
| E-N | Eosin Nigrosine |
| ER | Estrogen Receptors |
| FSH | Follicle-Stimulating Hormone |
| GNRH | Hypothalamic Gonadotropin-Releasing Hormone |
| H-E | Hematoxylin–Eosin |
| KISS-1 | metastasis suppressor gene |
| LH | Luteinizing Hormone |
| SPI | Epidydimal Somatic Index |
| STI | Somatic testicular index |
| T | Testosterone |
| MT | Masson’s Trichromic |
| WHO | World Health Organization |
References
- Rastrelli, G.; Lotti, F.; Reisman, Y.; Sforza, A.; Maggi, M.; Corona, G. Metabolically healthy and unhealthy obesity in erectile dysfunction and male infertility. Expert Rev. Endocrinol. Metab. 2019, 14, 321–334. [Google Scholar] [CrossRef] [PubMed]
- Atakan, M.M.; Koşar, Ş.N.; Güzel, Y.; Tin, H.T.; Yan, X. The Role of Exercise, Diet, and Cytokines in Preventing Obesity and Improving Adipose Tissue. Nutrients 2021, 13, 1459. [Google Scholar] [CrossRef] [PubMed]
- Trayhurn, P.; Beattie, J.H. Physiological role of adipose tissue: White adipose tissue as an endocrine and secretory organ. Proc. Nutr. Soc. 2001, 60, 329–339. [Google Scholar] [CrossRef]
- Eisenberg, M.L.; Kim, S.; Chen, Z.; Sundaram, R.; Schisterman, E.F. The relationship between male BMI and waist circumference on semen quality: Data from the LIFE study. Human Reprod. 2014, 29, 193–200. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Liu, Y.; Xue, K.; Gu, G.; Fan, W.; Xu, Y.; Ding, Z. Diet-induced obesity in male C57BL/6 mice decreases fertility as a consequence of disrupted blood-testis barrier. PLoS ONE 2015, 10, e0120775. [Google Scholar] [CrossRef]
- Syriou, V.; Papanikolaou, D.; Kozyraki, A.; Goulis, D.G. Cytokines and male infertility. Eur. Cytokine Netw. 2018, 29, 73–82. [Google Scholar] [CrossRef]
- Kelly, D.M.; Jones, T.H. Testosterone and obesity. Obes. Rev. 2015, 16, 581–606. [Google Scholar] [CrossRef]
- Robaire, B.; Hamzeh, M. Androgen Action in the Epididymis. J. Androl. 2011, 32, 592–599. [Google Scholar] [CrossRef]
- O’Donnell, L.; Stanton, P.; de Kretser, D.M. Endocrinology of the Male Reproductive System and Spermatogenesis. In Endotext; De Groot, L.J., Chrousos, G., Dungan, K., Feingold, K.R., Grossman, A., Hershman, J.M., Koch, C., Korbonits, M., McLachlan, R., New, M., et al., Eds.; MDText.com, Inc.: South Dartmouth, MA, USA, 2017. [Google Scholar]
- Robaire, B.; Hinton, B.T. The Epididymis. In Knobil and Neill’s Physiology of Reproduction; Elsevier: San Diego, CA, USA, 2015; pp. 691–771. [Google Scholar]
- Robaire, B.; Viger, R.S. Regulation of Epididymal Epithelial Cell Functions. Biol. Reprod. 1995, 52, 226–236. [Google Scholar] [CrossRef]
- Robaire, B.; Seenundun, S.; Hamzeh, M.; Lamour, S.-A. Androgenic regulation of novel genes in the epididymis. Asian J. Androl. 2007, 9, 545–553. [Google Scholar] [CrossRef]
- Dacheux, J.-L.; Dacheux, F. New insights into epididymal function in relation to sperm maturation. Reproduction 2013, 147, R27–R42. [Google Scholar] [CrossRef]
- Tekpetey, F.R.; Veeramachaneni, D.N.R.; Amann, R.P. Localization of androgen receptors in ram epididymal principal cells. Reproduction 1989, 87, 311–319. [Google Scholar] [CrossRef] [PubMed]
- Olivera, M.; Ruiz, T.; Tarazona, A.; Giraldo, C. El espermatozoide, desde la eyaculación hasta la fertilización. Rev. Colomb. Cienc. Pecuarias 2006, 19, 426–436. [Google Scholar] [CrossRef]
- Posadas, R.J. Efecto del Sobrepeso y la Obesidad en la Actividad de las Enzimas Antioxidantes en Testículo y Epidídimo de Rata Wistar. Master’s Thesis, Universidad Autónoma Metropolitana Iztapalapa, Ciudad de México, Mexico, 2020. [Google Scholar] [CrossRef]
- Turner, T.T.; Johnston, D.S.; Jelinsky, S.A.; Tomsig, J.L.; Finger, J.N. Segment boundaries of the adult rat epididymis limit interstitial signaling by potential paracrine factors and segments lose differential gene expression after efferent duct ligation. Asian J. Androl. 2007, 9, 565–573. [Google Scholar] [CrossRef]
- Bellastella, G.; Menafra, D.; Puliani, G.; Colao, A.; Savastano, S. Obesity Programs of nutrition, Education, Research and Assessment (OPERA) Group. How much does obesity affect the male reproductive function? Int. J. Obes. Suppl. 2019, 9, 50–64. [Google Scholar] [CrossRef] [PubMed]
- De Grava, K.W.; Klinefelter, G.R. Interpreting histopathology in the epididymis. Spermatogenesis 2014, 4, e979114. [Google Scholar] [CrossRef]
- Vigueras, V.R.M.; Rojas, C.J.C.; Chávez, S.M.; Gutiérrez, P.O.; García, C.M.E.; Cuevas, A.O.; Reyes, R.M.M.; Zambrano, E. Alterations in the spermatic function generated by obesity in rats. Acta Histochem. 2011, 113, 214–220. [Google Scholar] [CrossRef]
- Teerds, K.J.; de Rooij, D.G.; Keijer, J. Functional relationship between obesity and male reproduction: From humans to animal models. Hum. Reprod. Update 2011, 17, 667–683. [Google Scholar] [CrossRef] [PubMed]
- Sengupta, P.; Dutta, S. Thyroid Disorders and Semen Quality. Biomed. Pharmacol. J. 2018, 11, 1–10. [Google Scholar] [CrossRef]
- Molinar, T.E.M. Estudio del Efecto de los Ácidos Grasos Poliinsaturados, Polifenoles e Iminociclitoles Sobre Marcadores Relacionados con el Síndrome Metabólico. Ph.D. Thesis, Departamento de Bioquímica y Biología Molecular, Universidad de Barcelona, Barcelona, Spain, 2015. [Google Scholar]
- Novelli, E.L.B.; Diniz, Y.S.; Galhardi, C.M.; Ebaid, G.M.X.; Rodrigues, H.G.; Mani, F.; Fernandes, A.A.H.; Cicogna, A.C.; Novelli, F. Anthropometrical parameters and markers of obesity in rats. Lab. Anim. 2007, 41, 111–119. [Google Scholar] [CrossRef]
- Jeyakumar, S.M.; Lopamudra, P.; Padmini, S.; Balakrishna, N.; Giridharan, N.V.; Vajreswari, A. Fatty acid desaturation index correlates with body mass and adiposity indices of obesity in Wistar NIN obese mutant rat strains WNIN/Ob and WNIN/GR-Ob. BioMed Cent. 2009, 6, 27. [Google Scholar] [CrossRef] [PubMed]
- Traish, A.M.; Guay, A.; Feeley, R.; Saad, F. The Dark Side of Testosterone Deficiency: I. Metabolic Syndrome and Erectile Dysfunction. J. Androl. 2008, 30, 10–22. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Valderrama, L.; Mendoza-Sánchez, J.E.; Rodríguez-Tobón, E.; Arrieta-Cruz, I.; González-Márquez, H.; Salame-Méndez, P.A.; Tarragó-Castellanos, R.; Cortés-Barberena, E.; Rodríguez-Tobón, A.; Arenas-Ríos, E. High-Fat Diets Disturb Rat Epididymal Sperm Maturation. Int. J. Mol. Sci. 2025, 26, 1850. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Suleiman, J.B.; Nna, V.U.; Othman, Z.A.; Zakaria, Z.; Bakar, A.B.A.; Mohamed, M. Orlistat attenuates obesity-induced decline in steroidogenesis and spermatogenesis by up-regulating steroidogenic genes. Andrology 2020, 8, 1471–1485. [Google Scholar] [CrossRef] [PubMed]
- Du Plessis, S.S.; Cabler, S.; McAlister, D.A.; Sabanegh, E.; Agarwal, A. The effect of obesity on sperm disorders and male infertility. Nat. Rev. Urol. 2010, 7, 153–161. [Google Scholar] [CrossRef]
- Chughtai, B.; Lee, R.K.; Te, A.E.; Kaplan, S.A. Metabolic syndrome and sexual dysfunction. Curr. Opin. Urol. 2011, 21, 514–518. [Google Scholar] [CrossRef]
- Tena, S.M.; Barreiro, M.L. Leptin in Male Reproduction: The Testis Paradigm. Mol. Cell. Endocrinol. 2002, 188, 9–13. [Google Scholar] [CrossRef]
- Zhao, J.; Zhai, L.; Liu, Z.; Wu, S.; Xu, L. Leptin Level and Oxidative Stress Contribute to Obesity-Induced Low Testosterone in Murine Testicular Tissue. Oxid. Med. Cell. Long. 2014, 2014, 190945. [Google Scholar] [CrossRef]
- Donner, D.G.; Elliott, G.E.; Beck, B.R.; Bulmer, A.C.; Du, T.E.F. Impact of Diet-Induced Obesity and Testosterone Deficiency on the Cardiovascular System: A Novel Rodent Model Representative of Males with Testosterone-Deficient Metabolic Syndrome (TDMetS). PLoS ONE 2015, 10, e0138019. [Google Scholar] [CrossRef]
- Varga, T.; Czimmerer, Z.; Nagy, L. PPARs are a unique set of fatty acid regulated transcription factors controlling both lipid metabolism and inflammation. Biochim. Biophys. Acta (BBA)-Mol. Basis Dis. 2011, 1812, 1007–1022. [Google Scholar] [CrossRef]
- Michalakis, K.; Mintziori, G.; Kaprara, A.; Tarlatzis, B.C.; Goulis, D.G. The complex interaction between obesity, metabolic syndrome and reproductive axis: A narrative review. Metabolism 2013, 62, 457–478. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, V.L.; Posadas, R.J.; Bonilla, J.H.; Tarragó, C.M.R.; González, M.H.; Arrieta, C.I.; González, N.L.; Salame, M.A.; Rodríguez, T.A.; Morales, M.J.G.; et al. Sperm dysfunction in the testes and epididymes due to overweight and obesity is not caused by oxidative stress. Int. J. Endocrinol. 2022, 2022, 3734572. [Google Scholar] [CrossRef]
- Champroux, A.; Torres-Carreira, J.; Gharagozloo, P.; Drevet, J.R.; Kocer, A. Mammalian sperm nuclear organization: Resiliencies and vulnerabilities. Basic Clin. Androl. 2016, 26, 17. [Google Scholar] [CrossRef]
- Drevet, J.R.; Aitken, R.J. Oxidation of sperm nucleus in mammals: A physiological necessity to some extent with adverse impacts on oocyte and offspring. Antioxidants 2020, 9, 95. [Google Scholar] [CrossRef]
- Rodríguez, E.; Fierro, R.; Gonz’alez, H.; Garc’ıa, F.; Arenas, R. Boar sperm incubation with reduced glutathione (GSH) diferentially modulates protein tyrosine phosphorylation patterns and reorganization of calcium in sperm, in vitro fertilization, and embryo development depending on concentrations. Res. Vet. Sci. 2020, 135, 386–396. [Google Scholar] [CrossRef]
- Genchi, V.A.; Rossi, E.; Lauriola, C.; Dória, R.; Palma, G.; Borrelli, A.; Caccioppoli, C.; Giogino, F.; Cignarelli, A. Adipose tissue dysfuntion and obesity related male hypogonadism. Int. J. Mol. Sci. 2022, 23, 8194. [Google Scholar] [CrossRef]
- Ishikawa, T.; Fujioka, H.; Ishimura, T.; Takenaka, A.; Fujisawa, M. Expression of Leptin and Leptin Receptor in the Testis of Fertile and Infertile Patients. Andrologia 2007, 39, 22–27. [Google Scholar] [CrossRef]
- Hess, R.A. Estrogen in the adult male reproductive tract: A review. Reprod. Biol. Endocrinol. 2003, 1, 52. [Google Scholar] [CrossRef] [PubMed]
- Moore, H.D.M.; Bedford, J.M. Short-term effects of androgen withdrawal on the structure of different epithelial cells in the rat epididymis. Anat. Rec. 1979, 193, 293–311. [Google Scholar] [CrossRef]
- Murakami, M.; Shimada, T.; Noda, S.; Kawano, T. Effects of castration and androgen-substitution on the morphology of the epididymal epithelium of the Japanese monkey, Macacus fuscatus, as revealed by scanning electron microscopy. Cell Tissue Res. 1976, 170, 515–521. [Google Scholar] [CrossRef] [PubMed]
- Turner, T.T.; Bang, H.J.; Attipoe, S.A.; Johnston, D.S.; Tomsig, J.L. Sonic hedgehog pathway inhibition alters epididymal function as assessed by the development of sperm motility. J. Androl. 2006, 27, 225–232. [Google Scholar] [CrossRef] [PubMed]
- Girardet, L.; Augiere, C.; Asselin, M.P.; Belleannée, C. Primary cilia: Biosensors of the male reproductive tract. Andrology 2019, 7, 588–602. [Google Scholar] [CrossRef] [PubMed]
- Dandona, P.; Aljada, A.; Chaudhuri, A.; Mohanty, P.; Garg, R. Metabolic Syndrome: A Comprehensive Perspective Based on Interactions Between Obesity, Diabetes, and Inflammation. Circulation 2005, 111, 1448–1454. [Google Scholar] [CrossRef]
- Waites, G.M.H.; Zorgniotti, A.W. (Eds.) Thermoregulation of the Scrotum and Testis: Studies in Animals and Significance for Man. In Temperature and Environmental Effects on the Testis; Springer: Boston, MA, USA, 1991; pp. 9–17. [Google Scholar]
- Rivas, C.; Flores, M.; Pérez, J.; Galelgos, E.; Cárdenas, M.; Ayala, M.E.; Aragón, A. Acute effects of para-chloroamphetamine on testosterone and markers of apoptosis in seminiferous epithelium of prepubertal male rats. Syst. Biol. Rep. Med. 2022, 68, 396–406. [Google Scholar] [CrossRef]
- Merve, A.E.; Arbak, S.; Ercan, F. Ameliorating effects of exercise on disrupted epididymal sperm parameters in high fat diet-induced obese rats. Mar. Med. J. 2019, 32, 14–19. [Google Scholar] [CrossRef]
- Alves, M.G.; Jesus, T.T.; Sousa, M.; Goldberg, E.; Silva, B.M.; Oliveira, P.F. Male fertility and obesity: Are ghrelin, leptin and glucagon-like peptide-1 pharmacologically relevant? Curr. Pharm. Des. 2016, 22, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Bernardis, L.L.; Patterson, B.D. Correlation between ‘Lee index’ and carcass fat content in weanling and adult female rats with hypothalamic lesions. J. Endocrinol. 1968, 40, 527–528. [Google Scholar] [CrossRef] [PubMed]
- León-Galván, M.A.; Fonseca, T.; López-Wilchis, R.; Rosado, A. Prolonged storage of spermatozoa in the genital tract of female Mexican big-eared bats (Corynorhinus mexicanus): The role of lipid peroxidation. Can. J. Zool. 1999, 77, 7–12. [Google Scholar] [CrossRef]
- World Health Organization. WHO Laboratory Manual for the Examination and Processing of Human Semen, 6th ed.; World Health Organization: Geneva, Switzerland, 2010; pp. 1–287. [Google Scholar]
- Lucio, R.A.; Tlachi, J.L.; López, A.A.; Zempoalteca, R.; Velázquez-Moctezuma, J. Análisis de los parámetros del eyaculado en la rata Wistar de laboratorio: Descripción de la técnica. Vet. México 2009, 40, 405–415. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
| Control | DIO | DIOb | |
|---|---|---|---|
| Lee Index (%) | 0.291 | 0.32 | 0.33 |
| Scrotal fat (g) | 3.75 ± 0.87 a | 6.61 ± 0.42 b | 13.31 ± 4.48 c |
| GSI (%) | 0.94 ± 0.04 a | 0.98 ± 0.03 a | 0.80 ± 0.03 b |
| ESI (%) | 0.36 ± 0.29 a | 0.35 ± 0.027 a | 0.80 ± 0.0017 b |
| Control | DIO | DIOb | |
|---|---|---|---|
| Seminiferous tubule diameter (µm) | 306.21 ± 33.37 d | 310.13 ± 36.82 d | 333.23 ± 25.59 e |
| Height of the seminiferous epithelium (µm) | 75.58 ± 14.46 d | 66.37 ± 10.07 e | 82.93 ± 12.37 f |
| Control | DIO | DIOb | ||
|---|---|---|---|---|
| Duct area (µm2) | Caput | 10,953 ± 540.3 d | 22,911 ± 540.3 e | 27,172 ± 540.3 e |
| Corpus | 27,866 ± 523.29 d | 25,189 ± 523.29 e | 21,723 ± 523.29 e | |
| Cauda | 28,770.23 ± 489.4 d | 24,149.41 ± 489.4 e | 25,149 ± 489.4 e | |
| Duct lumen (%) | Caput | 42.3 ± 0.55 d | 66.4 ± 0.55 e | 64.8 ± 0.55 e |
| Corpus | 67.4 ± 0.49 d | 65.3 ± 0.49 e | 67.6 ± 0.02 d | |
| Cauda | 70.83 ± 0.27 d | 67.71 ± 0.27 e | 68.8 ± 0.27 e | |
| Area of the ducts occupied by sperm (%) | Caput | 15.5 ± 0.46 d | 42.2 ± 0.46 e | 48.44 ± 0.46 e |
| Corpus | 44.94 ± 0.48 | 43.87 ± 0.48 | 44.1 ± 0.48 | |
| Cauda | 44.05 ± 0.38 d | 45.2 ± 0.38 d | 52.49 ± 0.38 e | |
| Epithelial height (µm) | Caput | 20.5 + 0.2 d | 15.66 ± 0.13 e | 14.55 ± 0.13 e |
| Corpus | 16.9 ± 0.16 d | 15.8 ± 0.16 e | 15.3 ± 0.16 e | |
| Cauda | 13.9 ± 0.13 d | 15.8 ± 0.16 e | 13.35 ± 0.13 d |
| Control | DIO | DIOb | |
|---|---|---|---|
| Caput (µm) | 142.24 ± 21.35 d | 40.24 ± 21.35 e | 10.09 ± 21.35 f |
| Corpus (µm) | 52.94 ± 15. 24 d | 71.02 ± 15.24 e | 15.89 ± 15.24 f |
| Cauda (µm) | 21.91 ± 17. 88 d | 102.50 ± 17.88 e | 55.02 ± 17.88 f |
| Control | DIO | DIOb | |
|---|---|---|---|
| Vitality (%) | 96 ± 3.1 d | 70 ± 3.1 e | 32 ± 3.1 f |
| Abnormalities (%) | |||
| Amorphous head | 1 ± 0.2 | 2 ± 0.2 | 3 ± 0.2 |
| Middle piece | |||
| angulated | 7 ± 1.9 d | 19 ± 1.9 e | 34 ± 1.9 f |
| and asymmetric | 0 | 1 ± 1.02 | 1 ± 0.2 |
| Flagellum | |||
| Angled | 13 ± 3 d | 45 ± 3 e | 52 ± 3 f |
| Coiled | 2 ± 3.4 d | 14 ± 3.4 e | 23 ± 3.4 f |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tarragó Castellanos, R.; Mendoza Delgado, M.; Ruiz Valderrama, L.; Arrieta Cruz, I.; Cortés Barberena, E.; Morales Otal, A.; Rodríguez Tobón, E.; Rodríguez Tobón, A.; Olvera Ramírez, J.; Arenas Ríos, E. Alterations in the Morphology of the Testis and Epididymis Caused by the Consumption of Hyperlipidic Diets in Wistar Rats. Life 2025, 15, 959. https://doi.org/10.3390/life15060959
Tarragó Castellanos R, Mendoza Delgado M, Ruiz Valderrama L, Arrieta Cruz I, Cortés Barberena E, Morales Otal A, Rodríguez Tobón E, Rodríguez Tobón A, Olvera Ramírez J, Arenas Ríos E. Alterations in the Morphology of the Testis and Epididymis Caused by the Consumption of Hyperlipidic Diets in Wistar Rats. Life. 2025; 15(6):959. https://doi.org/10.3390/life15060959
Chicago/Turabian StyleTarragó Castellanos, Rosario, Maria Mendoza Delgado, Lorena Ruiz Valderrama, Isabel Arrieta Cruz, Edith Cortés Barberena, Adriana Morales Otal, Ernesto Rodríguez Tobón, Ahiezer Rodríguez Tobón, Javier Olvera Ramírez, and Edith Arenas Ríos. 2025. "Alterations in the Morphology of the Testis and Epididymis Caused by the Consumption of Hyperlipidic Diets in Wistar Rats" Life 15, no. 6: 959. https://doi.org/10.3390/life15060959
APA StyleTarragó Castellanos, R., Mendoza Delgado, M., Ruiz Valderrama, L., Arrieta Cruz, I., Cortés Barberena, E., Morales Otal, A., Rodríguez Tobón, E., Rodríguez Tobón, A., Olvera Ramírez, J., & Arenas Ríos, E. (2025). Alterations in the Morphology of the Testis and Epididymis Caused by the Consumption of Hyperlipidic Diets in Wistar Rats. Life, 15(6), 959. https://doi.org/10.3390/life15060959