
The Role of the Gut Microbiota in Female Reproductive and Gynecological Health: Insights into Endometrial Signaling Pathways




Abstract
1. Introduction
2. Determinants of Gut Microbial Composition and Its Implications for Endometrial Function
2.1. Microbial Trajectories Across the Lifespan: Impact on Reproductive Immunity and Estrogen Homeostasis
2.2. Environmental Modulators of the Gut Microbiota and Their Influence on Systemic Metabolic and Immune Responses
2.3. Host Genetic Control of Gut Microbiota: Relevance for Endometrial Signaling Pathways
3. The Gut Microbiome as an Endocrine Modulator of Reproductive Signaling
3.1. Estrobolome-Driven Estrogen Recycling: Molecular Links to Endometrial Receptivity
3.2. Microbial Modulation of Metabolic Hormones: Downstream Effects on Implantation and Uterine Function
4. Microbiota-Mediated Mechanisms Underlying Endometrial Dysfunction in Gynecological Disorders
4.1. Endometriosis Pathogenesis Through Gut-Driven Inflammatory and Estrogenic Dysregulation
4.2. Gut Microbial Dysbiosis in PCOS: Crosstalk Between Inflammation, Hormonal Imbalance, and Endometrial Disruption
4.3. Actions of Gut–Immune–Endometrial Axis in Recurrent Implantation Failure (RIF) and Recurrent Pregnancy Loss (RPL)
- (1)
- The disruption of microbial diversity and ecological stability;
- (2)
- The skewing of immune cell polarization and cytokine networks;
- (3)
- The dysregulation of microbiota-derived metabolites crucial for immune tolerance;
- (4)
- Immunogenetic susceptibility, which is mediated by molecular mimicry and autoimmune activation.
4.3.1. Microbial Diversity Collapse and Taxonomic Shifts in RPL and RIF
4.3.2. Microbiota-Driven Autoimmunity and Molecular Mimicry in Endometrial Rejection
4.4. Gut Microbial Dysbiosis in Preterm Birth: Immune Activation and Barrier Dysfunction at the Maternal–Fetal Interface
4.5. Gut-Endometrial Crosstalk in Preeclampsia: Microbial Influences on Vascular Inflammation and Placental Signaling
5. Microbial Dysbiosis Beyond Classical Gynecological Disorders: Endocrine–Immune Disruption and Endometrial Signaling
5.1. Bacterial Vaginosis and the Gut–Vaginal Axis: Microbial Crosstalk and Endometrial Consequences
5.2. Uterine Fibroids: Estrogen Dysregulation and Immune Modulation Mediated by the Gut Microbiota
5.3. Gynecologic Cancers: Gut Microbiota, Inflammation, and Hormone-Driven Oncogenesis
5.4. Gut Dysbiosis-Driven Immune Priming and Hormonal Imbalance in Reproductive Dysfunction
6. Gut Microbiota and Endometrial Biology in Reproductive Function: Mechanistic Insights and Future Perspectives
- (1)
- Immune dysregulation, characterized by elevated IL-6, TNF-α, and IL-1β; increased Th17 cells; and reduced Tregs [118];
- (2)
- The activation of inflammatory signaling pathways, including NF-κB, TLR4–LPS, and NLRP3 inflammasome [118];
- (3)
- (4)
- (5)
- (6)
- Maladaptive microbial–endometrial crosstalk involving translocated microbial metabolites, local immune activation, and shifts in the endometrial microbial community [74].
- (1)
- The mechanistic dissection of SCFA- and tryptophan-mediated gene regulation;
- (2)
- An investigation into the inflammatory and fibrotic consequences of microbial DNA and viable microbe translocation;
- (3)
- The identification of reproducible microbial and metabolic biomarkers across fertility-relevant conditions;
- (4)
- The development of precision microbiota-based therapies tailored to specific immunoendocrine profiles.
7. Conclusions: Integrating Microbial, Immune, and Hormonal Networks in Fertility Research
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AhR | Aryl hydrocarbon receptor |
| BAs | Bile acids |
| BV | Bacterial vaginosis |
| CRP | C-reactive protein |
| EMT | Epithelial–mesenchymal transition |
| EDCs | Endocrine-disrupting chemicals |
| FGT | Female genital tract |
| FMT | Fecal microbiota transplantation |
| FXR | Farnesoid X receptor |
| GPBAR1 | G protein-coupled bile acid receptor 1 (also known as TGR5) |
| GPCR | G protein-coupled receptor |
| GLP-1 | Glucagon-like peptide-1 |
| HPG axis | Hypothalamic–pituitary–gonadal axis |
| HDAC | Histone deacetylase |
| IL | Interleukin |
| IPA | Indolepropionic acid |
| IVF | In vitro fertilization |
| KP | Kynurenine pathway |
| LPS | Lipopolysaccharide |
| MMT | Mesothelial–mesenchymal transition |
| MR | Mendelian randomization |
| NF-κB | Nuclear factor kappa-light-chain-enhancer of activated B cells |
| NK cells | Natural killer cells |
| PCOS | Polycystic ovary syndrome |
| PE | Preeclampsia |
| PID | Pelvic inflammatory disease |
| PYY | Peptide YY |
| RIF | Recurrent implantation failure |
| RPL | Recurrent pregnancy loss |
| ROS | Reactive oxygen species |
| SCFA | Short-chain fatty acid |
| SRC-1 | Steroid receptor coactivator-1 |
| TAGLN | Transgelin |
| Th | T helper (cell) |
| TGF-β | Transforming growth factor beta |
| TLR | Toll-like receptor |
| TNF-α | Tumor necrosis factor alpha |
| Treg | Regulatory T cell |
| ZO-1 | Zonula occludens-1 |
References
- Pérez-Prieto, I.; Rodríguez-Santisteban, A.; Altmäe, S. Beyond the Reproductive Tract: Gut Microbiome and Its Influence on Gynecological Health. Curr. Opin. Clin. Nutr. Metab. Care 2024, 36, 134–147. [Google Scholar] [CrossRef] [PubMed]
- Elkafas, H.; Walls, M.; Al-Hendy, A.; Ismail, N. Gut and Genital Tract Microbiomes: Dysbiosis and Link to Gynecological Disorders. Front. Cell Infect. Microbiol. 2022, 12, 1059825. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.S.B.; Giacobini, P. Don’t Trust Your Gut: When Gut Microbiota Disrupt Fertility. Cell Metab. 2019, 30, 616–618. [Google Scholar] [CrossRef] [PubMed]
- Baker, J.M.; Al-Nakkash, L.; Herbst-Kralovetz, M.M. Estrogen-Gut Microbiome Axis: Physiological and Clinical Implications. Maturitas 2017, 103, 45–53. [Google Scholar] [CrossRef]
- Yu, W.; Sun, S.; Fu, Q. The Role of Short-Chain Fatty Acid in Metabolic Syndrome and Its Complications: Focusing on Immunity and Inflammation. Front. Immunol. 2025, 16, 1519925. [Google Scholar] [CrossRef]
- Konopelski, P.; Mogilnicka, I. Biological Effects of Indole-3-Propionic Acid, a Gut Microbiota-Derived Metabolite, and Its Precursor Tryptophan in Mammals’ Health and Disease. Int. J. Mol. Sci. 2022, 23, 1222. [Google Scholar] [CrossRef]
- Vítek, L.; Haluzík, M. The Role of Bile Acids in Metabolic Regulation. J. Endocrinol. 2016, 228, R85–R96. [Google Scholar] [CrossRef]
- Li, B.; Xiong, Y.; Guo, D.; Deng, G.; Wu, H. The Gut-Reproductive Axis: Bridging Microbiota Balances to Reproductive Health and Fetal Development. Int. Immunopharmacol. 2025, 144, 113627. [Google Scholar] [CrossRef]
- Hamamah, S.; Barry, F.; Vannier, S.; Anahory, T.; Haahtela, T.; Antó, J.M.; Chapron, C.; Ayoubi, J.-M.; Czarlewski, W.; Bousquet, J. Infertility, IL-17, IL-33 and Microbiome Cross-Talk: The Extended ARIA-MeDALL Hypothesis. Int. J. Mol. Sci. 2024, 25, 11981. [Google Scholar] [CrossRef]
- Zhao, Z.; Ji, X.; Zhang, T.; Li, Q.; Marcella, C.; Wen, Q.; Cui, B.; Zhang, F. Washed Microbiota Transplantation Improves the Fertility of Patients with Inflammatory Bowel Disease. Chin. Med. J. 2022, 135, 1489–1491. [Google Scholar] [CrossRef]
- Lynch, S.V.; Pedersen, O. The Human Intestinal Microbiome in Health and Disease. N. Engl. J. Med. 2016, 375, 2369–2379. [Google Scholar] [CrossRef] [PubMed]
- Dominguez-Bello, M.G.; Costello, E.K.; Contreras, M.; Magris, M.; Hidalgo, G.; Fierer, N.; Knight, R. Delivery Mode Shapes the Acquisition and Structure of the Initial Microbiota across Multiple Body Habitats in Newborns. Proc. Natl. Acad. Sci. USA 2010, 107, 11971–11975. [Google Scholar] [CrossRef] [PubMed]
- Tamburini, S.; Shen, N.; Wu, H.C.; Clemente, J.C. The Microbiome in Early Life: Implications for Health Outcomes. Nat. Med. 2016, 22, 713–722. [Google Scholar] [CrossRef]
- Odamaki, T.; Kato, K.; Sugahara, H.; Hashikura, N.; Takahashi, S.; Xiao, J.-Z.; Abe, F.; Osawa, R. Age-Related Changes in Gut Microbiota Composition from Newborn to Centenarian: A Cross-Sectional Study. BMC Microbiol. 2016, 16, 90. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, T.S.; Shanahan, F.; O’Toole, P.W. The Gut Microbiome as a Modulator of Healthy Ageing. Nat. Rev. Gastroenterol. Hepatol. 2022, 19, 565–584. [Google Scholar] [CrossRef]
- Le Cosquer, G.; Vergnolle, N.; Motta, J.-P. Gut Microb-Aging and Its Relevance to Frailty Aging. Microbes Infect. 2024, 26, 105309. [Google Scholar] [CrossRef]
- Bosco, N.; Noti, M. The Aging Gut Microbiome and Its Impact on Host Immunity. Genes. Immun. 2021, 22, 289–303. [Google Scholar] [CrossRef]
- Wu, G.D.; Chen, J.; Hoffmann, C.; Bittinger, K.; Chen, Y.-Y.; Keilbaugh, S.A.; Bewtra, M.; Knights, D.; Walters, W.A.; Knight, R.; et al. Linking Long-Term Dietary Patterns with Gut Microbial Enterotypes. Science 2011, 334, 105–108. [Google Scholar] [CrossRef]
- Bradley, E.; Haran, J. The Human Gut Microbiome and Aging. Gut Microbes 2024, 16, 2359677. [Google Scholar] [CrossRef]
- David, L.A.; Maurice, C.F.; Carmody, R.N.; Gootenberg, D.B.; Button, J.E.; Wolfe, B.E.; Ling, A.V.; Devlin, A.S.; Varma, Y.; Fischbach, M.A.; et al. Diet Rapidly and Reproducibly Alters the Human Gut Microbiome. Nature 2014, 505, 559–563. [Google Scholar] [CrossRef]
- Bonder, M.J.; Kurilshikov, A.; Tigchelaar, E.F.; Mujagic, Z.; Imhann, F.; Vila, A.V.; Deelen, P.; Vatanen, T.; Schirmer, M.; Smeekens, S.P.; et al. The Effect of Host Genetics on the Gut Microbiome. Nat. Genet. 2016, 48, 1407–1412. [Google Scholar] [CrossRef] [PubMed]
- Karbownik, M.S.; Mokros, Ł.; Dobielska, M.; Kowalczyk, M.; Kowalczyk, E. Association Between Consumption of Fermented Food and Food-Derived Prebiotics With Cognitive Performance, Depressive, and Anxiety Symptoms in Psychiatrically Healthy Medical Students Under Psychological Stress: A Prospective Cohort Study. Front. Nutr. 2022, 9, 850249. [Google Scholar] [CrossRef] [PubMed]
- Clauss, M.; Gérard, P.; Mosca, A.; Leclerc, M. Interplay Between Exercise and Gut Microbiome in the Context of Human Health and Performance. Front. Nutr. 2021, 8, 637010. [Google Scholar] [CrossRef]
- Depommier, C.; Everard, A.; Druart, C.; Plovier, H.; Van Hul, M.; Vieira-Silva, S.; Falony, G.; Raes, J.; Maiter, D.; Delzenne, N.M.; et al. Supplementation with Akkermansia Muciniphila in Overweight and Obese Human Volunteers: A Proof-of-Concept Exploratory Study. Nat. Med. 2019, 25, 1096–1103. [Google Scholar] [CrossRef]
- Castellanos, N.; Diez, G.G.; Antúnez-Almagro, C.; Bailén, M.; Bressa, C.; González Soltero, R.; Pérez, M.; Larrosa, M. A Critical Mutualism—Competition Interplay Underlies the Loss of Microbial Diversity in Sedentary Lifestyle. Front. Microbiol. 2019, 10, 3142. [Google Scholar] [CrossRef]
- Xu, L.; Li, W.; Ling, L.; Zhang, Z.; Cui, Z.; Ge, J.; Wang, Y.; Meng, Q.; Wang, Y.; Liu, K.; et al. A Sedentary Lifestyle Changes the Composition and Predicted Functions of the Gut Bacterial and Fungal Microbiota of Subjects from the Same Company. Curr. Microbiol. 2023, 80, 368. [Google Scholar] [CrossRef]
- Karl, J.P.; Hatch, A.M.; Arcidiacono, S.M.; Pearce, S.C.; Pantoja-Feliciano, I.G.; Doherty, L.A.; Soares, J.W. Effects of Psychological, Environmental and Physical Stressors on the Gut Microbiota. Front. Microbiol. 2018, 9, 2013. [Google Scholar] [CrossRef]
- Camilleri, M. The Leaky Gut: Mechanisms, Measurement and Clinical Implications in Humans. Gut 2019, 68, 1516–1526. [Google Scholar] [CrossRef]
- Doenyas, C.; Clarke, G.; Cserjési, R. Gut–Brain Axis and Neuropsychiatric Health: Recent Advances. Sci. Rep. 2025, 15, 3415. [Google Scholar] [CrossRef]
- Ashonibare, V.J.; Akorede, B.A.; Ashonibare, P.J.; Akhigbe, T.M.; Akhigbe, R.E. Gut Microbiota-Gonadal Axis: The Impact of Gut Microbiota on Reproductive Functions. Front. Immunol. 2024, 15, 1346035. [Google Scholar] [CrossRef]
- Kurilshikov, A.; Medina-Gomez, C.; Bacigalupe, R.; Radjabzadeh, D.; Wang, J.; Demirkan, A.; Le Roy, C.I.; Raygoza Garay, J.A.; Finnicum, C.T.; Liu, X.; et al. Large-Scale Association Analyses Identify Host Factors Influencing Human Gut Microbiome Composition. Nat. Genet. 2021, 53, 156–165. [Google Scholar] [CrossRef] [PubMed]
- Manor, O.; Dai, C.L.; Kornilov, S.A.; Smith, B.; Price, N.D.; Lovejoy, J.C.; Gibbons, S.M.; Magis, A.T. Health and Disease Markers Correlate with Gut Microbiome Composition across Thousands of People. Nat. Commun. 2020, 11, 5206. [Google Scholar] [CrossRef]
- Hu, S.; Ding, Q.; Zhang, W.; Kang, M.; Ma, J.; Zhao, L. Gut Microbial Beta-Glucuronidase: A Vital Regulator in Female Estrogen Metabolism. Gut Microbes 2023, 15, 2236749. [Google Scholar] [CrossRef] [PubMed]
- Schiffer, L.; Barnard, L.; Baranowski, E.S.; Gilligan, L.C.; Taylor, A.E.; Arlt, W.; Shackleton, C.H.L.; Storbeck, K.-H. Human Steroid Biosynthesis, Metabolism and Excretion Are Differentially Reflected by Serum and Urine Steroid Metabolomes: A Comprehensive Review. J. Steroid Biochem. Mol. Biol. 2019, 194, 105439. [Google Scholar] [CrossRef]
- Fernández-Murga, M.L.; Gil-Ortiz, F.; Serrano-García, L.; Llombart-Cussac, A. A New Paradigm in the Relationship between Gut Microbiota and Breast Cancer: β-Glucuronidase Enzyme Identified as Potential Therapeutic Target. Pathogens 2023, 12, 1086. [Google Scholar] [CrossRef]
- Bulun, S.E.; Yang, S.; Fang, Z.; Gurates, B.; Tamura, M.; Sebastian, S. Estrogen Production and Metabolism in Endometriosis. Ann. N. Y. Acad. Sci. 2002, 955, 75–85. [Google Scholar] [CrossRef]
- Scarfò, G.; Daniele, S.; Chelucci, E.; Papini, F.; Epifani, F.; Ruggiero, M.; Cela, V.; Franzoni, F.; Artini, P.G. Endometrial Dysbiosis: A Possible Association with Estrobolome Alteration. Biomolecules 2024, 14, 1325. [Google Scholar] [CrossRef]
- O’Riordan, K.J.; Collins, M.K.; Moloney, G.M.; Knox, E.G.; Aburto, M.R.; Fülling, C.; Morley, S.J.; Clarke, G.; Schellekens, H.; Cryan, J.F. Short Chain Fatty Acids: Microbial Metabolites for Gut-Brain Axis Signalling. Mol. Cell. Endocrinol. 2022, 546, 111572. [Google Scholar] [CrossRef]
- Forte, N.; Marfella, B.; Nicois, A.; Palomba, L.; Paris, D.; Motta, A.; Pina Mollica, M.; Di Marzo, V.; Cristino, L. The Short-Chain Fatty Acid Acetate Modulates Orexin/Hypocretin Neurons: A Novel Mechanism in Gut-Brain Axis Regulation of Energy Homeostasis and Feeding. Biochem. Pharmacol. 2024, 226, 116383. [Google Scholar] [CrossRef]
- Ashraf, A.; Hassan, M.I. Microbial Endocrinology: Host Metabolism and Appetite Hormones Interaction with Gut Microbiome. Mol. Cell. Endocrinol. 2024, 592, 112281. [Google Scholar] [CrossRef]
- Wang, Z.; Zhang, L.; Liu, X.; Xu, L. The Role of Reproductive Tract Microbiota in Gynecological Health and Diseases. J. Reprod. Immunol. 2025, 167, 104418. [Google Scholar] [CrossRef] [PubMed]
- Garmendia, J.V.; De Sanctis, C.V.; Hajdúch, M.; De Sanctis, J.B. Microbiota and Recurrent Pregnancy Loss (RPL); More than a Simple Connection. Microorganisms 2024, 12, 1641. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Chen, H.; Feng, L.; Zhang, J. Interactions between Gut Microbiota and Metabolites Modulate Cytokine Network Imbalances in Women with Unexplained Miscarriage. Npj Biofilms Microbiomes 2021, 7, 1–12. [Google Scholar] [CrossRef]
- Vomstein, K.; Krog, M.C.; Wrønding, T.; Nielsen, H.S. The Microbiome in Recurrent Pregnancy Loss—A Scoping Review. J. Reprod. Immunol. 2024, 163, 104251. [Google Scholar] [CrossRef]
- Li, Z.; Zheng, Y.; Zhang, M.; Wu, K.; Zhang, L.; Yao, Y.; Zheng, C. Gut Microbiota-Derived Metabolites Associate with Circulating Immune Cell Subsets in Unexplained Recurrent Spontaneous Abortion. Heliyon 2024, 10, e24571. [Google Scholar] [CrossRef]
- Hang, S.; Paik, D.; Yao, L.; Kim, E.; Trinath, J.; Lu, J.; Ha, S.; Nelson, B.N.; Kelly, S.P.; Wu, L.; et al. Bile Acid Metabolites Control TH17 and Treg Cell Differentiation. Nature 2019, 576, 143–148. [Google Scholar] [CrossRef]
- Beni, F.A.; Saffarfar, H.; Elhami, A.; Kazemi, M. Gut Microbiota Dysbiosis: A Neglected Risk Factor for Male and Female Fertility. Cell. Microbiol. 2024, 2024, 7808354. [Google Scholar] [CrossRef]
- Qi, X.; Yun, C.; Pang, Y.; Qiao, J. The Impact of the Gut Microbiota on the Reproductive and Metabolic Endocrine System. Gut Microbes 2021, 13, 1894070. [Google Scholar] [CrossRef]
- Wei, Y.; Tan, H.; Yang, R.; Yang, F.; Liu, D.; Huang, B.; OuYang, L.; Lei, S.; Wang, Z.; Jiang, S.; et al. Gut Dysbiosis-Derived β-Glucuronidase Promotes the Development of Endometriosis. Fertil. Steril. 2023, 120, 682–694. [Google Scholar] [CrossRef]
- Pai, A.H.-Y.; Wang, Y.-W.; Lu, P.-C.; Wu, H.-M.; Xu, J.-L.; Huang, H.-Y. Gut Microbiome–Estrobolome Profile in Reproductive-Age Women with Endometriosis. Int. J. Mol. Sci. 2023, 24, 16301. [Google Scholar] [CrossRef]
- Ramírez-Pavez, T.N.; Martínez-Esparza, M.; Ruiz-Alcaraz, A.J.; Marín-Sánchez, P.; Machado-Linde, F.; García-Peñarrubia, P. The Role of Peritoneal Macrophages in Endometriosis. Int. J. Mol. Sci. 2021, 22, 10792. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Liu, Y.; Zhong, Z.; Wei, C.; Liu, Y.; Zhu, X. Peritoneal Immune Microenvironment of Endometriosis: Role and Therapeutic Perspectives. Front. Immunol. 2023, 14, 1134663. [Google Scholar] [CrossRef] [PubMed]
- Iba, Y.; Harada, T.; Horie, S.; Deura, I.; Iwabe, T.; Terakawa, N. Lipopolysaccharide-Promoted Proliferation of Endometriotic Stromal Cells via Induction of Tumor Necrosis Factor α and Interleukin-8 Expression. Fertil. Steril. 2004, 82, 1036–1042. [Google Scholar] [CrossRef] [PubMed]
- Harada, T.; Yoshioka, H.; Yoshida, S.; Iwabe, T.; Onohara, Y.; Tanikawa, M.; Terakawa, N. Increased Interleukin-6 Levels in Peritoneal Fluid of Infertile Patients with Active Endometriosis. Am. J. Obstet. Gynecol. 1997, 176, 593–597. [Google Scholar] [CrossRef]
- Iwabe, T.; Harada, T.; Tsudo, T.; Tanikawa, M.; Onohara, Y.; Terakawa, N. Pathogenetic Significance of Increased Levels of Interleukin-8 in the Peritoneal Fluid of Patients with Endometriosis. Fertil. Steril. 1998, 69, 924–930. [Google Scholar] [CrossRef]
- Surrey, E.S.; Halme, J. Effect of Peritoneal Fluid from Endometriosis Patients on Endometrial Stromal Cell Proliferation in Vitro. Obstet. Gynecol. 1990, 76, 792–797. [Google Scholar] [CrossRef]
- Chadchan, S.B.; Popli, P.; Ambati, C.R.; Tycksen, E.; Han, S.J.; Bulun, S.E.; Putluri, N.; Biest, S.W.; Kommagani, R. Gut Microbiota–Derived Short-Chain Fatty Acids Protect against the Progression of Endometriosis. Life Sci. Alliance 2021, 4, e202101224. [Google Scholar] [CrossRef]
- Guo, J.; Wang, Y.; Jiang, P.; Yao, H.; Zhao, C.; Hu, X.; Cao, Y.; Zhang, N.; Fu, Y.; Shen, H. Sodium Butyrate Alleviates Lipopolysaccharide-Induced Endometritis in Mice through Inhibiting Inflammatory Response. Microb. Pathog. 2019, 137, 103792. [Google Scholar] [CrossRef]
- Cui, H.; Wang, N.; Li, H.; Bian, Y.; Wen, W.; Kong, X.; Wang, F. The Dynamic Shifts of IL-10-Producing Th17 and IL-17-Producing Treg in Health and Disease: A Crosstalk between Ancient “Yin-Yang” Theory and Modern Immunology. Cell Commun. Signal. 2024, 22, 99. [Google Scholar] [CrossRef]
- Tanaka, Y.; Mori, T.; Ito, F.; Koshiba, A.; Takaoka, O.; Kataoka, H.; Maeda, E.; Okimura, H.; Mori, T.; Kitawaki, J. Exacerbation of Endometriosis Due To Regulatory T-Cell Dysfunction. J. Clin. Endocrinol. Metab. 2017, 102, 3206–3217. [Google Scholar] [CrossRef]
- Chadchan, S.B.; Naik, S.K.; Popli, P.; Talwar, C.; Putluri, S.; Ambati, C.R.; Lint, M.A.; Kau, A.L.; Stallings, C.L.; Kommagani, R. Gut Microbiota and Microbiota-Derived Metabolites Promotes Endometriosis. Cell Death Discov. 2023, 9, 28. [Google Scholar] [CrossRef] [PubMed]
- Mor, A.; Tankiewicz-Kwedlo, A.; Ciwun, M.; Lewkowicz, J.; Pawlak, D. Kynurenines as a Novel Target for the Treatment of Inflammatory Disorders. Cells 2024, 13, 1259. [Google Scholar] [CrossRef] [PubMed]
- Bergqvist, A.; Bruse, C.; Carlberg, M.; Carlström, K. Interleukin 1β, Interleukin-6, and Tumor Necrosis Factor-α in Endometriotic Tissue and in Endometrium. Fertil. Steril. 2001, 75, 489–495. [Google Scholar] [CrossRef]
- Talwar, C.; Davuluri, G.V.N.; Kamal, A.H.M.; Coarfa, C.; Han, S.J.; Veeraragavan, S.; Parsawar, K.; Putluri, N.; Hoffman, K.; Jimenez, P.; et al. Identification of Distinct Stool Metabolites in Women with Endometriosis for Non-Invasive Diagnosis and Potential for Microbiota-Based Therapies. Med 2025, 6, 100517. [Google Scholar] [CrossRef]
- Lu, J.; Ling, X.; Liu, L.; Jiang, A.; Ren, C.; Lu, C.; Yu, Z. Emerging Hallmarks of Endometriosis Metabolism: A Promising Target for the Treatment of Endometriosis. Biochim. Biophys. Acta (BBA)—Mol. Cell Res. 2023, 1870, 119381. [Google Scholar] [CrossRef]
- Muraoka, A.; Suzuki, M.; Hamaguchi, T.; Watanabe, S.; Iijima, K.; Murofushi, Y.; Shinjo, K.; Osuka, S.; Hariyama, Y.; Ito, M.; et al. Fusobacterium Infection Facilitates the Development of Endometriosis through the Phenotypic Transition of Endometrial Fibroblasts. Sci. Transl. Med. 2023, 15, eadd1531. [Google Scholar] [CrossRef]
- Hicks, C.; Leonardi, M.; Chua, X.-Y.; Mari-Breedt, L.; Espada, M.; El-Omar, E.M.; Condous, G.; El-Assaad, F. Oral, Vaginal, and Stool Microbial Signatures in Patients With Endometriosis as Potential Diagnostic Non-Invasive Biomarkers: A Prospective Cohort Study. BJOG Int. J. Obstet. Gynaecol. 2025, 132, 326–336. [Google Scholar] [CrossRef]
- Torres, P.J.; Siakowska, M.; Banaszewska, B.; Pawelczyk, L.; Duleba, A.J.; Kelley, S.T.; Thackray, V.G. Gut Microbial Diversity in Women With Polycystic Ovary Syndrome Correlates With Hyperandrogenism. J. Clin. Endocrinol. Metab. 2018, 103, 1502–1511. [Google Scholar] [CrossRef]
- Olaniyi, K.S.; Bashir, A.M.; Areloegbe, S.E.; Sabinari, I.W.; Akintayo, C.O.; Oniyide, A.A.; Aturamu, A. Short Chain Fatty Acid, Acetate Restores Ovarian Function in Experimentally Induced PCOS Rat Model. PLoS ONE 2022, 17, e0272124. [Google Scholar] [CrossRef]
- Patel, J.; Chaudhary, H.; Rajput, K.; Parekh, B.; Joshi, R. Assessment of Gut Microbial β-Glucuronidase and β-Glucosidase Activity in Women with Polycystic Ovary Syndrome. Sci. Rep. 2023, 13, 11967. [Google Scholar] [CrossRef]
- Huang, F.; Deng, Y.; Zhou, M.; Tang, R.; Zhang, P.; Chen, R. Fecal Microbiota Transplantation from Patients with Polycystic Ovary Syndrome Induces Metabolic Disorders and Ovarian Dysfunction in Germ-Free Mice. BMC Microbiol. 2024, 24, 364. [Google Scholar] [CrossRef] [PubMed]
- Martinelli, S.; Nannini, G.; Cianchi, F.; Staderini, F.; Coratti, F.; Amedei, A. Microbiota Transplant and Gynecological Disorders: The Bridge between Present and Future Treatments. Microorganisms 2023, 11, 2407. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Gao, W.; Li, D. Recurrent Implantation Failure: A Comprehensive Summary from Etiology to Treatment. Front. Endocrinol. 2023, 13, 1061766. [Google Scholar] [CrossRef]
- Patel, N.; Patel, N.; Pal, S.; Nathani, N.; Pandit, R.; Patel, M.; Patel, N.; Joshi, C.; Parekh, B. Distinct Gut and Vaginal Microbiota Profile in Women with Recurrent Implantation Failure and Unexplained Infertility. BMC Womens Health 2022, 22, 113. [Google Scholar] [CrossRef]
- Lee, S.K.; Kim, J.Y.; Hur, S.E.; Kim, C.J.; Na, B.J.; Lee, M.; Gilman-Sachs, A.; Kwak-Kim, J. An Imbalance in Interleukin-17-Producing T and Foxp3+ Regulatory T Cells in Women with Idiopathic Recurrent Pregnancy Loss. Hum. Reprod. 2011, 26, 2964–2971. [Google Scholar] [CrossRef]
- Balla, B.; Illés, A.; Tobiás, B.; Pikó, H.; Beke, A.; Sipos, M.; Lakatos, P.; Kósa, J.P. The Role of the Vaginal and Endometrial Microbiomes in Infertility and Their Impact on Pregnancy Outcomes in Light of Recent Literature. Int. J. Mol. Sci. 2024, 25, 13227. [Google Scholar] [CrossRef]
- Bhutta, N.K.; Xu, X.; Jian, C.; Wang, Y.; Liu, Y.; Sun, J.; Han, B.; Wu, S.; Javeed, A. Gut Microbiota Mediated T Cells Regulation and Autoimmune Diseases. Front. Microbiol. 2024, 15, 1477187. [Google Scholar] [CrossRef]
- D’Ippolito, S.; Gasbarrini, A.; Castellani, R.; Rocchetti, S.; Sisti, L.G.; Scambia, G.; Di Simone, N. Human Leukocyte Antigen (HLA) DQ2/DQ8 Prevalence in Recurrent Pregnancy Loss Women. Autoimmun. Rev. 2016, 15, 638–643. [Google Scholar] [CrossRef]
- Yoshihara, H.; Goto, S.; Kitaori, T.; Sugiura-Ogasawara, M. Association between Antinuclear Antibodies and Pregnancy Prognosis in Recurrent Pregnancy Loss Patients. Hum. Reprod. 2025, 40, 236–243. [Google Scholar] [CrossRef]
- Chioma, O.S.; Mallott, E.; Shah-Gandhi, B.; Wiggins, Z.; Langford, M.; Lancaster, A.W.; Gelbard, A.; Wu, H.; Johnson, J.E.; Lancaster, L.; et al. Low Gut Microbial Diversity Augments Estrogen-Driven Pulmonary Fibrosis in Female-Predominant Interstitial Lung Disease. Cells 2023, 12, 766. [Google Scholar] [CrossRef]
- Inoue, T.; Kanzaki, H.; Iwai, M.; Imai, K.; Narukawa, S.; Higuchi, T.; Katsuragawa, H.; Mori, T. Tumour Necrosis Factor Alpha Inhibits In-Vitro Decidualization of Human Endometrial Stromal Cells. Hum. Reprod. 1994, 9, 2411–2417. [Google Scholar] [CrossRef] [PubMed]
- Larsen, J.M. The Immune Response to Prevotella Bacteria in Chronic Inflammatory Disease. Immunology 2017, 151, 363–374. [Google Scholar] [CrossRef] [PubMed]
- Farshchi, M.; Abdollahi, E.; Saghafi, N.; Hosseini, A.; Fallahi, S.; Rostami, S.; Rostami, P.; Rafatpanah, H.; Habibagahi, M. Evaluation of Th17 and Treg Cytokines in Patients with Unexplained Recurrent Pregnancy Loss. J. Clin. Transl. Res. 2022, 8, 256–265. [Google Scholar] [PubMed]
- de la Fuente-Munoz, E.; Fernández-Arquero, M.; Subbhi-Issa, N.; Guevara-Hoyer, K.; Suárez, L.P.; Laborda, R.G.; Sánchez, M.; Ochoa-Grullón, J.; Guzmán-Fulgencio, M.; Villegas, Á.; et al. Recurrent Reproductive Failure and Celiac Genetic Susceptibility, a Leading Role of Gluten. Front. Immunol. 2024, 15, 1451552. [Google Scholar] [CrossRef]
- Jarne-Borràs, M.; Miró-Mur, F.; Anunciación-Llunell, A.; Alijotas-Reig, J. Antiphospholipid Antibodies in Women with Recurrent Embryo Implantation Failure: A Systematic Review and Meta-Analysis. Autoimmun. Rev. 2022, 21, 103101. [Google Scholar] [CrossRef]
- Bayar, E.; Bennett, P.R.; Chan, D.; Sykes, L.; MacIntyre, D.A. The Pregnancy Microbiome and Preterm Birth. Semin. Immunopathol. 2020, 42, 487–499. [Google Scholar] [CrossRef]
- Dahl, C.; Stanislawski, M.; Iszatt, N.; Mandal, S.; Lozupone, C.; Clemente, J.C.; Knight, R.; Stigum, H.; Eggesbø, M. Gut Microbiome of Mothers Delivering Prematurely Shows Reduced Diversity and Lower Relative Abundance of Bifidobacterium and Streptococcus. PLoS ONE 2017, 12, e0184336. [Google Scholar] [CrossRef]
- Shiozaki, A.; Yoneda, S.; Yoneda, N.; Yonezawa, R.; Matsubayashi, T.; Seo, G.; Saito, S. Intestinal Microbiota Is Different in Women with Preterm Birth: Results from Terminal Restriction Fragment Length Polymorphism Analysis. PLoS ONE 2014, 9, e111374. [Google Scholar] [CrossRef]
- Gomez-Lopez, N.; StLouis, D.; Lehr, M.A.; Sanchez-Rodriguez, E.N.; Arenas-Hernandez, M. Immune Cells in Term and Preterm Labor. Cell Mol. Immunol. 2014, 11, 571–581. [Google Scholar] [CrossRef]
- Karrar, S.A.; Martingano, D.J.; Hong, P.L. Preeclampsia. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2025. [Google Scholar]
- Deady, C.; McCarthy, F.P.; Barron, A.; McCarthy, C.M.; O’Keeffe, G.W.; O’Mahony, S.M. An Altered Gut Microbiome in Pre-Eclampsia: Cause or Consequence. Front. Cell. Infect. Microbiol. 2024, 14, 1352267. [Google Scholar] [CrossRef]
- Zhao, H.J.; Chen, Y.; Liu, T.; McArthur, K.; Mueller, N.T. Short-Chain Fatty Acids and Preeclampsia: A Scoping Review. Nutr. Rev. 2025, 83, e683–e693. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Li, J.; Li, N.; Liu, H.; Tang, J. Increased Circulating Trimethylamine N-Oxide Plays a Contributory Role in the Development of Endothelial Dysfunction and Hypertension in the RUPP Rat Model of Preeclampsia. Hypertens. Pregnancy 2019, 38, 96–104. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Gu, X.; Yang, J.; Wei, Y.; Zhao, Y. Gut Microbiota Dysbiosis and Increased Plasma LPS and TMAO Levels in Patients With Preeclampsia. Front. Cell Infect. Microbiol. 2019, 9, 409. [Google Scholar] [CrossRef] [PubMed]
- Wilson Tang, W.H.; Wang, Z.; Kennedy, D.J.; Wu, Y.; Buffa, J.A.; Agatisa-Boyle, B.; Li, X.S.; Levison, B.S.; Hazen, S.L. Gut Microbiota-Dependent Trimethylamine N-Oxide (TMAO) Pathway Contributes to Both Development of Renal Insufficiency and Mortality Risk in Chronic Kidney Disease. Circ. Res. 2015, 116, 448–455. [Google Scholar] [CrossRef]
- Wen, Y.; Peng, L.; Xu, R.; Zang, N.; Huang, Q.; Zhong, M. Maternal Serum Trimethylamine-N-Oxide Is Significantly Increased in Cases with Established Preeclampsia. Pregnancy Hypertens. 2019, 15, 114–117. [Google Scholar] [CrossRef]
- Xu, H.; Feng, P.; Sun, Y.; Wu, D.; Wang, D.; Wu, L.; Peng, H.; Li, H. Plasma Trimethylamine N-Oxide Metabolites in the Second Trimester Predict the Risk of Hypertensive Disorders of Pregnancy: A Nested Case-Control Study. Hypertens. Res. 2024, 47, 778–789. [Google Scholar] [CrossRef]
- Chang, Y.; Chen, Y.; Zhou, Q.; Wang, C.; Chen, L.; Di, W.; Zhang, Y. Short-Chain Fatty Acids Accompanying Changes in the Gut Microbiome Contribute to the Development of Hypertension in Patients with Preeclampsia. Clin. Sci. 2020, 134, 289–302. [Google Scholar] [CrossRef]
- Mo, C.; Lou, X.; Xue, J.; Shi, Z.; Zhao, Y.; Wang, F.; Chen, G. The Influence of Akkermansia Muciniphila on Intestinal Barrier Function. Gut Pathog. 2024, 16, 41. [Google Scholar] [CrossRef]
- Romão-Veiga, M.; Bannwart-Castro, C.F.; Borges, V.T.M.; Golim, M.A.; Peraçoli, J.C.; Peraçoli, M.T.S. Increased TLR4 Pathway Activation and Cytokine Imbalance Led to Lipopolysaccharide Tolerance in Monocytes from Preeclamptic Women. Pregnancy Hypertens. 2020, 21, 159–165. [Google Scholar] [CrossRef]
- Karoor, V.; Strassheim, D.; Sullivan, T.; Verin, A.; Umapathy, N.S.; Dempsey, E.C.; Frank, D.N.; Stenmark, K.R.; Gerasimovskaya, E. The Short-Chain Fatty Acid Butyrate Attenuates Pulmonary Vascular Remodeling and Inflammation in Hypoxia-Induced Pulmonary Hypertension. Int. J. Mol. Sci. 2021, 22, 9916. [Google Scholar] [CrossRef]
- Wan, Y.-J.Y.; Sheng, L. Regulation of Bile Acid Receptor Activity. Liver Res. 2018, 2, 180–185. [Google Scholar] [CrossRef] [PubMed]
- Ching, T.; Ha, J.; Song, M.-A.; Tiirikainen, M.; Molnar, J.; Berry, M.J.; Towner, D.; Garmire, L.X. Genome-Scale Hypomethylation in the Cord Blood DNAs Associated with Early Onset Preeclampsia. Clin. Epigenetics 2015, 7, 21. [Google Scholar] [CrossRef] [PubMed]
- Korczynska, L.; Zeber-Lubecka, N.; Zgliczynska, M.; Zarychta, E.; Zareba, K.; Wojtyla, C.; Dabrowska, M.; Ciebiera, M. The Role of Microbiota in the Pathophysiology of Uterine Fibroids—A Systematic Review. Front. Cell Infect. Microbiol. 2023, 13, 1177366. [Google Scholar] [CrossRef] [PubMed]
- Kong, Y.; Liu, S.; Wang, X.; Qie, R. Associations between Gut Microbiota and Gynecological Cancers: A Bi-Directional Two-Sample Mendelian Randomization Study. Medicine 2024, 103, e37628. [Google Scholar] [CrossRef]
- Xavier, J.B.; Young, V.B.; Skufca, J.; Ginty, F.; Testerman, T.; Pearson, A.T.; Macklin, P.; Mitchell, A.; Shmulevich, I.; Xie, L.; et al. The Cancer Microbiome: Distinguishing Direct and Indirect Effects Requires a Systemic View. Trends Cancer 2020, 6, 192–204. [Google Scholar] [CrossRef]
- Schilling, J.; Loening-Baucke, V.; Dörffel, Y. Increased Gardnerella Vaginalis Urogenital Biofilm in Inflammatory Bowel Disease. J. Crohns Colitis 2014, 8, 543–549. [Google Scholar] [CrossRef]
- Amabebe, E.; Anumba, D.O.C. Female Gut and Genital Tract Microbiota-Induced Crosstalk and Differential Effects of Short-Chain Fatty Acids on Immune Sequelae. Front. Immunol. 2020, 11, 2184. [Google Scholar] [CrossRef]
- Li, M.; Zhang, Q.; Wu, T.; Ma, L.; Hu, D.; Yuan, Z.; Wang, S.; Luo, A.; Zhang, J. How Does Gut Microbiota Affect the Vaginitis Axis? The Mediating Role of Plasma Metabolites. Microbiol. Spectr. 2025, 13, e02263-24. [Google Scholar] [CrossRef]
- Delgado-Diaz, D.J.; Tyssen, D.; Hayward, J.A.; Gugasyan, R.; Hearps, A.C.; Tachedjian, G. Distinct Immune Responses Elicited From Cervicovaginal Epithelial Cells by Lactic Acid and Short Chain Fatty Acids Associated With Optimal and Non-Optimal Vaginal Microbiota. Front. Cell. Infect. Microbiol. 2020, 9, 446. [Google Scholar] [CrossRef]
- Wei, T.; Geiser, A.G.; Qian, H.-R.; Su, C.; Helvering, L.M.; Kulkarini, N.H.; Shou, J.; N’Cho, M.; Bryant, H.U.; Onyia, J.E. DNA Microarray Data Integration by Ortholog Gene Analysis Reveals Potential Molecular Mechanisms of Estrogen-Dependent Growth of Human Uterine Fibroids. BMC Womens Health 2007, 7, 5. [Google Scholar] [CrossRef]
- Peng, S.; Liu, M.; Zeng, Y.; Wang, L.; Man, Y. Exploring the Gut-Inflammation Connection: A Mendelian Randomization Study on Gut Microbiota, Inflammatory Factors, and Uterine Fibroids Risk. Medicine 2024, 103, e40514. [Google Scholar] [CrossRef] [PubMed]
- Protic, O.; Toti, P.; Islam, M.S.; Occhini, R.; Giannubilo, S.R.; Catherino, W.H.; Cinti, S.; Petraglia, F.; Ciavattini, A.; Castellucci, M.; et al. Possible Involvement of Inflammatory/Reparative Processes in the Development of Uterine Fibroids. Cell Tissue Res. 2016, 364, 415–427. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Mo, J.; Huang, W.; Bao, Y.; Luo, X.; Yuan, L. The Ovarian Cancer-Associated Microbiome Contributes to the Tumor’s Inflammatory Microenvironment. Front. Cell Infect. Microbiol. 2024, 14, 1440742. [Google Scholar] [CrossRef]
- Liu, Y.; Cao, L. Intratumoral Lactobacillus Iners as a Poor Prognostic Biomarker and Potential Therapeutic Target for Cervical Cancer. Front. Cell. Infect. Microbiol. 2024, 14, 1469924. [Google Scholar] [CrossRef]
- Jimenez, N.; Norton, T.; Diadala, G.; Bell, E.; Valenti, M.; Farland, L.V.; Mahnert, N.; Herbst-Kralovetz, M.M. Vaginal and Rectal Microbiome Contribute to Genital Inflammation in Chronic Pelvic Pain. BMC Med. 2024, 22, 283. [Google Scholar] [CrossRef]
- Yao, Y.; Hu, H.; Chen, L.; Zheng, H. Association between Gut Microbiota and Menstrual Disorders: A Two-Sample Mendelian Randomization Study. Front. Microbiol. 2024, 15, 1321268. [Google Scholar] [CrossRef]
- Dekel, N.; Gnainsky, Y.; Granot, I.; Racicot, K.; Mor, G. The Role of Inflammation for a Successful Implantation. Am. J. Reprod. Immunol. 2014, 72, 141–147. [Google Scholar] [CrossRef]
- Wang, J.; Huang, C.; Jiang, R.; Du, Y.; Zhou, J.; Jiang, Y.; Yan, Q.; Xing, J.; Hou, X.; Zhou, J.; et al. Decreased Endometrial IL-10 Impairs Endometrial Receptivity by Downregulating HOXA10 Expression in Women with Adenomyosis. BioMed Res. Int. 2018, 2018, 2549789. [Google Scholar] [CrossRef]
- Pathare, A.D.S.; Zaveri, K.; Hinduja, I. Downregulation of Genes Related to Immune and Inflammatory Response in IVF Implantation Failure Cases under Controlled Ovarian Stimulation. Am. J. Reprod. Immunol. 2017, 78, e12679. [Google Scholar] [CrossRef]
- Al-Lamee, H.; Ellison, A.; Drury, J.; Hill, C.J.; Drakeley, A.J.; Hapangama, D.K.; Tempest, N. Altered Endometrial Oestrogen-Responsiveness and Recurrent Reproductive Failure. Reprod. Fertil. 2022, 3, 30–38. [Google Scholar] [CrossRef]
- Potiris, A.; Alyfanti, E.; Drakaki, E.; Mavrogianni, D.; Karampitsakos, T.; Machairoudias, P.; Topis, S.; Zikopoulos, A.; Skentou, C.; Panagopoulos, P.; et al. The Contribution of Proteomics in Understanding Endometrial Protein Expression in Women with Recurrent Implantation Failure. J. Clin. Med. 2024, 13, 2145. [Google Scholar] [CrossRef] [PubMed]
- Grund, S.; Grümmer, R. Direct Cell–Cell Interactions in the Endometrium and in Endometrial Pathophysiology. Int. J. Mol. Sci. 2018, 19, 2227. [Google Scholar] [CrossRef]
- Burnham, P.; Chen, F.; Cheng, A.P.; Srivatana, V.; Zhang, L.T.; Edusei, E.; Albakry, S.; Botticelli, B.; Guo, X.; Renaghan, A.; et al. Peritoneal Effluent Cell-Free DNA Sequencing in Peritoneal Dialysis Patients With and Without Peritonitis. Kidney Med. 2022, 4, 100383. [Google Scholar] [CrossRef]
- Di Vincenzo, F.; Del Gaudio, A.; Petito, V.; Lopetuso, L.R.; Scaldaferri, F. Gut Microbiota, Intestinal Permeability, and Systemic Inflammation: A Narrative Review. Intern. Emerg. Med. 2024, 19, 275–293. [Google Scholar] [CrossRef]
- Takada, K.; Melnikov, V.G.; Kobayashi, R.; Komine-Aizawa, S.; Tsuji, N.M.; Hayakawa, S. Female Reproductive Tract-Organ Axes. Front. Immunol. 2023, 14, 1110001. [Google Scholar] [CrossRef]
- Łaniewski, P.; Ilhan, Z.E.; Herbst-Kralovetz, M.M. The Microbiome and Gynaecological Cancer Development, Prevention and Therapy. Nat. Rev. Urol. 2020, 17, 232–250. [Google Scholar] [CrossRef] [PubMed]
- Rieder, F. The Gut Microbiome in Intestinal Fibrosis: Environmental Protector or Provocateur? Sci. Transl. Med. 2013, 5, 190ps10. [Google Scholar] [CrossRef]
- Chadchan, S.B.; Singh, V.; Kommagani, R. Female Reproductive Dysfunctions and the Gut Microbiota. J. Mol. Endocrinol. 2022, 69, R81–R94. [Google Scholar] [CrossRef]
- Wu, H.-J.; Wu, E. The Role of Gut Microbiota in Immune Homeostasis and Autoimmunity. Gut Microbes 2012, 3, 4–14. [Google Scholar] [CrossRef]
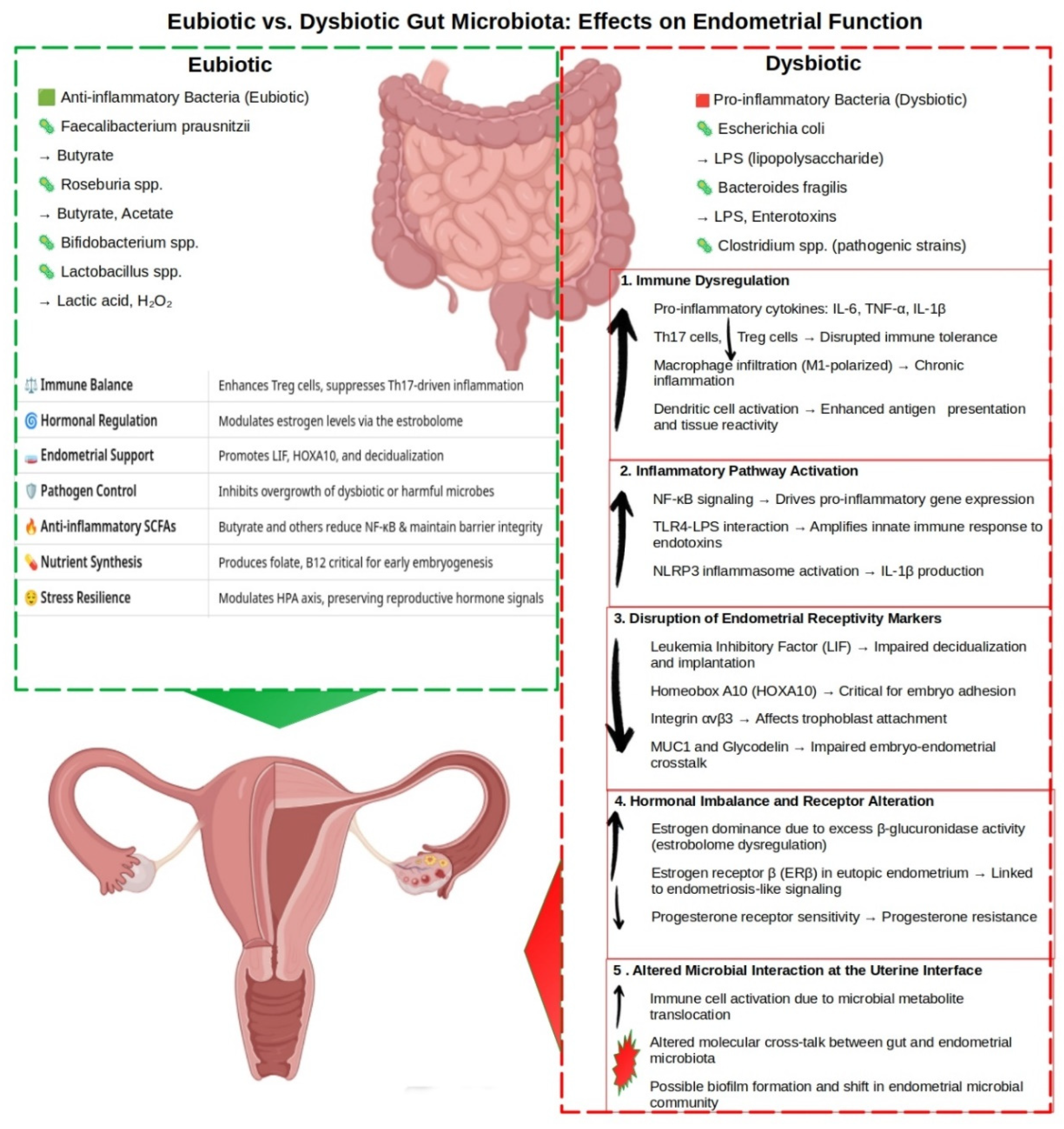

{kind=link}
| Hormone | Primary Source | Microbial Influence | Physiological Function | Therapeutic Potential |
|---|---|---|---|---|
| GLP-1 | Intestinal L-cells | SCFAs (butyrate and propionate) upregulate secretion via GPR41/GPR43 | Enhances insulin secretion; promotes satiety | Prebiotics and SCFA-promoting probiotics enhance GLP-1 for glycemic control and satiety |
| PYY | Intestinal L-cells | Stimulated by SCFAs; influenced by microbial density | Inhibits appetite; slows gastric emptying | Prebiotic fibers modulate PYY via SCFA production to reduce appetite and improve weight management |
| Ghrelin | Stomach X/A-like cells (in fundus) | Modulated by gut microbiota composition; lower in SCFA-rich profiles | Stimulates appetite; regulates energy balance | Microbial modulation (e.g., Akkermansia) may suppress ghrelin to reduce food intake and aid obesity management |
| Leptin | Adipocytes | Indirectly influenced via gut barrier integrity and systemic signals | Regulates satiety and energy expenditure | Gut barrier restoration via probiotics may enhance leptin sensitivity and reduce inflammation-associated resistance |
| Insulin | Pancreatic β-cells | Enhanced via SCFA-mediated GLP-1 secretion; impacted by bile acids | Regulates blood glucose levels | SCFA-driven incretin release and bile acid signaling can improve insulin sensitivity and glucose regulation |
| Neuropeptide Y (NPY) | Hypothalamic arcuate nucleus neurons. (AgRP/NPY neurons) | Suppressed by SCFA signaling and the microbial modulation of CNS | Promotes feeding behavior and energy storage | Diet and microbiota interventions targeting SCFA-NPY pathways may help control appetite and metabolic diseases |
| Orexin | Lateral hypothalamic area (LHA neurons) | Indirect modulation through microbiota–brain axis | Regulates arousal, wakefulness, and appetite | The probiotic modulation of the gut–brain axis may regulate orexin and improve sleep, mood, and appetite patterns |
| Regulatory Pathways | Microbial Drivers | Molecular Mediators | Target System | Action on Maternal Immune Interface | Key References |
|---|---|---|---|---|---|
| Immune Homeostasis | Faecalibacterium prausnitzii, Roseburia, Bifidobacterium spp. | SCFAs (butyrate, acetate), IL-10, TGF-β | CD4+ T cells (↑ Treg, ↓ Th17), NK cells | Promotes immune tolerance; suppresses pro-inflammatory cytokines | [42,43] |
| Inflammatory Regulation | Dysbiotic expansion: Prevotella, Erysipelotrichaceae, Enterococcus | LPS, IL-6, IL-1β, TNF-α, IL-17A | TLR4/NF-κB signaling in endometrium and periphery | Induces Th17-skewed inflammation; trophoblast apoptosis | [44,45] |
| Metabolite Signaling | ↓ Akkermansia, Anaerostipes, Ruminococcaceae | ↓ Bile acids (HDCA, LCA), ↑ Imidazolepropionic acid | FXR/GPBAR1, oxidative stress; epithelial integrity | Disrupted mucosal tolerance; elevated cytokines | [43,46] |
| Barrier Function | Lactobacillus spp., Akkermansia muciniphila | Mucin; tight junction proteins (ZO-1, claudins) | Intestinal and uterine epithelia | Reduced LPS translocation; protects from “leaky gut” | [42,47] |
| Endocrine Modulation | Clostridium scindens, Bacteroides spp. | Estrobolome (β-glucuronidase), steroid-modulating enzymes | Estrogen/progesterone bioavailability | Impacts endometrial receptivity; decidualization | [2,48] |
| Autoimmune Susceptibility | Dysbiotic networks in HLA-DQ2/DQ8+ hosts | Molecular mimicry, cross-reactive antigens | Autoantibodies, complement system | Increased maternal immune rejection of fetal cells | [42,44] |
| Mechanistic Domain | Microbial Features | Key Mediators | Immunological/Metabolic Consequences | References |
|---|---|---|---|---|
| 1. Microbial Diversity Collapse | ↓ Faecalibacterium, Bifidobacterium, Akkermansia, SCFA-producing Lactobacillus spp. ↑ Prevotella, Bacteroides, Enterococcus, Erysipelotrichaceae | LPS, peptidoglycan, flagellin TLR4 → NF-κB → IL-1β, IL-6, TNF-α | Increased gut permeability; systemic inflammation; impaired maternal–fetal tolerance | [42,43,44] |
| 2. Th17/Treg Imbalance | ↓ SCFA-producers (Roseburia, Anaerostipes, Blautia) ↑ Prevotella, Escherichia/Shigella | ↓ Tregs, ↑ Th17 IL-17A, IL-6, IL-8, IFN-γ | Impaired decidualization; abnormal NK cell recruitment; pro-inflammatory uterine environment | [43,75,76] |
| 3. Metabolite Dysregulation | ↓ Butyrate and bile acid-producing genera (Eubacterium, Clostridium XIVa) ↑ Histidine-metabolizing bacteria | ↓ SCFAs, ↓ HDCA and isoLCA ↑ Imidazolepropionic acid | Oxidative stress; barrier dysfunction; altered IL-10/IL-17 signaling | [43,45,52,77] |
| 4. Immunogenetic Susceptibility | HLA-DQ2/DQ8+genotype ↑ Firmicutes, Proteobacteria, Prevotella | Autoantibodies (ANA, aPL), complement activation | Cross-reactivity to fetal antigens; immune rejection; recurrent miscarriage | [42,44,78,79] |
| Microbial Taxa | Abundance in Pe | Associated Metabolites | Host Pathways Affected | Proposed Mechanism in Pe Pathophysiology | Key References |
|---|---|---|---|---|---|
| Escherichia/Shigella (Proteobacteria) | ↑ Increased | LPS (endotoxin) | TLR4 → NF-κB → IL-6, TNF-α | Promotes systemic inflammation; endothelial dysfunction | [42,98] |
| Blautia (Firmicutes) | ↓ Decreased | Butyrate and valerate | GPCR41/43 signaling; HDAC inhibition | Anti-inflammatory, vasodilatory; protects endothelial barrier | [98] |
| Eubacterium hallii | ↓ Decreased | Butyrate | SCFA receptor activation; mitochondrial support | Improves vascular tone; reduces oxidative stress | [98] |
| Bifidobacterium spp. | ↓ Decreased | Acetate and lactate | Enhances mucosal barrier integrity; immune modulation | Loss may increase gut permeability and LPS leakage | [43,98] |
| Subdoligranulum | ↓ Decreased | Butyrate | Treg induction; anti-inflammatory cytokines | Supports immune tolerance; depletion linked to Th17 shift | [98] |
| Enterobacter | ↑ Increased | LPS and TMA precursors | TLR4 activation; endothelial stress | Associated with hypertension and cytokine elevation | [98] |
| Akkermansia muciniphila | ↓ Decreased | Mucin-degradation products | Mucin layer maintenance; gut barrier protection | Depletion leads to “leaky gut” and metabolic inflammation | [42,99] |
| General SCFA Producers (Roseburia, Faecalibacterium) | ↓ Decreased | Butyrate, acetate, and propionate | GPR109A; Treg expansion | Critical for immune balance and endothelial protection | [43,46] |
| Condition | Microbial Features | Key Immune Mediators | Pathophysiological Impact | References |
|---|---|---|---|---|
| Bacterial Vaginosis (BV) | ↓ Lactobacillus, ↑ anaerobes (Gardnerella, etc.) | IL-6, IL-1β, TNF-α | Vaginal inflammation, biofilm formation, ↑ PID, miscarriage, preterm birth risk | [30] |
| Uterine Fibroids | ↑ β-glucuronidase (Clostridia) → ↑ estrogen | IL-6, M1 macrophages | Estrogen-driven fibroid growth, immune cell infiltration, fibrosis | [104] |
| Gynecologic Cancers | ↑ Fusobacterium, Bacteroides, E. coli | IL-6, TNF-α, ROS, ↓ NK cells | Chronic inflammation, immune evasion, mucosal breakdown, tumor promotion | [105,106] |
| Pelvic Inflammatory Disease (PID) | ↑ Gardnerella, Mycoplasma, Bacteroides | IL-1β, IL-8, TNF-α | Persistent pelvic inflammation, tubal scarring, infertility | [30] |
| Menstrual Irregularities | ↓ diversity, ↑ Bacteroides, Clostridium | IL-6, CRP, β-glucuronidase | Hormonal imbalance, anovulation, cycle disruption | [8] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Escorcia Mora, P.; Valbuena, D.; Diez-Juan, A. The Role of the Gut Microbiota in Female Reproductive and Gynecological Health: Insights into Endometrial Signaling Pathways. Life 2025, 15, 762. https://doi.org/10.3390/life15050762
Escorcia Mora P, Valbuena D, Diez-Juan A. The Role of the Gut Microbiota in Female Reproductive and Gynecological Health: Insights into Endometrial Signaling Pathways. Life. 2025; 15(5):762. https://doi.org/10.3390/life15050762
Chicago/Turabian StyleEscorcia Mora, Patricia, Diana Valbuena, and Antonio Diez-Juan. 2025. "The Role of the Gut Microbiota in Female Reproductive and Gynecological Health: Insights into Endometrial Signaling Pathways" Life 15, no. 5: 762. https://doi.org/10.3390/life15050762
APA StyleEscorcia Mora, P., Valbuena, D., & Diez-Juan, A. (2025). The Role of the Gut Microbiota in Female Reproductive and Gynecological Health: Insights into Endometrial Signaling Pathways. Life, 15(5), 762. https://doi.org/10.3390/life15050762