
Metagenomic Characterization of Microbiome Taxa Associated with Coral Reef Communities in North Area of Tabuk Region, Saudia Arabia

 , , , , , ,
, , , , , ,  ,
, 
Abstract
1. Introduction
1.1. Diverse Bacterial Symbionts Associated with Corals
1.2. Function of Coral-Associated Microbiome in Coral Life and Health
1.3. Role of Nitrogen-Fixing Microbiome Associated with Corals
1.4. Coral-Associated Microbiome in Red Sea
2. Materials and Methods
2.1. Study Location and Sampling
2.2. Isolation and Quantitative Analysis of DNA
2.3. Setting Up the Library
2.4. Cluster Generation and Sequencing
2.5. Data Generation
2.6. Gene Prediction
2.6.1. Metagenomic Sequencing and Analysis
2.6.2. Taxonomic Annotation
2.6.3. Alpha Diversity Analysis
2.6.4. Statistical Analysis and Data Visualization
3. Results
3.1. Study Site Characteristics
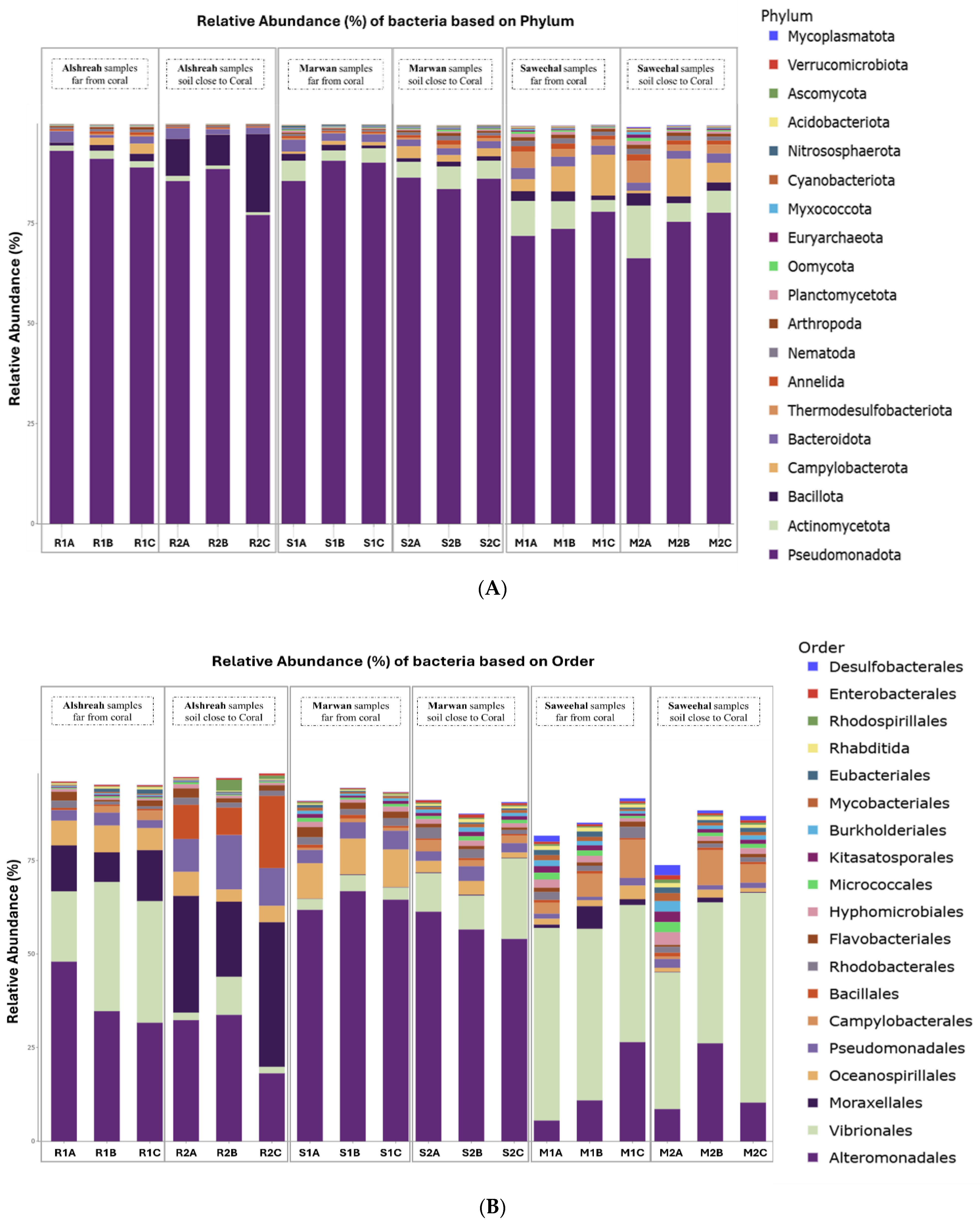
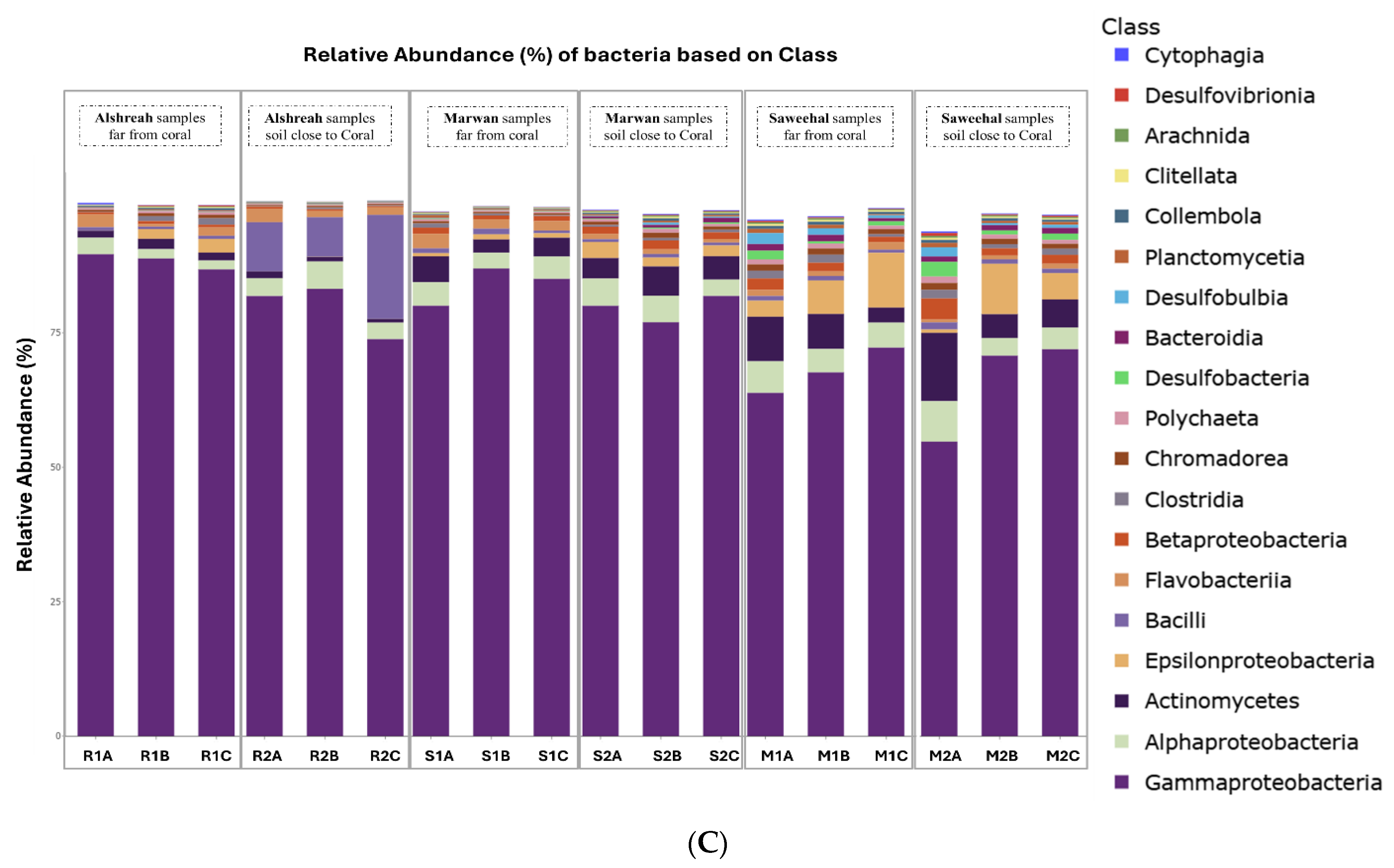
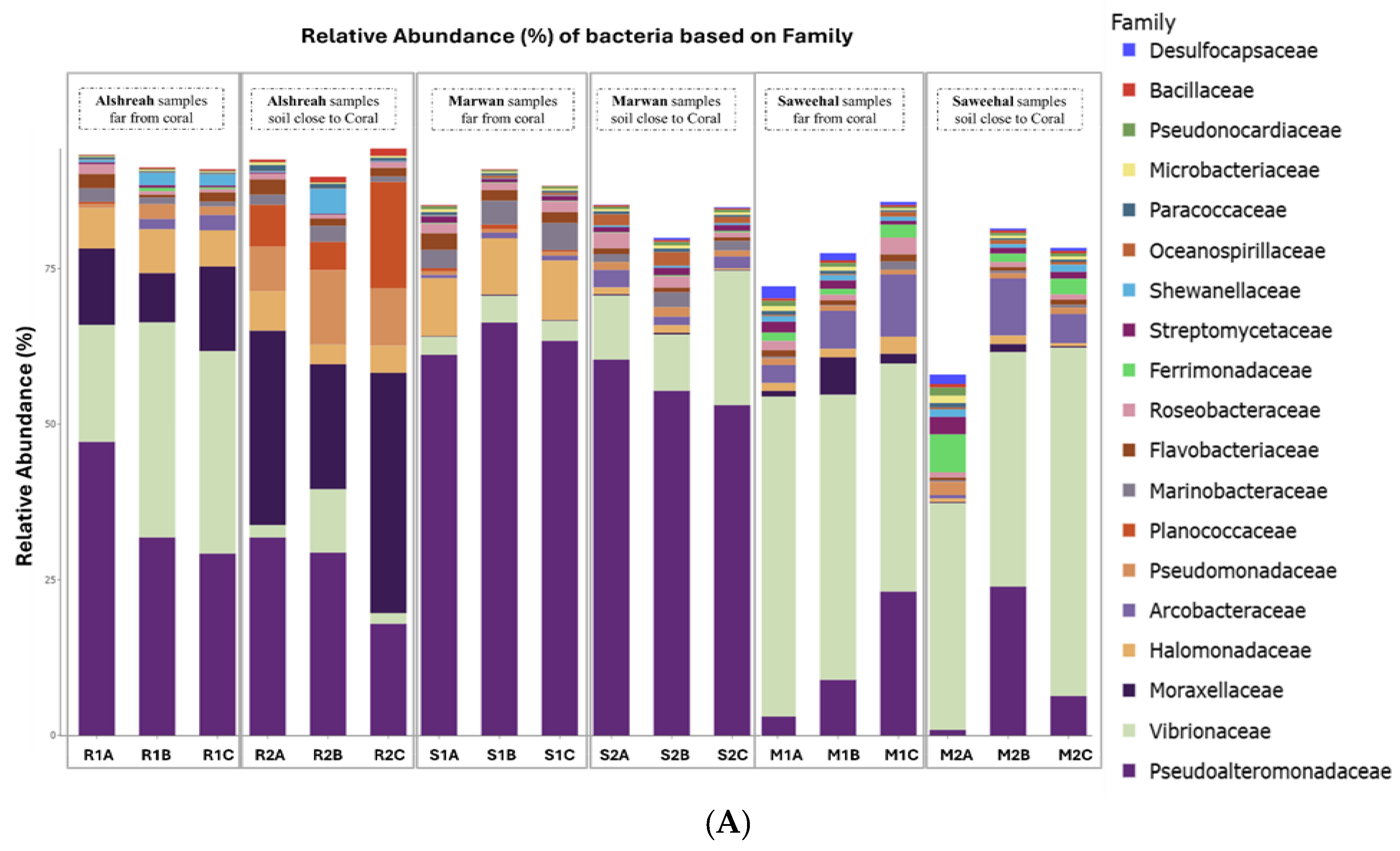
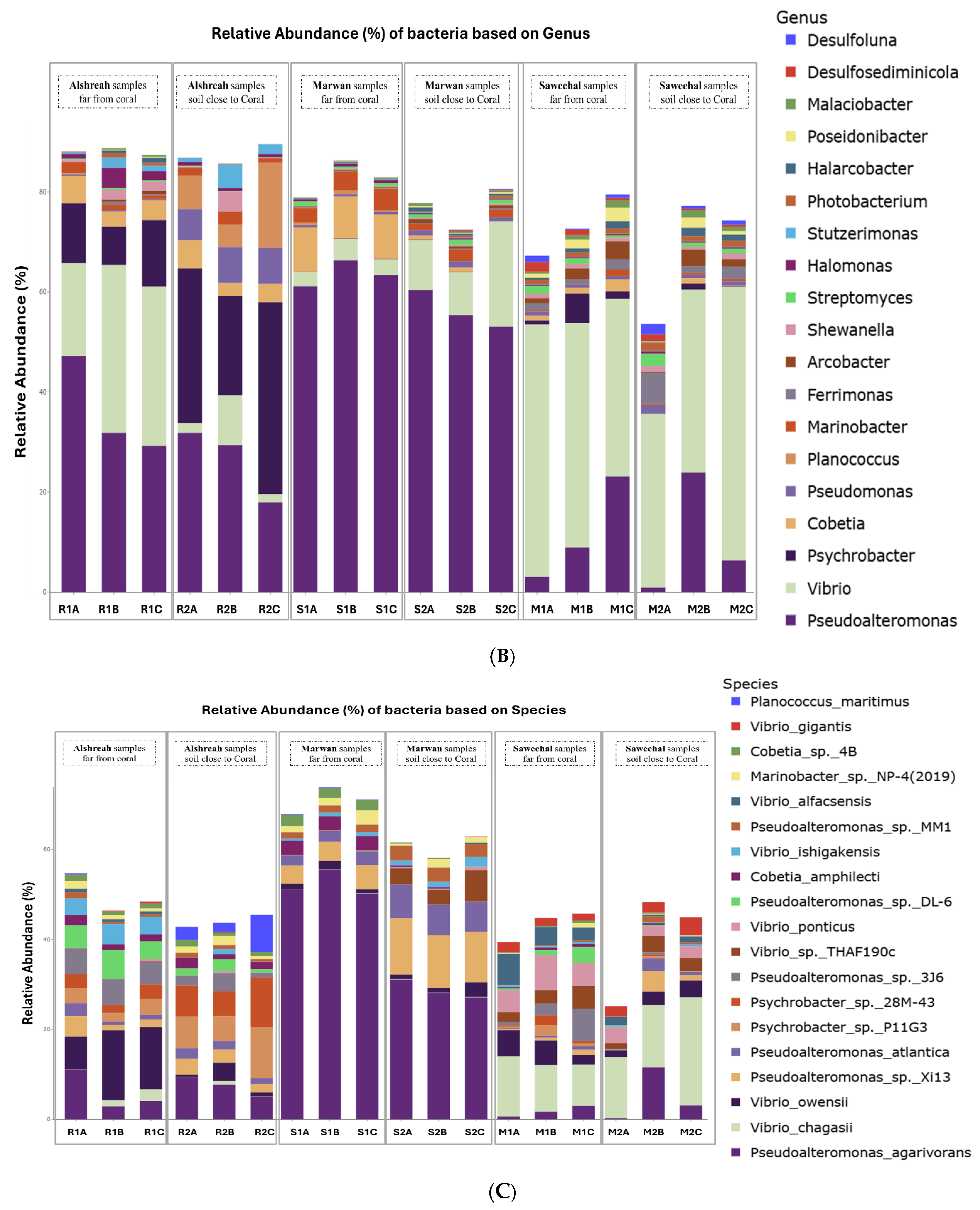
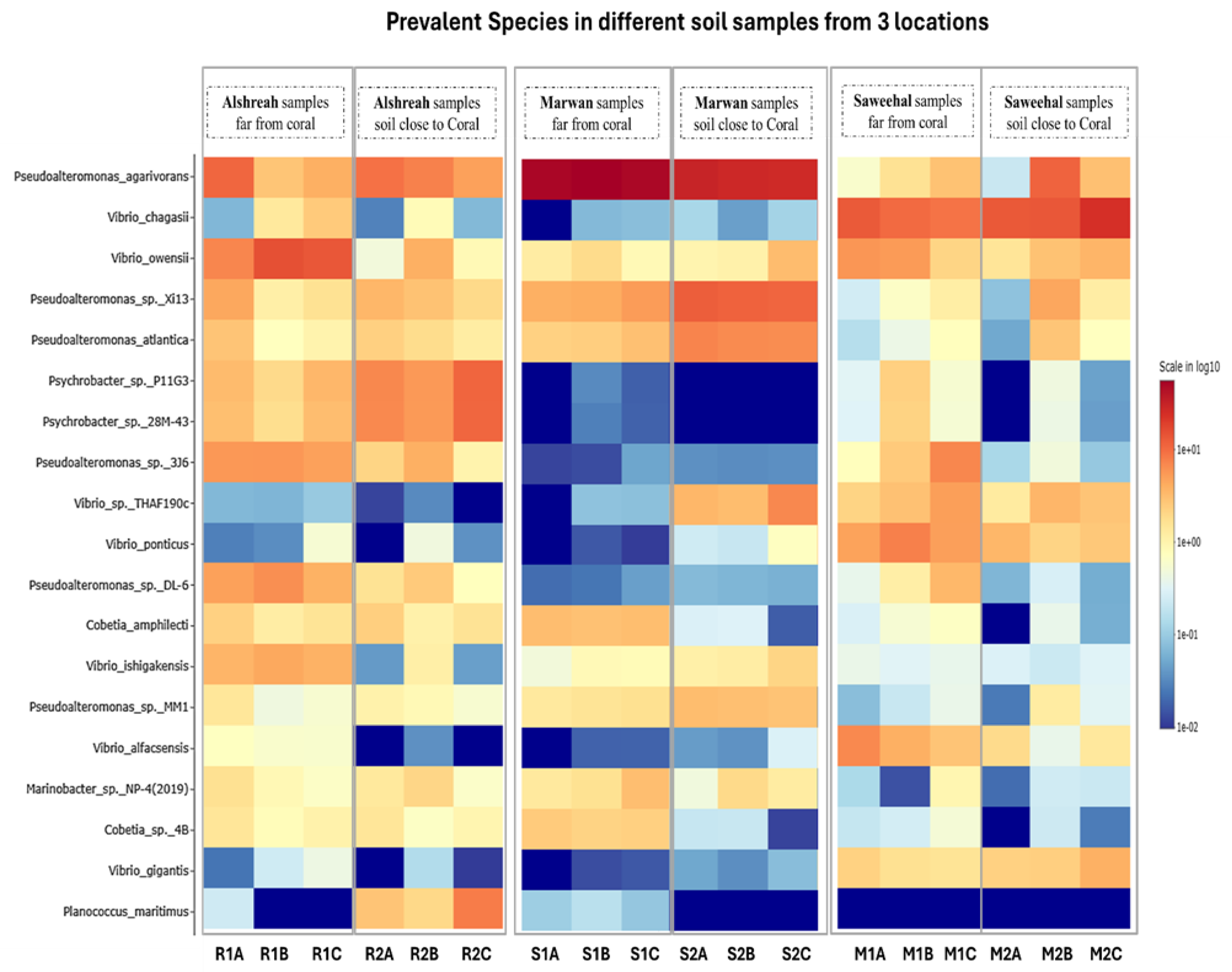
3.2. Taxonomic Composition
3.3. Dominant Microbial Genera per Sample
3.4. Analysis of Bacterial Communities in Coral Proximity and Distances
3.5. Microbial Alpha Diversity
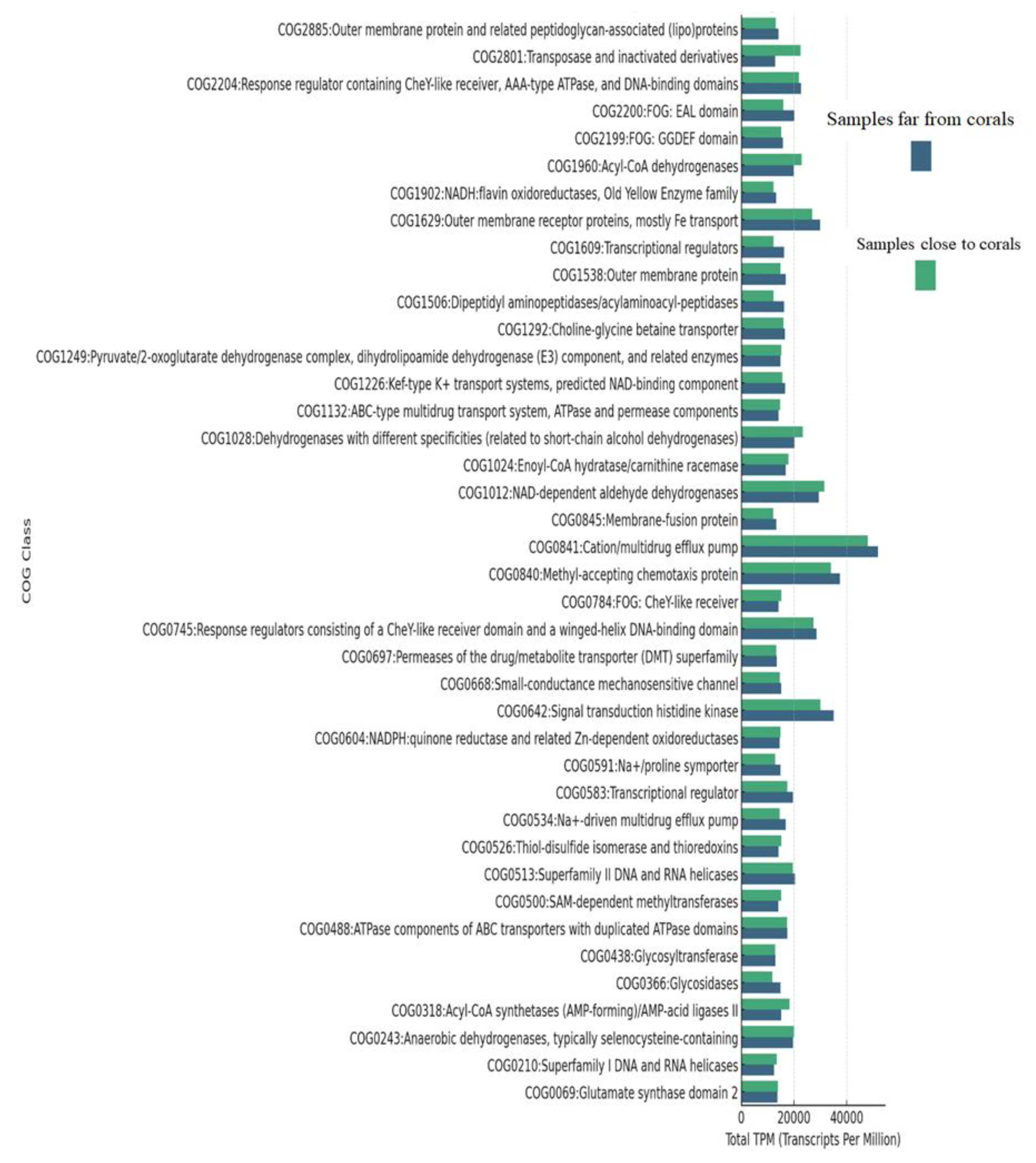
3.6. Clusters of Orthologous Groups (COG) Functional Annotation
3.7. COG Functional Categories in Far-from-Coral Samples
- COG0840: Methyl-accepting chemotaxis protein: This category was present prominently in all the far-from-coral samples, averaging 6000–7147 TPM, which means that this category might be responsible for the adaptation and movement of microorganisms according to environmental stimuli.
- COG0841: Cation/multidrug efflux pump: This functional class showed the highest average TPM values across the far-from-coral samples, ranging from 5358 to 7147. It may reflect involvement in efflux functions and, as such, might be responsible for contributing to resistance in microbial communities of nutrient-limited or stress environments.
- COG0642: Signal transduction histidine kinase: COG group with function related to signal transduction and mechanisms responding to the environment; all have high TPM values; this average is 5051–5533. The presence of such proteins might be indicative of microbes’ adaptation to changing conditions.
- COG1629: Outer membrane receptor proteins, primarily Fe transport: In high abundance in the far-from-coral samples (average TPM = 4200–4533), these proteins can presumably participate in nutrient acquisition, particularly iron, since iron is a limiting element for marine environments.
3.8. COG Functional Categories in Close-to-Coral Samples
- COG0841: Cation/multidrug efflux pump: The category of COG0841, like far-from-coral samples, dominated close-to-coral samples, with TPM ranging from 4165 to 7543. These efflux pumps could be contributing toward cellular integrity and environmental responses, possibly in association with defense mechanisms mediated by coral-associated microbes.
- COG0840: Methyl-accepting chemotaxis protein: Proteins in this category were also highly represented (average TPM = 4338–6524), which reflects their participation in microbial responses to chemical gradients and signaling in the coral-associated environment.
- COG0642: Signal transduction histidine kinase: This category showed a high level of abundance (average TPM = 4220–5166) in coral-associated samples, indicating active regulation of the microbial signaling pathways in the presence of the coral ecosystem.
- COG1012: NAD-dependent aldehyde dehydrogenases: These metabolism and detoxification enzymes were significantly enriched in the near-coral samples (average TPM = 4347–4649), reflecting the metabolic shifts microbes might undergo in response to coral-driven nutrient environments.
- COG0243: Anaerobic dehydrogenases, usually selenocysteine-containing: This functional category has TPM values of about 4124, and this would reflect the microbial adaptation to oxygen-limited environments that could be a reflection of anaerobic or microaerophilic conditions near the coral.
- COG0841/COG0840: The most abundant functional categories between the two environments were COG0841 (Cation/multidrug efflux pump) and COG0840 (Methyl-accepting chemotaxis protein), indicating their key role in microbial survival and adaptation to different environmental conditions. Other important functions include signal transduction (COG0642), which plays a role in response to environmental stimuli, and iron transport (COG1629), which is potentially crucial for nutrient acquisition in both environments. These results give a broad view of the functional roles of microbial communities in both far-from-coral and close-to-coral samples, with key biological processes such as stress response, nutrient transport, and environmental signaling.
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mohamed, A.R.; Ochsenkühn, M.A.; Kazlak, A.M.; Moustafa, A.; Amin, S.A. The coral microbiome: Towards an understanding of the molecular mechanisms of coral–microbiota interactions. FEMS Microbiol. Rev. 2023, 47, fuad005. [Google Scholar] [CrossRef]
- Jaspers, C.; Fraune, S.; Arnold, A.E.; Miller, D.J.; Bosch, T.C.G.; Voolstra, C.R.; Consortium of Australian Academy of Science Boden Research Conference Participants. Resolving structure and function of metaorganisms through a holistic framework combining reductionist and integrative approaches. Zoology 2019, 133, 81–87. [Google Scholar] [PubMed]
- Robbins, S.J.; Singleton, C.M.; Chan, C.X.; Messer, L.F.; Geers, A.U.; Ying, H.; Baker, A.; Bell, S.C.; Morrow, K.M.; Ragan, M.A.; et al. A genomic view of the reef building coral Porites lutea and its microbial symbionts. Nat. Microbiol. 2019, 4, 2090–2100. [Google Scholar] [PubMed]
- Peixoto, R.S.; Sweet, M.; Villela, H.D.M.; Cardoso, P.; Thomas, T.; Voolstra, C.R.; Høj, L.; Bourne, D.G. Coral probiotics: Premise, promise, prospects. Annu. Rev. Anim. Biosci. 2021, 9, 265–288. [Google Scholar]
- Rohwer, F.; Seguritan, V.; Azam, F.; Knowlton, N. Diversity and distribution of coral-associated bacteria. Mar. Ecol. Prog. Ser. 2002, 243, 1–10. [Google Scholar]
- Rosenberg, E.; Koren, O.; Reshef, L.; Efrony, R.; Zilber-Rosenberg, I. The role of microorganisms in coral health, disease and evolution. Nat. Rev. Microbiol. 2007, 5, 355–362. [Google Scholar] [PubMed]
- Rosenberg, E.; Zilber-Rosenberg, I. The hologenome concept of evolution after 10 years. Microbiome 2018, 6, 78. [Google Scholar]
- Zilber-Rosenberg, I.; Rosenberg, E. Microbial-driven genetic variation in holobionts. FEMS Microbiol. Rev. 2021, 45, fuab022. [Google Scholar]
- Muscatine, L. The role of symbiotic algae in carbon and energy flux in reef corals. In Ecosystems of the World; Dubinsky, Z., Ed.; Elsevier: Amsterdam, The Netherlands, 1990; pp. 75–87. [Google Scholar]
- Morris, L.A.; Voolstra, C.R.; Quigley, K.M.; Bourne, D.G.; Bay, L.K. Nutrient availability and metabolism affect the stability of coral–Symbiodiniaceae symbioses. Trends Microbiol. 2019, 27, 678–689. [Google Scholar]
- Bourne, D.G.; Morrow, K.M.; Webster, N.S. Insights into the coral microbiome: Underpinning the health and resilience of reef ecosystems. Annu. Rev. Microbiol. 2016, 70, 317–340. [Google Scholar] [CrossRef]
- Vohsen, S.A.; Herrera, S. Coral microbiomes are structured by environmental gradients in deep waters. Environ. Microbiol. 2024, 19, 38. [Google Scholar]
- Blackall, L.L.; Wilson, B.; van Oppen, M.J. Coral—The world’smost diverse symbiotic ecosystem. Mol. Ecol. 2015, 24, 5330–5347. [Google Scholar] [PubMed]
- Huggett, M.J.; Apprill, A. Coral microbiome database: Integration of sequences reveals high diversity and relatedness of coral associated microbes. Environ. Microbiol. Rep. 2019, 11, 372–385. [Google Scholar] [PubMed]
- Chen, J.; Guo, Y.; Jia, Y.; Liu, G.; Li, D.; Xu, D.; Wang, B.; Zhou, L.; Peng, L.; Zhao, F.; et al. Diversity, function and evolution of marine microbe genomes. bioRxiv 2021. [Google Scholar] [CrossRef]
- Ziegler, M.; Grupstra, C.G.B.; Barreto, M.M.; Eaton, M.; Baomar, J.; Zubier, K.; Al-Sofyani, A.; Turki, A.J.; Ormond, R.; Voolstra, C.R. Coral bacterial community structure responds to environmental change in a host-specific manner. Nat. Commun. 2019, 10, 3092. [Google Scholar]
- Voolstra, C.R.; Ziegler, M. Adapting with microbial help: Microbiome flexibility facilitates rapid responses to environmental change. BioEssays 2020, 42, 2000004. [Google Scholar]
- Meunier, V.; Geissler, L.; Bonnet, S.; Rädecker, N.; Perna, G.; Grosso, O.; Lambert, C.; Rodolfo-Metalpa, R.; Voolstra, C.R.; Houlbrèque, F. Microbes support enhanced nitrogen requirements of coral holobionts in a high CO2 environment. Mol. Ecol. 2021, 30, 5888–5899. [Google Scholar]
- Haas, A.F.; Fairoz, M.F.M.; Kelly, L.W.; Nelson, C.E.; Dinsdale, E.A.; Edwards, R.A.; Giles, S.; Hatay, M.; Hisakawa, N.; Knowles, B.; et al. Global microbialization of coral reefs. Nat. Microbiol. 2016, 1, 16042. [Google Scholar] [CrossRef]
- Prioux, C.; Ferrier-Pages, C.; Deter, J.; Tignat-Perrier, R.; Guilbert, A.; Ballesta, L.; Allemand, D.; Van de Water, J.A.J.M. Insights into the occurrence of phylosymbiosis and co-phylogeny in the holobionts of octocorals from the Mediterranean Sea and Red Sea. Anim. Microbiome 2024, 6, 62. [Google Scholar]
- Vanwonterghem, I.; Webster, N.S. Coral reef microorganisms in a changing climate. iScience 2020, 23, 100972. [Google Scholar]
- Wangpraseurt, D.; Jacques, S.L.; Petrie, T.; Kühl, M. Monte Carlo modeling of photon propagation reveals highly scattering coral tissue. Front. Plant Sci. 2016, 7, 1404. [Google Scholar] [CrossRef] [PubMed]
- Pernice, M.; Raina, J.-B.; Rädecker, N.; Cárdenas, A.; Pogoreutz, C.; Voolstra, C.R. Down to the bone: The role of overlooked endolithic microbiomes in reef coral health. ISME J. 2020, 14, 325–334. [Google Scholar] [CrossRef] [PubMed]
- Sweet, M.J.; Croquer, A.; Bythell, J.C. Bacterial assemblages differ between compartments within the coral holobiont. Coral Reefs 2010, 30, 39–52. [Google Scholar] [CrossRef]
- Leite, D.C.A.; Salles, J.F.; Calderon, E.N.; Castro, C.B.; Bianchini, A.; Marques, J.A.; van Elsas, J.D.; Peixoto, R.S. Coral bacterial-core abundance and network complexity as proxies for anthropogenic pollution. Front. Microbiol. 2018, 9, 833. [Google Scholar] [CrossRef]
- Ide, K.; Nishikawa, Y.; Maruyama, T.; Tsukada, Y.; Kogawa, M.; Takeda, H.; Ito, H.; Wagatsuma, R.; Miyaoka, R.; Nakano, Y.; et al. Targeted single-cell genomics reveals novel host adaptation strategies of the symbiotic bacteria Endozoicomonas in Acropora tenuis coral. Microbiome 2022, 10, 220. [Google Scholar] [CrossRef] [PubMed]
- Pogoreutz, C.; Voolstra, C.R.; Rädecker, N.; Weis, V.; Cardenas, A.; Raina, J.-B. The coral holobiont highlights the dependence of cnidarian animal hosts on their associated microbes. In Cellular Dialogues in the Holobiont; Bosch, T.C.G., Hadfield, M.G., Eds.; CRC Press: Boca Raton, FL, USA, 2020. [Google Scholar]
- Falkowski, P.G.; Dubinsky, Z.; Muscatine, L.; McCloskey, L. Population control in symbiotic corals. Bioscience 1993, 43, 606–611. [Google Scholar] [CrossRef]
- Cunning, R.; Muller, E.B.; Gates, R.D.; Nisbet, R.M. A dynamic bioenergetic model for coral–Symbiodinium symbioses and coral bleaching as an alternate stable state. J. Theor. Biol. 2017, 431, 49–62. [Google Scholar] [CrossRef]
- Muscatine, L.; Porter, J.W. Reef corals: Mutualistic symbioses adapted to nutrient-poor environments. Bioscience 1977, 27, 454–460. [Google Scholar] [CrossRef]
- Lema, K.A.; Willis, B.L.; Bourne, D.G. Corals form characteristic associations with symbiotic nitrogen-fixing bacteria. Appl. Environ. Microbiol. 2012, 78, 3136–3144. [Google Scholar] [CrossRef]
- Wang, J.; Douglas, A. Essential amino acid synthesis and nitrogen recycling in an alga–invertebrate symbiosis. Mar. Biol. 1999, 135, 219–222. [Google Scholar] [CrossRef]
- Yellowlees, D.; Rees, T.A.V.; Leggat, W. Metabolic interactions between algal symbionts and invertebrate hosts. Plant Cell Environ. 2008, 31, 679–694. [Google Scholar] [CrossRef]
- Reynaud, S.; Martinez, P.; Houlbrèque, F.; Billy, I.; Allemand, D.; Ferrier-Pagès, C. Effect of light and feeding on the nitrogen isotopic composition of a zooxanthellate coral: Role of nitrogen recycling. Mar. Ecol. Prog. Ser. 2009, 392, 103–110. [Google Scholar] [CrossRef]
- Olson, N.D.; Lesser, M.P. Diazotrophic diversity in the Caribbean coral, Montastraea cavernosa. Arch. Microbiol. 2013, 195, 853–859. [Google Scholar] [CrossRef] [PubMed]
- Lema, K.A.; Bourne, D.G.; Willis, B.L. Onset and establishment of diazotrophs and other bacterial associates in the early life history stages of the coral Acropora millepora. Mol. Ecol. 2014, 23, 4682–4695. [Google Scholar] [CrossRef]
- Kimes, N.E.; Van Nostrand, J.D.; Weil, E.; Zhou, J.; Morris, P.J. Microbial functional structure of Montastraea faveolata, an important Caribbean reef building coral, differs between healthy and yellow-band diseased colonies. Environ. Microbiol. 2010, 12, 541–556. [Google Scholar] [CrossRef] [PubMed]
- Rädecker, N.; Pogoreutz, C.; Gegner, H.M.; Cárdenas, A.; Perna, G.; Geißler, L.; Roth, F.; Bougoure, J.; Guagliardo, P.; Struck, U.; et al. Heat stress reduces the contribution of diazotrophs to coral holobiont nitrogen cycling. ISME J. 2022, 16, 1110–1118. [Google Scholar] [CrossRef] [PubMed]
- Seymour, J.R.; Amin, S.A.; Raina, J.B.; Stocker, R. Zooming in on the phycosphere: The ecological interface for phytoplankton–bacteria relationships. Nat. Microbiol. 2017, 2, 17065. [Google Scholar] [CrossRef]
- Cirri, E.; Pohnert, G. Algae–bacteria interactions that balance the planktonic microbiome. New Phytol. 2019, 223, 100–106. [Google Scholar] [CrossRef]
- Delgadillo-Ordoñez, N.; Raimundo, I.; Barno, A.R.; Osman, E.O.; Villela, H.; Bennett-Smith, M.; Voolstra, C.R.; Benzoni, F.; Peixoto, R.S. Red Sea Atlas of Coral-Associated Bacteria Highlights Common Microbiome Members and Their Distribution across Environmental Gradients. Microorganisms 2022, 10, 2340. [Google Scholar] [CrossRef]
- Osman, E.O.; Suggett, D.J.; Voolstra, C.R.; Pettay, D.T.; Clark, D.R.; Pogoreutz, C.; Sampayo, E.M.; Warner, M.E.; Smith, D.J. Coral Microbiome composition along the northern Red Sea suggests high plasticity of bacterial and specificity of endosymbiotic dinoflagellate communities. Microbiome 2020, 8, 8. [Google Scholar]
- Ziegler, M.; Roik, A.; Porter, A.; Zubier, K.; Mudarris, M.S.; Ormond, R.; Voolstra, C.R. Coral microbial community dynamics in response to anthropogenic impacts near a major city in the Central Red Sea. Mar. Pollut. Bull. 2016, 105, 629–640. [Google Scholar] [CrossRef] [PubMed]
- Raimundo, I.; Rosado, P.M.; Barno, A.R.; Antony, C.P.; Peixoto, R.S. Unlocking the genomic potential of Red Sea coral probiotics. Sci. Rep. 2024, 14, 14514. [Google Scholar] [CrossRef] [PubMed]
- Winkler, R.; Antonius, A.; Renegar, D.A. The skeleton eroding band disease on coral reefs of Aqaba, Red Sea. Mar. Ecol. 2004, 25, 129–144. [Google Scholar] [CrossRef]
- Roik, A.; Röthig, T.; Order, C.; Ziegler, M.; Kremb, S.G.; Voolstra, C.R. Year-long monitoring of Physico-chemical and biological variables provide a comparative baseline of coral reef functioning in the Central Red Sea. PLoS ONE 2016, 11, e0163939. [Google Scholar] [CrossRef]
- Röthig, T.; Ochsenkühn, M.A.; Roik, A.; van der Merwe, R.; Voolstra, C.R. Long-term salinity tolerance is accompanied by major restructuring of the coral bacterial microbiome. Mol. Ecol. 2016, 25, 1308–1323. [Google Scholar] [CrossRef]
- Röthig, T.; Roik, A.; Yum, L.K.; Voolstra, C.R. Distinct bacterial microbiomes associate with the Deep-Sea coral Eguchipsammia fistula from the Red Sea and from aquaria settings. Front. Mar. Sci. 2017, 4, 259. [Google Scholar] [CrossRef]
- Isaza, D.F.G.; Cramp, R.L.; Franklin, C.E. Living in polluted waters: A meta-analysis of the effects of nitrate and interactions with other environmental stressors on freshwater taxa. Environ. Pollut. 2020, 261, 114091. [Google Scholar] [CrossRef]
- Li, D.; Liu, C.M.; Luo, R.; Sadakane, K.; Lam, T.W. MEGAHIT: An ultra-fast single-node solution for large and complex metagenomics assembly via succinct de Bruijn graph. Bioinformatics 2015, 31, 1674–1676. [Google Scholar] [CrossRef]
- Newton, R.J.; Jones, S.E.; Eiler, A.; McMahon, K.D.; Bertilsson, S. A guide to the natural history of freshwater lake bacteria. Microbiol. Mol. Biol. Rev. 2011, 75, 14–49. [Google Scholar] [CrossRef]
- Hyatt, D.; Chen, G.L.; Locascio, P.F.; Land, M.L.; Larimer, F.W.; Hauser, L.J. Prodigal: Prokaryotic gene recognition and translation initiation site identification. BMC Bioinform. 2010, 11, 119. [Google Scholar] [CrossRef]
- Wu, Y.-W.; Simmons, B.A.; Singer, S.W. MaxBin 2.0: An automated binning algorithm to recover genomes from multiple metagenomic datasets. Bioinformatics 2016, 32, 605–607. [Google Scholar] [CrossRef] [PubMed]
- Menzel, P.; Ng, K.L.; Krogh, A. Fast and sensitive taxonomic classification for metagenomics with Kaiju. Nat. Commun. 2016, 7, 11257. [Google Scholar] [CrossRef]
- McMurdie, P.J.; Holmes, S. phyloseq: An R Package for Reproducible Interactive Analysis and Graphics of Microbiome Census Data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef]
- Wickham, H.; Averick, M.; Bryan, J.; Chang, W.; McGowan, L.D.A.; François, R.; Grolemund, G.; Hayes, A.; Henry, L.; Hester, J.; et al. Welcome to the Tidyverse. J. Open Source Softw. 2019, 4, 1686. [Google Scholar] [CrossRef]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar] [CrossRef]
- Gu, Z.; Eils, R.; Schlesner, M. Complex heatmaps reveal patterns and correlations in multidimensional genomic data. Bioinformatics 2016, 32, 2847–2849. [Google Scholar] [CrossRef]
- Sweet, M.; Villela, H.; Keller-Costa, T.; Costa, R.; Romano, S.; Bourne, D.G.; Cárdenas, A.; Huggett, M.J.; Kerwin, A.H.; Kuek, F.; et al. Insights into the cultured bacterial fraction of corals. mSystems 2021, 6, e0124920. [Google Scholar] [CrossRef] [PubMed]
- Pollock, F.J.; McMinds, R.; Smith, S.; Bourne, D.G.; Willis, B.L.; Medina, M.; Thurber, R.V.; Zaneveld, J.R. Coral-associated bacteria demonstrate phylosymbiosis and co- phylogeny. Nat. Commun. 2018, 9, 4921. [Google Scholar]
- Sato, Y.; Civiello, M.; Bell, S.C.; Willis, B.L.; Bourne, D.G. Integrated approach to understanding the onset and pathogenesis of black band disease in corals. Environ. Microbiol. 2016, 18, 752–756. [Google Scholar] [CrossRef]
- Moriarty, T.; Leggat, W.; Huggett, M.J.; Ainsworth, T. Coral disease causes, consequences, and risk within coral restoration. Trends Microbiol. 2020, 28, 793–807. [Google Scholar]
- Zhang, J.; Chen, M.; Huang, J.; Guo, X.; Zhang, Y.; Liu, D.; Wu, R.; He, H.; Wang, J. Diversity of the microbial community and cultivable protease-producing bacteria in the sediments of the Bohai Sea, Yellow Sea and South China Sea. PLoS ONE 2019, 14, e0215328. [Google Scholar]
- Ye, Q.; Wu, Y.; Zhu, Z.; Wang, X.; Li, Z.; Zhang, J. Bacterial diversity in the surface sediments of the hypoxic zone near the Changjiang Estuary and in the East China Sea. Microbiol. Open 2016, 5, 323–339. [Google Scholar] [CrossRef] [PubMed]
- Chung, H.W.; Liu, C.C.; Chiu, Y.S.; Liu, J.T. Spatiotemporal variation of Gaoping River plume observed by Formosat-2 high resolution imagery. J. Mar. Syst. 2014, 132, 28–37. [Google Scholar] [CrossRef]
- Bhattacharyya, A.; Haldar, A.; Bhattacharyya, M.; Ghosh, A. Anthropogenic influence shapes the distribution of antibiotic-resistant bacteria (ARB) in the sediment of Sundarban estuary in India. Sci. Total Environ. 2019, 647, 1626–1639. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Qian, P.Y. Conservative fragments in bacterial 16S rRNA genes and primer design for 16S ribosomal DNA amplicons in metagenomic studies. PLoS ONE 2009, 4, e7401. [Google Scholar] [CrossRef]
- Zhou, M.-Y.; Wang, G.-L.; Li, D.; Zhao, D.-L.; Qin, Q.-L.; Chen, X.-L.; Chen, B.; Zhou, B.-C.; Zhang, X.-Y.; Zhang, Y.-Z. Diversity of both the cultivable protease-producing bacteria and bacterial extracellular proteases in the coastal sediments of King George Island, Antarctica. PLoS ONE 2013, 8, e79668. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.Y.; Han, X.X.; Chen, X.L.; Dang, H.Y.; Xie, B.B.; Qin, Q.L.; Shi, M.; Zhou, B.C.; Zhang, Y.Z. Diversity of cultivable protease-producing bacteria in sediments of Jiaozhou Bay, China. Front. Microbiol. 2015, 6, 1021. [Google Scholar] [CrossRef]
- Röthig, T.; Yum, L.K.; Kremb, S.G.; Roik, A.; Voolstra, C.R. Microbial community composition of deep-sea corals from the Red Sea provides insight into functional adaption to a unique environment. Sci. Rep. 2017, 7, 44714. [Google Scholar] [CrossRef]
- Neave, M.J.; Michell, C.T.; Apprill, A.; Voolstra, C.R. Endozoicomonas genomes reveal functional adaptation and plasticity in bacterial strains symbiotically associated with diverse marine hosts. Sci. Rep. 2017, 7, 40579. [Google Scholar] [CrossRef]
- Rosado, P.M.; Leite, D.C.A.; Duarte, G.A.S.; Chaloub, R.M.; Jospin, G.; da Rocha, U.N.; Saraiva, J.P.; Dini-Andreote, F.; Eisen, J.A.; Bourne, D.G.; et al. Marine probiotics: Increasing coral resistance to bleaching through microbiome manipulation. ISME J. 2019, 13, 921–936. [Google Scholar] [CrossRef]
- Tandon, K.; Lu, C.-Y.; Chiang, P.-W.; Wada, N.; Yang, S.-H.; Chan, Y.-F.; Chen, P.-Y.; Chang, H.-Y.; Chiou, Y.-J.; Chou, M.-S.; et al. Comparative genomics: Dominant coral-bacterium Endozoicomonas acroporae metabolizes dimethyl sulfoniopropionate (DMSP). ISME J. 2020, 14, 1290–1303. [Google Scholar] [CrossRef]
- Hopkins, F.E.; Bell, T.G.; Yang, M.; Suggett, D.J.; Steinke, M. Air exposure of coral is a significant source of dimethylsulfide (DMS) to the atmosphere. Sci. Rep. 2016, 6, 36031. [Google Scholar] [CrossRef]
- Peixoto, R.S.; Rosado, P.M.; de Assis Leite, D.C.; Rosado, A.S.; Bourne, D.G. Beneficial Microorganisms for Corals (BMC): Proposed mechanisms for coral health and resilience. Front. Microbiol. 2017, 8, 341. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Shinde, P.B.; Lee, H.J.; Yoo, E.S.; Lee, C.-O.; Hong, J.; Choi, S.H.; Jung, J.H. Bile acid derivatives from a sponge-associated bacterium Psychrobacter sp. Arch. Pharm. Res. 2009, 32, 857–862. [Google Scholar] [CrossRef] [PubMed]
- Gardner, S.G.; Nielsen, D.A.; Laczka, O.; Shimmon, R.; Beltran, V.H.; Ralph, P.J.; Petrou, K. Dimethylsulfoniopropionate, superoxide dismutase and glutathione as stress response indicators in three corals under short-term hyposalinity stress. Proc. R Soc. B Biol. Sci. 2016, 283, 20152418. [Google Scholar] [CrossRef] [PubMed]
- Shore-Maggio, A.; Aeby, G.S.; Callahan, S.M. Influence of salinity and sedimentation on Vibrio infection of the Hawaiian Coral Montipora capitata. Dis. Aquat. Org. 2018, 128, 63–71. [Google Scholar] [CrossRef]
- Ceh, J.; Kilburn, M.R.; Cliff, J.B.; Raina, J.-B.B.; Van Keulen, M.; Bourne, D.G. Nutrient cycling in early coral life stages: Pocillopora damicornis larvae provide their algal symbiont (Symbiodinium) with nitrogen acquired from bacterial associates. Ecol. Evol. 2013, 3, 2393–2400. [Google Scholar] [CrossRef]
- van Oppen, M.J.H.; Blackall, L.L. Coral microbiome dynamics, functions and design in a changing world. Nat. Rev. Microbiol. 2019, 17, 557–567. [Google Scholar] [CrossRef]
- Raimundo, I.; Silva, R.; Meunier, L.; Valente, S.M.; Lago-Lestón, A.; Keller-Costa, T.; Costa, R. Functional metagenomics reveals differential chitin degradation and utilization features across free-living and host-associated marine microbiomes. Microbiome 2021, 9, 43. [Google Scholar] [CrossRef]
- Santoro, E.P.; Borges, R.M.; Espinoza, J.L.; Freire, M.; Messias, C.S.M.A.; Villela, H.D.M.; Pereira, L.M.; Vilela, C.L.S.; Rosado, J.G.; Cardoso, P.M.; et al. Coral microbiome manipulation elicits metabolic and genetic restructuring to mitigate heat stress and evade mortality. Sci. Adv. 2021, 7, eabg3088. [Google Scholar] [CrossRef]
- Huang, J.; Zeng, B.; Liu, D.; Wu, R.; Zhang, J.; Liao, B.; He, H.; Bian, F. Classification and Structural Insight into Vibriolysin-Like Proteases of Vibrio Pathogenicity. Microb. Pathog. 2018, 117, 335–340. [Google Scholar] [CrossRef]
- Kabbara, S.; Hérivaux, A.; Dugé de Bernonville, T.; Courdavault, V.; Clastre, M.; Gastebois, A.; Osman, M.; Hamze, M.; Cock, J.M.; Schaap, P.; et al. Diversity and evolution of sensor histidine kinases in eukaryotes. Genome Biol. Evolut. 2019, 11, 86–108. [Google Scholar]
- Zhang, Y.; Ip, J.C.; Xie, J.Y.; Yeung, Y.H.; Sun, Y.; Qiu, J.W. Host-symbiont transcriptomic changes during natural bleaching and recovery in the leaf coral Pavona decussata. Sci. Total Environ. 2021, 806, 150656. [Google Scholar]
- Wild, C.; Holger, W.; Huettel, M. Influence of coral mucus on nutrient fluxes in carbonate sands. Mar. Ecol. Prog. Ser. 2005, 287, 87–98. [Google Scholar] [CrossRef]
- Dennis, P.G.; Seymour, J.; Kumbun, K.; Tyson, G.W. Diverse populations of lake water bacteria exhibit chemotaxis towards inorganic nutrients. ISME J. 2013, 7, 1661–1664. [Google Scholar] [PubMed]
- Mustafa, G.A.; Abd-Elgawad, A.; Abdel-Haleem, A.M.; Siam, R. Egypt’s Red Sea coast: Phylogenetic analysis of cultured microbial consortia in industrialized sites. Front. Microbiol. 2014, 5, 363. [Google Scholar] [CrossRef]
- Faust, K.; Lahti, L.; Gonze, D.; de Vos, W.M.; Raes, J. Metagenomics meets time series analysis: Unraveling microbial community dynamics. Curr. Opin. Microbiol. 2015, 25, 56–66. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Water Parameters | Average in Site 1 | Average in Site 2 | Average in Site 3 | Average in Site 4 | Average in Site 4 | Average in Site 5 | Average in Site 6 | Average in Site 7 | Average in Site 8 | Average in Site 9 |
|---|---|---|---|---|---|---|---|---|---|---|
| pH | 8.1 | 8.3 | 8.0 | 7.3 | 8.3 | 8.4 | 7.2 | 8.1 | 8.4 | 8.3 |
| Temperature °C | 28.7 | 28.4 | 29.7 | 29.3 | 28.0 | 29.8 | 29.4 | 29.5 | 28.8 | 29.6 |
| Turbidity (NTU) | 13 | 14 | 12 | 13 | 13 | 11 | 14 | 15 | 14 | 16 |
| DO (mg/L) | 3.36 | 3.54 | 3.31 | 3.44 | 3.72 | 3.46 | 3.33 | 3.34 | 3.52 | 3.50 |
| Salinity ppt | 44.5 | 43.3 | 43.6 | 44.4 | 43.0 | 44.0 | 44.3 | 44.2 | 43.47 | 44.2 |
| Genus | Alshreah | Saweehal | Marwan | |||
|---|---|---|---|---|---|---|
| R1A-R1C (%) Far from Corals | R2A-R2C (%) Close to Corals | S1A-S1C (%) Far from Corals | S2A-S2C (%) Close to Corals | M1A-M1C (%) Far from Corals | M2A-M2C (%) Close to Corals | |
| Pseudoalteromonas | 36 | 26 | 63 | 57 | 13 | 10 |
| Vibrio | 28 | 8 | 3 | 8 | 44 | 42 |
| Psychrobacter | 11 | 29 | ~1 | ~1 | 2 | 2 |
| Cobetia | 4 | 4 | 9 | 7 | 3 | 2 |
| Pseudomonas | ~1 | 7 | ~1 | ~1 | 3 | 4 |
| Species | Alshreah Far (%) | Alshreah Close (%) | Saweehal Far (%) | Saweehal Close (%) | Marwan Far (%) | Marwan Close (%) | Far-From-Coral (%) | Close-to-Coral (%) |
|---|---|---|---|---|---|---|---|---|
| Pseudoalteromonas agarivorans | 5.99 | 7.35 | 52.28 | 28.77 | 1.75 | 4.94 | 20.00 | 13.6 |
| Vibrio chagasii | 1.35 | 0.31 | 0.05 | 0.1 | 10.98 | 17.2 | 4.12 | 5.87 |
| Vibrio owensii | 12.24 | 1.8 | 1.34 | 1.79 | 4.49 | 2.73 | 6.02 | 2.10 |
| Pseudoalteromonas sp. Xi13 | 2.47 | 2.84 | 4.55 | 11.84 | 0.71 | 1.97 | 2.57 | 5.55 |
| Pseudoalteromonas atlantica | 1.55 | 1.82 | 2.58 | 6.93 | 0.45 | 1.2 | 1.52 | 3.31 |
| Psychrobacter sp. P11G3 | 2.96 | 7.98 | 0.02 | 0 | 1.07 | 0.17 | 1.35 | 2.71 |
| Psychrobacter sp. 28M-43 | 2.72 | 7.86 | 0.02 | 0 | 1.03 | 0.16 | 1.25 | 2.67 |
| Pseudoalteromonas sp. 3J6 | 5.46 | 2.4 | 0.03 | 0.04 | 3.5 | 0.24 | 2.99 | 0.89 |
| Vibrio sp. THAF190c | 0.08 | 0.02 | 0.06 | 4.63 | 3.45 | 2.59 | 1.19 | 2.41 |
| Vibrio ponticus | 0.21 | 0.17 | 0.01 | 0.39 | 5.93 | 2.81 | 2.05 | 1.12 |
| Sample ID | Observed | Chao1 | ACE | Shannon | Simpson | Fisher |
|---|---|---|---|---|---|---|
| Alshreah Far (R1A, R1B, R1C) | 5.99 | 7.35 | 52.28 | 28.77 | 1.75 | 4.94 |
| Alshreah Close (R2A, R2B, R2C) | 5.46 | 2.4 | 2.57 | 5.55 | 2.99 | 0.89 |
| Saweehal Far (S1A, S1B, S1C) | 52.28 | 1.75 | 0.31 | 2.12 | 5.87 | 0.89 |
| Saweehal Close (S2A, S2B, S2C) | 4.94 | 3.31 | 1.52 | 5.55 | 2.77 | 0.88 |
| Marwan Far (M1A, M1B, M1C) | 1.75 | 4.12 | 1.75 | 4.94 | 2.94 | 2 |
| Marwan Close (M2A, M2B, M2C) | 4.94 | 1.75 | 4.94 | 2.1 | 0.95 | 1.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ghobashy, M.O.I.; Al-otaibi, A.S.; Alharbi, B.M.; Alshehri, D.; Ghabban, H.; Albalawi, D.A.; Alenzi, A.M.; Alatawy, M.; Alatawi, F.A.; Algammal, A.M.; et al. Metagenomic Characterization of Microbiome Taxa Associated with Coral Reef Communities in North Area of Tabuk Region, Saudia Arabia. Life 2025, 15, 423. https://doi.org/10.3390/life15030423
Ghobashy MOI, Al-otaibi AS, Alharbi BM, Alshehri D, Ghabban H, Albalawi DA, Alenzi AM, Alatawy M, Alatawi FA, Algammal AM, et al. Metagenomic Characterization of Microbiome Taxa Associated with Coral Reef Communities in North Area of Tabuk Region, Saudia Arabia. Life. 2025; 15(3):423. https://doi.org/10.3390/life15030423
Chicago/Turabian StyleGhobashy, Madeha O. I., Amenah S. Al-otaibi, Basmah M. Alharbi, Dikhnah Alshehri, Hanaa Ghabban, Doha A. Albalawi, Asma Massad Alenzi, Marfat Alatawy, Faud A. Alatawi, Abdelazeem M. Algammal, and et al. 2025. "Metagenomic Characterization of Microbiome Taxa Associated with Coral Reef Communities in North Area of Tabuk Region, Saudia Arabia" Life 15, no. 3: 423. https://doi.org/10.3390/life15030423
APA StyleGhobashy, M. O. I., Al-otaibi, A. S., Alharbi, B. M., Alshehri, D., Ghabban, H., Albalawi, D. A., Alenzi, A. M., Alatawy, M., Alatawi, F. A., Algammal, A. M., Mir, R., & Mahrous, Y. M. (2025). Metagenomic Characterization of Microbiome Taxa Associated with Coral Reef Communities in North Area of Tabuk Region, Saudia Arabia. Life, 15(3), 423. https://doi.org/10.3390/life15030423