
The Neuroscience Behind Writing: Handwriting vs. Typing—Who Wins the Battle?


 , , , , ,
, , , , , 
 ,
,  ,
, 

Abstract

1. Introduction
1.1. Handwriting Versus Typing
1.2. Aim of the Review
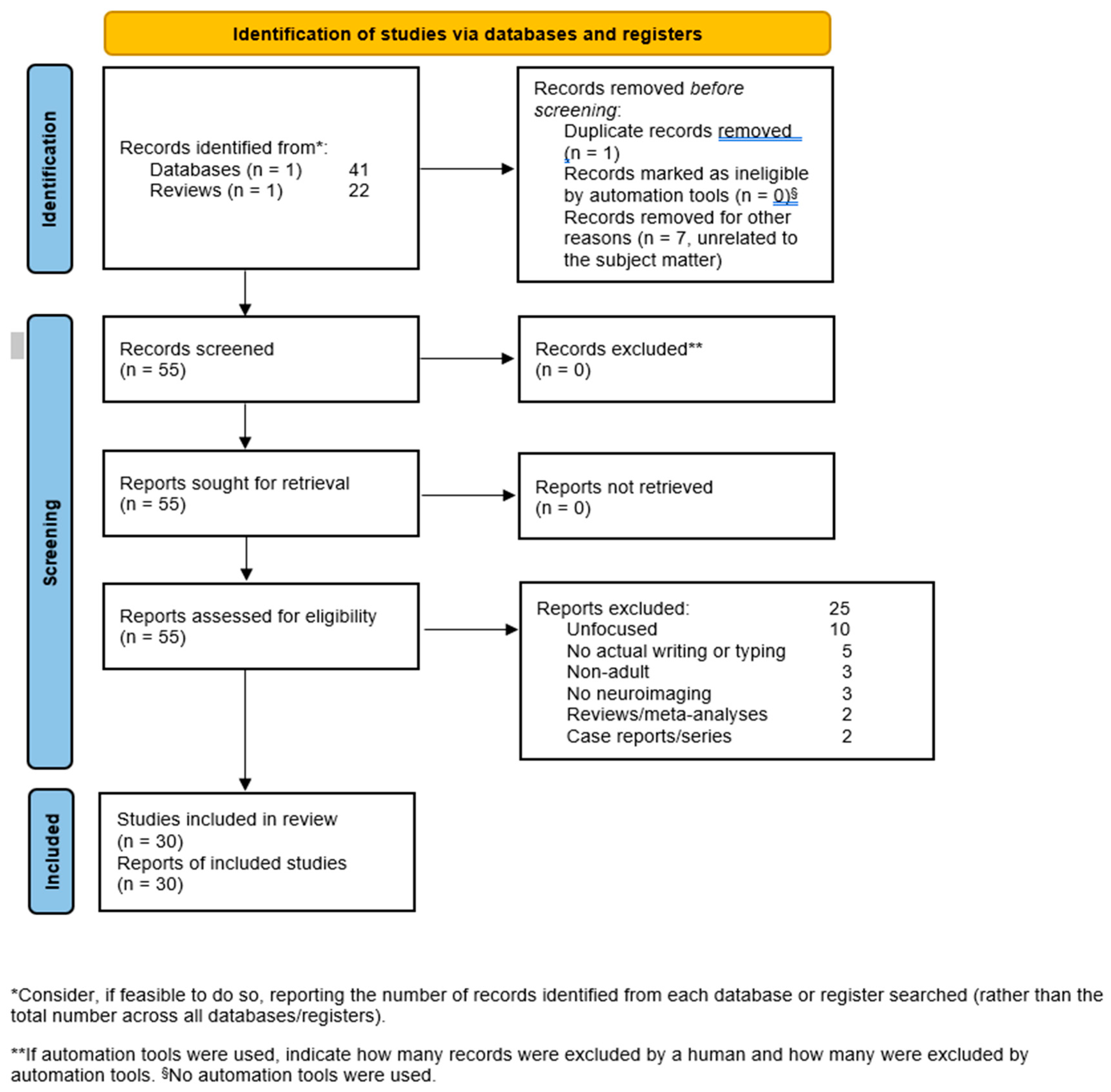
2. Materials and Methods
3. Results
4. Discussion
4.1. Motor Control and Cognitive Engagement
4.2. Cognitive Processing: Memory and Learning Outcomes
4.3. Functional Connectivity and Neural Integration
4.4. Educational Implications
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Marano, G.; Traversi, G.; Gaetani, E.; Sani, G.; Mazza, S.; Mazza, M. Graphology: An Interface Between Biology, Psychology and Neuroscience. Psychol. Disord. Res. 2019, 3, 3–13. [Google Scholar] [CrossRef]
- Gowda, M.R.; Harish, N.; Aslam, A.; Padmanabiah, M.; Magaji, R. Handprints of the mind: Decoding personality traits and handwritings. Indian J. Psychol. Med. 2015, 37, 409–412. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Mak, H.Y.; Wang, J.; Cai, Q. How Characters Are Learned Leaves Its Mark on the Neural Substrates of Chinese Reading. eNeuro 2022, 9, ENEURO.0111-22.2022. [Google Scholar] [CrossRef]
- Brewer, J.F. Graphology. Complement. Ther. Nurs. Midwifery 1999, 5, 6–14. [Google Scholar] [CrossRef] [PubMed]
- Planton, S.; Longcamp, M.; Péran, P.; Démonet, J.F.; Jucla, M. How specialized are writing-specific brain regions? An fMRI study of writing, drawing and oral spelling. Cortex 2017, 88, 66–80. [Google Scholar] [CrossRef] [PubMed]
- Venturelli, A. Dal Gesto alla Scrittura; Mursia: Milano, Italy, 2000. [Google Scholar]
- Mueller, P.A.; Oppenheimer, D.M. The pen is mightier than the keyboard: Advantages of longhand over laptop note taking. Psychol. Sci. 2014, 25, 1159–1168, Erratum in Psychol. Sci. 2018, 29, 1565–1568. https://doi.org/10.1177/0956797618781773. [Google Scholar] [CrossRef] [PubMed]
- Laganaro, F.; Mazza, M.; Marano, G.; Piuzzi, E.; Pallotti, A. Classification-Based Screening of Depressive Disorder Patients Through Graph, Handwriting and Voice Signals. In Proceedings of the 2023 International Workshop on Biomedical Applications, Technologies and Sensors (BATS), Catanzaro, Italy, 28–29 September 2023; pp. 6–10. [Google Scholar]
- Laganaro, F.; Mazza, M.; Marano, G.; Piuzzi, E.; Pallotti, A. Grapho-Phonological Signals-Based Machine Learning Models Development for the Depressive Disorder. In Annual Meeting of the Italian Electronics Society; Lecture Notes in Electrical Engineering; Springer Nature: Cham, Switzerland, 2023; pp. 436–454. [Google Scholar]
- Mancini, A.; Albani, G.; Marano, G.; Calabrese, R.; Veneziano, G.; Paffi, A.; Angelucci, M.; Mazza, M.; Pallotti, A. Graph and Handwriting Signals-Based Machine Learning Models Development in Parkinson’s Screening and Telemonitoring. In Proceedings of the 2023 IEEE International Workshop on Metrology for Industry 4.0 & IoT (MetroInd4.0&IoT), Brescia, Italy, 6–8 June 2023; pp. 183–188. [Google Scholar]
- Fratello, M.; Cordella, F.; Albani, G.; Veneziano, G.; Marano, G.; Paffi, A.; Pallotti, A. Classification-Based Screening of Parkinson’s Disease Patients through Graph and Handwriting Signals. Eng. Proc. 2021, 11, 49. [Google Scholar] [CrossRef]
- Prattichizzo, D.; Meli, L.; Malvezzi, M. Digital Handwriting with a Finger or a Stylus: A Biomechanical Comparison. IEEE Trans. Haptics 2015, 8, 356–370. [Google Scholar] [CrossRef] [PubMed]
- Pinet, S.; Dubarry, A.S.; Alario, F.X. Response retrieval and motor planning during typing. Brain Lang. 2016, 159, 74–83. [Google Scholar] [CrossRef]
- Maiero, J.; Eibich, D.; Kruijff, E.; Hinkenjann, A.; Stuerzlinger, W.; Benko, H.; Ghinea, G. Back-of-Device Force Feedback Improves Touchscreen Interaction for Mobile Devices. IEEE Trans. Haptics 2019, 12, 483–496. [Google Scholar] [CrossRef] [PubMed]
- Ihara, A.S.; Nakajima, K.; Kake, A.; Ishimaru, K.; Osugi, K.; Naruse, Y. Advantage of Handwriting over Typing on Learning Words: Evidence from an N400 Event-Related Potential Index. Front. Hum. Neurosci. 2021, 15, 679191. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Chen, D.; Song, A.; Tian, L.; Yu, Y.; Zhu, L. MH-Pen: A Pen-Type Multi-Mode Haptic Interface for Touch Screens Interaction. IEEE Trans. Haptics 2018, 11, 555–567. [Google Scholar] [CrossRef] [PubMed]
- Askvik, E.O.; van der Weel, F.R.R.; van der Meer, A.L.H. The importance of cursive handwriting over typewriting for learning in the classroom: A high-density EEG study of 12-year-old children and young adults. Front. Psychol. 2020, 11, 1810. [Google Scholar] [CrossRef]
- Beeson, P.M.; Rapcsak, S.Z.; Plante, E.; Chargualaf, J.; Chung, A.; Johnson, S.C.; Trouard, T.P. The neural substrates of writing: A functional magnetic resonance imaging study. Aphasiology 2003, 17, 647–665. [Google Scholar] [CrossRef]
- Roux, F.E.; Dufor, O.; Giussani, C.; Wamain, Y.; Draper, L.; Longcamp, M.; Démonet, J.F. The graphemic/motor frontal area Exner’s area revisited. Ann. Neurol. 2009, 66, 537–545. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, K.; Honda, M.; Okada, T.; Hanakawa, T.; Toma, K.; Fukuyama, H.; Konishi, J.; Shibasaki, H. Participation of the left posterior inferior temporal cortex in writing and mental recall of kanji orthography: A functional MRI study. Brain 2000, 123 Pt 5, 954–967. [Google Scholar] [CrossRef]
- Segal, E.; Petrides, M. The anterior superior parietal lobule and its interactions with language and motor areas during writing. Eur. J. Neurosci. 2012, 35, 309–322. [Google Scholar] [CrossRef]
- Longcamp, M.; Boucard, C.; Gilhodes, J.C.; Anton, J.L.; Roth, M.; Nazarian, B.; Velay, J.L. Learning through hand- or typewriting influences visual recognition of new graphic shapes: Behavioral and functional imaging evidence. J. Cogn. Neurosci. 2008, 20, 802–815. [Google Scholar] [CrossRef]
- Palmis, S.; Velay, J.L.; Fabiani, E.; Nazarian, B.; Anton, J.L.; Habib, M.; Kandel, S.; Longcamp, M. The impact of spelling regularity on handwriting production: A coupled fMRI and kinematics study. Cortex 2019, 113, 111–127. [Google Scholar] [CrossRef]
- James, K.H.; Engelhardt, L. The effects of handwriting experience on functional brain development in pre-literate children. Trends Neurosci. Educ. 2012, 1, 32–42. [Google Scholar] [CrossRef] [PubMed]
- Cisneros-Franco, J.M.; Voss, P.; Thomas, M.E.; de Villers-Sidani, E. Critical periods of brain development. Handb. Clin. Neurol. 2020, 173, 75–88. [Google Scholar] [CrossRef] [PubMed]
- Karimpoor, M.; Churchill, N.W.; Tam, F.; Fischer, C.E.; Schweizer, T.A.; Graham, S.J. Functional MRI of handwriting tasks: A study of healthy young adults interacting with a novel touch-sensitive tablet. Front. Hum. Neurosci. 2018, 12, 30. [Google Scholar] [CrossRef] [PubMed]
- Purcell, J.J.; Napoliello, E.M.; Eden, G.F. A combined fMRI study of typed spelling and reading. Neuroimage 2011, 55, 750–762. [Google Scholar] [CrossRef] [PubMed]
- Shah, C.; Erhard, K.; Ortheil, H.J.; Kaza, E.; Kessler, C.; Lotze, M. Neural correlates of creative writing: An fMRI study. Hum. Brain Mapp. 2013, 34, 1088–1101. [Google Scholar] [CrossRef] [PubMed]
- Petrides, M.; Alivisatos, B.; Evans, A.C. Functional activation of the human ventrolateral frontal cortex during mnemonic retrieval of verbal information. Proc. Natl. Acad. Sci. USA 1995, 92, 5803–5807. [Google Scholar] [CrossRef] [PubMed]
- Katanoda, K.; Yoshikawa, K.; Sugishita, M. A functional MRI study on the neural substrates for writing. Hum. Brain Mapp. 2001, 13, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Siebner, H.R.; Limmer, C.; Peinemann, A.; Bartenstein, P.; Drzezga, A.; Conrad, B. Brain correlates of fast and slow handwriting in humans: A PET-performance correlation analysis. Eur. J. Neurosci. 2001, 14, 726–736. [Google Scholar] [CrossRef]
- Barton, J.J.; Fox, C.J.; Sekunova, A.; Iaria, G. Encoding in the visual word form area: An fMRI adaptation study of words versus handwriting. J. Cogn. Neurosci. 2010, 22, 1649–1661. [Google Scholar] [CrossRef] [PubMed]
- Longcamp, M.; Lagarrigue, A.; Nazarian, B.; Roth, M.; Anton, J.L.; Alario, F.X.; Velay, J.L. Functional specificity in the motor system: Evidence from coupled fMRI and kinematic recordings during letter and digit writing. Hum. Brain Mapp. 2014, 35, 6077–6087. [Google Scholar] [CrossRef]
- Bartoň, M.; Fňašková, M.; Rektorová, I.; Mikl, M.; Mareček, R.; Rapcsak, S.Z.; Rektor, I. The role of the striatum in visuomotor integration during handwriting: An fMRI study. J. Neural Transm. 2020, 127, 331–337. [Google Scholar] [CrossRef]
- Higashiyama, Y.; Takeda, K.; Someya, Y.; Kuroiwa, Y.; Tanaka, F. The neural basis of typewriting: A functional MRI study. PLoS ONE 2015, 10, e0134131, Erratum in PLoS ONE 2015, 10, e0137265. https://doi.org/10.1371/journal.pone.0137265. [Google Scholar] [CrossRef]
- Van der Weel, F.R.R.; Van der Meer, A.L.H. Handwriting but not typewriting leads to widespread brain connectivity: A high-density EEG study with implications for the classroom. Front. Psychol. 2024, 14, 1219945. [Google Scholar] [CrossRef] [PubMed]
- Boyraz, R.K.; Kirpinar, I.; Yilmaz, O.; Özyurt, O.; Kiliçarslan, T.; Aralasmak, A. A treatment-response comparison study of resting-state functional magnetic resonance imaging between standard treatment of SSRI and standard treatment of SSRI plus non-dominant hand-writing task in patients with major depressive disorder. Front. Psychiatry 2021, 12, 698954. [Google Scholar] [CrossRef]
- Mangen, A.; Velay, J.L. Digitizing literacy:reflection on the haptic of wrinting. Adv. Haptics 2010, 1, 86–401. [Google Scholar]
- Suggate, S.P.; Karle, V.L.; Kipfelsberger, T.; Stoeger, H. The effect of fine motor skills, handwriting, and typing on reading development. J. Exp. Child Psychol. 2023, 232, 105674. [Google Scholar] [CrossRef] [PubMed]
- Natta, F. Corsivo vs computer. Perché scrivere a mano. Percorsi cognitivi e orizzonti di ricerca. In Studium Educationis; Pensa MultiMedia Editore: Lecce, Italy, 2016; Volume XVII. [Google Scholar]
- Tremblay, P.; Shiller, D.M.; Gracco, V.L. On the time-course and frequency selectivity of the EEG for different modes of response selection: Evidence from speech production and keyboard pressing. Clin. Neurophysiol. 2008, 119, 88–99. [Google Scholar] [CrossRef] [PubMed]
- Seitz, R.J.; Canavan, A.G.; Yágüez, L.; Herzog, H.; Tellmann, L.; Knorr, U.; Huang, Y.; Hömberg, V. Representations of graphomotor trajectories in the human parietal cortex: Evidence for controlled processing and automatic performance. Eur. J. Neurosci. 1997, 9, 378–389. [Google Scholar] [CrossRef]
- Matsuo, K.; Kato, C.; Tanaka, S.; Sugio, T.; Matsuzawa, M.; Inui, T.; Moriya, T.; Glover, G.H.; Nakai, T. Visual language and handwriting movement: Functional magnetic resonance imaging at 3 tesla during generation of ideographic characters. Brain Res. Bull. 2001, 55, 549–554. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, K.; Honda, M.; Hirano, S.; Oga, T.; Sawamoto, N.; Hanakawa, T.; Inoue, H.; Ito, J.; Matsuda, T.; Fukuyama, H.; et al. Modulation of the visual word retrieval system in writing: A functional MRI study on the Japanese orthographies. J. Cogn. Neurosci. 2002, 14, 104–115. [Google Scholar] [CrossRef]
- Omura, K.; Tsukamoto, T.; Kotani, Y.; Ohgami, Y.; Yoshikawa, K. Neural correlates of phoneme-to-grapheme conversion. Neuroreport 2004, 15, 949–953. [Google Scholar] [CrossRef] [PubMed]
- Rektor, I.; Rektorová, I.; Mikl, M.; Brázdil, M.; Krupa, P. An event-related fMRI study of self-paced alphabetically ordered writing of single letters. Exp. Brain Res. 2006, 173, 79–85, Erratum in Exp. Brain Res. 2006, 173, 193–194. [Google Scholar] [CrossRef]
- Harrington, G.S.; Farias, D.; Davis, C.H.; Buonocore, M.H. Comparison of the neural basis for imagined writing and drawing. Hum. Brain Mapp. 2007, 28, 450–459. [Google Scholar] [CrossRef] [PubMed]
- Brownsett, S.L.; Wise, R.J. The contribution of the parietal lobes to speaking and writing. Cereb. Cortex 2010, 20, 517–523. [Google Scholar] [CrossRef] [PubMed]
- Bisio, A.; Pedullà, L.; Bonzano, L.; Ruggeri, P.; Brichetto, G.; Bove, M. Evaluation of handwriting movement kinematics: From an ecological to a magnetic resonance environment. Front. Hum. Neurosci. 2016, 10, 488. [Google Scholar] [CrossRef]
- Longcamp, M.; Hlushchuk, Y.; Hari, R. What differs in visual recognition of handwritten vs. printed letters? An fMRI study. Hum. Brain Mapp. 2011, 32, 1250–1259. [Google Scholar] [CrossRef]
- Wamain, Y.; Tallet, J.; Zanone, P.G.; Longcamp, M. Brain responses to handwritten and printed letters differentially depend on the activation state of the primary motor cortex. Neuroimage 2012, 63, 1766–1773. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, H.; Nakajima, K.; Takagi, S.; Mizuyama, R.; Saito, M.; Furusawa, K.; Nakatani, K.; Yokota, Y.; Kataoka, H.; Nakajima, H.; et al. Differences in Mechanical Parameters of Keyboard Switches Modulate Motor Preparation: A Wearable EEG Study. Front. Neuroergonomics 2021, 2, 644449. [Google Scholar] [CrossRef]
- Smoker, T.J.; Murphy, C.E.; Rockwell, A.K. Comparing Memory for Handwriting versus Typing. Proc. Hum. Factors Ergon. Soc. Annu. Meet. 2009, 53, 1744–1747. [Google Scholar] [CrossRef]
- Mangen, A.; Anda, L.G.; Oxborough, G.H.; Brřnnick, K. Handwriting versus keyboard writing: Effect on word recall. J. Writ. Res. 2015, 7, 227–247. [Google Scholar] [CrossRef]
- Graham, S.; MacArthur, C.A.; Fitzgerald, J. Best Practices in Writing Instruction; Guilford Press: New York, NY, USA, 2013. [Google Scholar]
- Baron, N.S. Who Wrote This? How AI and the Lure of Efficiency Threaten Human Writing; Hardcover: Redwood City, CA, USA, 2023. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
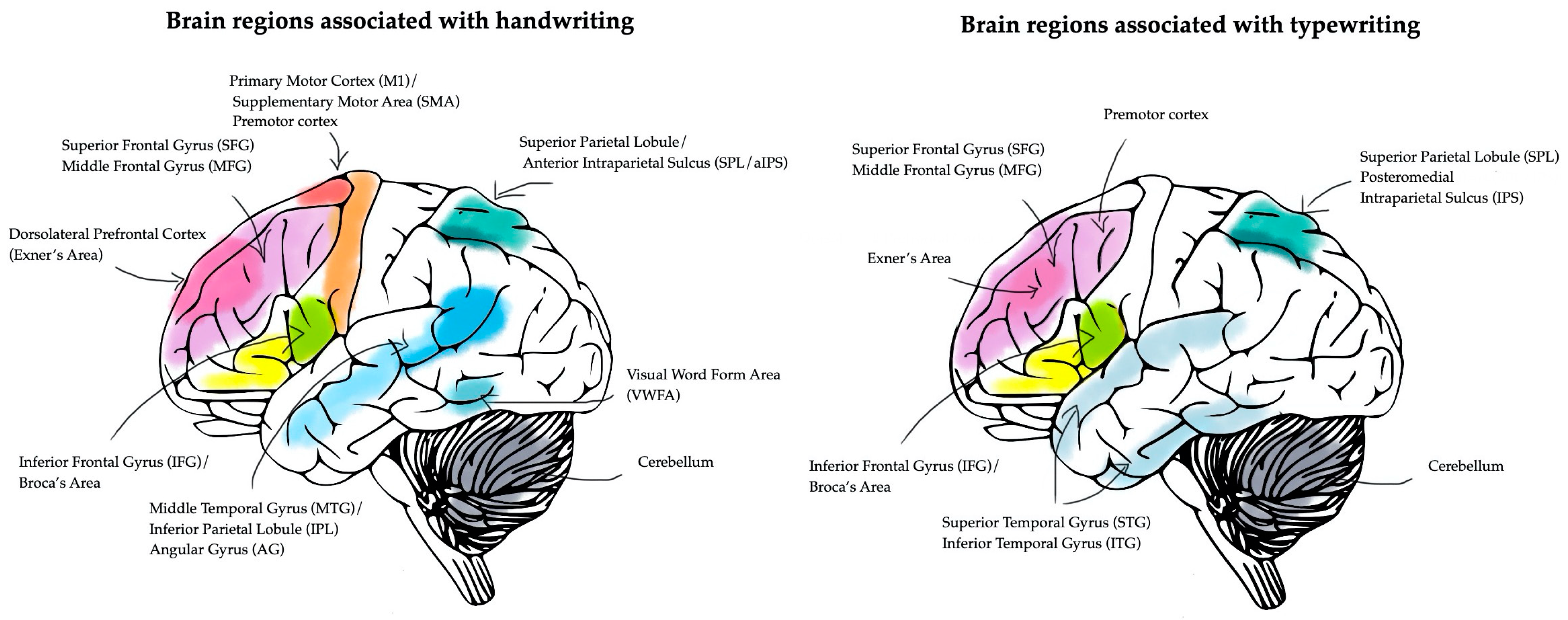
| Cerebral Area | Function |
|---|---|
| Visual word form area (VWFA) | Stores letter shapes and orthographic knowledge. |
| Superior parietal lobule, anterior intraparietal sulcus (SPL/aIPS) | Visuospatial integration, motor planning, and letter-specific motor programs. |
| Dorsolateral prefrontal cortex (Exner’s area) | Translates letter representations into motor commands. |
| Primary motor cortex (M1) and supplementary motor area (SMA) | Executes and coordinates motor commands. |
| Inferior frontal gyrus (IFG), Broca’s area | Phoneme-to-grapheme conversion, crucial in phonological agraphia. |
| Middle frontal gyrus (MFG) and superior frontal gyrus (SFG) | Higher-level motor control and selection of well-practiced graphic patterns. |
| Cuneus/precuneus | Visual processing and attention. |
| Middle temporal gyrus (MTG) and angular gyrus (AG) | Language processing and integration of phonological and orthographic representations. |
| Ventral precentral gyrus and inferior parietal lobule (IPL) | Specialized for writing movements. |
| Sensorimotor striatum (smStr) | Integrates visual letter forms with motor execution. |
| Basal ganglia (striatum, putamen, pallidum, caudate nucleus) | Fine-tunes writing kinematics, motor sequence learning, and inhibition of competing actions. |
| Thalamus | Relays motor and sensory information. |
| Cerebellum | Adjusts movement precision, coordination, and timing. |
| Cerebral Area | Function |
|---|---|
| Inferior frontal gyrus (IFG), Broca’s area | Supports phoneme-to-grapheme conversion, spelling, and lexical retrieval. |
| Middle frontal gyrus (MFG), superior frontal gyrus (SFG) | Involved in higher-level motor planning and executive functions related to typing. |
| Intraparietal sulcus (IPS), superior parietal lobule (SPL) | Engages spatial processing and attention control, supporting key-to-finger mappings in typewriting. |
| Inferior temporal gyrus (ITG), fusiform gyrus (FuG) | Plays a role in orthographic processing and letter recognition. |
| Superior temporal gyrus (STG) | Involved in auditory and phonological processing during spelling tasks. |
| Right supramarginal gyrus (BA 40) | Engaged in spatial learning and attentional shifts when typing characters. |
| Posteromedial intraparietal sulcus (IPS) | Specific to typing, linked to fine motor control. |
| Basal ganglia (striatum) | Plays a role in procedural learning and automation of keypress sequences. |
| Thalamus | Relays motor and sensory information, but with reduced engagement compared with handwriting. |
| Cerebellum | Contributes to motor learning and coordination of typing movements. |
| Study | Population | Technique | Conclusion/Observations |
|---|---|---|---|
| Petrides et al., 1995 [29] | 11 ♂ r-h prtcs, = 24, age range 18–32 yrs. | PET, H215° rCBF, 1.5T MRI co-registration on T&TSA. | VLPFC is involved in the strategic retrieval of verbal episodic information, while the DLPFC is activated during tasks requiring mnemonic output monitoring, and the SMCx and posterior PCx are activated during writing, suggesting distinct functional specializations within the frontal Cx. |
| Katanoda et al., 2001 [30] | 17 r-h ♂ healthy prtcs, age range 19–31 yrs, with normal or corrected-to-normal vision. | fMRI, 1.5T, T2*-weighted BOLD aligned with T1-weighted scans for anatomic localization. | The results differentiate writing from other linguistic and motor activities. |
| Siebner et al., 2001 [31] | 11 healthy r-h prtcs (2 ♀, 9 ♂; age 42 yrs, range 26–58 yrs) vs. 11 healthy l-h converted to right handwriting (4 ♀, 7 ♂; age 47 yrs, range 34–64 yrs) vs. controls, 6 consistent l-h, not converted to right hand writing (1 ♀, 5 ♂; age 32 yrs, range 25–55 yrs). | PET, H215°-rCBF; analysis through SPM implemented in PRO-MATLAB environment normalized on MNI template; location of peak activation with T&TSA. | Writing elicited activation patterns in consistent r-h prtcs in the dominant (contralateral) hemisphere. Stronger activation of the right lateral PFC in converted l-h compared with more prominent activation in the left dorsal and ventral PFC during right-hand writing in r-h might reflect adaptation, i.e., more activation is needed to support a “corrected” behavior. |
| Beeson et al., 2003 [18] | 12 healthy r-h adults (6 ♀; 6 ♂) age range 20- 53 yrs, age = 38.4 yrs, English speakers. | fMRI, 1.5T, T2*-weighted BOLD aligned with T1-weighted scans to localize areas according to MNI-SPM99. | This study contradicts the long-proposed notion that orthographic representations for familiar words are stored in the dominant (left) AG (BA39) in the form of visual images. |
| Roux et al., 2009 [19] | 12 > 63 ± 19.18% r-h ( age 22 ± 3.88 yrs and 12 > 69 ± 18.91% l-h healthy volunteers ( age 25 ± 5.05). Sex of volunteers not specified. | fMRI 1.5T; EPI sequenced images normalized on T&TSA template and analyzed with MATLAB 7 using SPM5. | An area located close to the superior frontal sulcus in BA6, anterior to the hand primary motor area, is selectively involved in handwritten word production. Handwriting-related activation in BA6 was bilateral in r-h prtcs but slightly ↑ in the dominant hemisphere. |
| Barton et al., 2010 [32] | 13 healthy r-h English-literate individuals (5 ♀, 8 ♂; age 28.1 ± 4.0 yrs, range 22–35 yrs) with normal or corrected-to-normal visual acuity. | fMRI 3T, T2-weighted functional images with EPI sequences; ROI approach combined with whole-brain analysis; images plotted on T&TSA. | Abstract word representations might emerge at higher processing levels. The VWFA is primarily involved in perceptual text analysis, particularly handwriting, rather than abstract linguistic processing and is sensitive to handwriting, supporting a primary role in perceptual rather than linguistic processing. Findings challenge the notion of the VWFA as exclusively dedicated to linguistic functions, emphasizing its contribution to visual and stylistic text analysis. |
| Segal & Petrides, 2012 [21] | Nine healthy r-h bilingual speakers (English–French or German or Chinese, six ♂, three ♀; age 26 ±3.72 yrs). | fMRI, 1.5T, T2*-weighted BOLD aligned with T1-weighted scans to map according to the MNI co-ordinates. | PE area in the SPL of the language-dominant hemisphere is a critical high-level motor control region for writing. This area acts as an interface between cortical motor and language regions, coordinating hand actions required for producing written language. |
| Longcamp et al., 2014 [33] | 18 native French speakers (11 ♀, 7 ♂; age 24 yrs, range 18–35 yrs), with normal audition and normal or corrected-to-normal vision. | fMRI 3T, T1-weighted MP-RAGE sequence superimposed on T2*-weighted EPI, analyzed with SPM8 on MNI template. | Observed differences between letters and digits were not related to low-level kinematic features. Writing engages an extended motor-perceptual network. Only two very restricted areas discriminate between letters and numbers. The dorsal preMCx is a fundamental brain network node for handwriting. |
| Planton et al., 2017 [5] | 16 healthy r-h native French (8 ♂, 8 ♀; age 25.3 ± 6.0 yrs). | fMRI 3T EPI sequences normalized and realigned on MNI template and analyzed through SPM12b; VOI approach | The ventral part of the precentral gyrus and the left IPL show a preference for writing, suggesting specialization. Writing and oral spelling overlap with traditional language areas like left IFG (Broca’s area) and STG. Writing activates additional motor and visuospatial regions not involved in oral spelling. Results support a hierarchical model in which regions specific to writing overlap with more general circuits used for motor control and language. Findings are relevant to understanding writing disorders (e.g., dysgraphia) and designing therapeutic interventions. |
| Karimpour et al., 2018 [26] | 12 r-h healthy Canadian adult graduates (5 ♂, 7 ♀; age 25). | fMRI 3T T2*-weighted BOLD analyzed with SPM normalized on MNI template. | Behavioral results confirm that IAT metrics successfully discriminate performance differences across tasks. VPMc regions have a role in guiding hand movements. Variations in brain activity across the three tasks reflected differing cognitive processing demands. |
| Bartoň et al., 2020[34] | 20 r-h healthy volunteers (9 ♀, 11 ♂; age 23 ± 2.4 yrs) | fMRI 1.5T BOLD, EPI sequencing T1-weighted MRI analysed through SPM12 with normalization on the MNI template. ROI approach | The striatum plays a crucial role in integrating stored letter-shape representations with motor planning and execution processes during handwriting. A writing-specific cortico-striatal network comprises the VWFA, SPL/anterior IPS, striatum, preMCx/Exner’s area, primary motor cortex, and SMA. The striatum is important for integrating information about letter shapes, selecting/retrieving abstract motor programs, and generating concrete motor commands |
| Study | Population | Technique | Conclusion/Observations |
|---|---|---|---|
| Purcell et al., 2011 [27] | 17 healthy r-h English speakers (7 ♂, 10 ♀; age 23.2 yrs, range 18–27 yrs). | fMRI, 3T; EPI sequences plus T1-weighted scans; images analyzed with SPM5 and normalized according to MNI. | Typewriting activates a spelling network that is active conjointly with reading. The typewriting spelling includes a region near Exner’s area, which is shared by handwriting. Lateral IFG activation associated with spelling was ˃ reading, while in the medial portion there was no significant difference spelling vs. reading. |
| Higashiyama et al., 2015 [35] | 16 healthy r-h native Japanese speakers, skillful typists (9 ♂, 7 ♀; age 27.8 ± 3.1 yrs, range 23–34 yrs). | fMRI 3T, T1-weighted images MP-RAGE sequences analyzed voxel-wise with FWE in MNI space using SPM8 implemented in MATLAB 7.7.0. | The brain regions known as the “writing centre” are the same for typing. Scan comparisons of typing and writing showed the left posteromedial IPS to be typing-specific. Activity in the left MFG/SFG was more rostral in the typing than in the writing task, pointing to differences in motor planning. |
| Askvik et al., 2020 [17] | 12 r-h healthy school-aged children (four ♂, eight ♀; age 11.83 ± 0.39 yrs); 12 r-h healthy young adults (six ♂, six ♀; age 23.58 ± 2.02 yrs). | hd EEG, 256 channel with electrodes distributed over scalp. | Handwriting and drawing are associated with memory and learning processes; there is different cognitive engagement between handwriting and drawing vs. typewriting. Findings underscore the importance of handwriting and drawing activities in early childhood education. Sensorimotor integration and fine motor skills involved in these activities contribute to the development of neural networks essential for learning and cognitive development. |
| Van der Weel and Van der Meer, 2024 [36] | 36 r-h prtcs in their early twenties; sex not specified. | hd EEG, 256 channel with electrodes distributed over scalp. | ↑ FC between the various brain regions seems to be linked to the specific sensorimotor processes that are typical in handwriting. Spatial temporal patterns from visual and proprioceptive information during handwriting movements have a beneficial impact on the brain’s FC patterns related to learning and remembering. |
| Study | Population | Technique | Conclusion/Observations |
|---|---|---|---|
| Seitz et al., 1997 [42] | Eight healthy r-h Caucasian medical students (four ♂, four ♀, age 28 ± 3 yrs) with normal or corrected visual acuity. | PET, [15O]butanol rCBF, 1.5T MRI co-registration on T&TSA. | The PCx engages in motor coordination and learning of new movement patterns associated with the automatic processing of previously consolidated movements. There are two subsystems, one engaged in controlled processing around the anterior IPS and another in posterior PCx during automatic execution. During mental imagery, both subsystems are simultaneously active. |
| Nakamura et al., 2000 [20] | 10 healthy r-h students at Kyoto University, native Japanese speakers (6 ♂, 4 ♀, age range 20–25 yrs). | fMRI, 1.5T images analyzed through SPM96. | Evidence suggests that the left PITCx plays a central role in the retrieval of visual engrams of letters. During the act of writing, there is no need for conscious effort to recall the visual images of letters or words to be written, as the sequential visuomotor skill proceeds automatically due to the presence of a specialized neural sub-system for retrieving visual graphic forms. |
| Matsuo et al., 2001 [43] | 12 r-h (1 ambidextrous) healthy Japanese (8 ♂, 4 ♀, age range 22–29 yrs). | fMRI 3T, T2-weighted images overlapping with fMRI; ROI analysis with SPM96 using T&TSA MR normalization. | Activation in higher motor areas may be induced by cognitive components related to motor function when performing visuospatial language tasks. |
| Siebner et al., 2001 [31] | 10 healthy r-h prtcs (8 ♂, 2 ♀; age 41.3 ± 10.9 yrs). | PET, H215O-rCBF normalized on T&TSA MRs | A set of cortical and subcortical brain regions is involved in the processing of movements associated with SC-L writing. FO-L handwriting is achieved by optimized cooperation of the manual sensorimotor network rather than by selective activation of a distinct network component. The functional activation pattern during handwriting is not influenced by different levels of motor learning. |
| Nakamura et al., 2002 [44] | Nine healthy r-h volunteers (age range 21–30 yrs), native Japanese speakers. | fMRI, 1.5T images analyzed through SPM96. | Contrary to the traditional view, the obtained functional imaging evidence indicated that in normal people, the neural systems for processing the two different scripts are separable only functionally, rather than anatomically. |
| Omura et al., 2004 [45] | 15 healthy r-h adults (9 ♂, 6 ♀), age range 18–39 yrs ( age 25.7 ± 6.4), Japanese speakers. | fMRI, 1.5T, T2*-weighted BOLD aligned with T1-weighted; areas defined with MNI-SPM99. | Findings suggest that the left premotor area transfers phonemic representations to graphemic motor output to create letters in writing to dictation. The left STG is thought to transform auditory information into phonetic (auditory) and/or graphic (visual) representations of letters. |
| Rektor et al., 2006 [46] | 10 healthy r-h adult Czech speakers (2 ♀, 8 ♂) age range 20–25 yrs, age 23.5 ± 1.28. | fMRI, 1.5T, T1-weighted scans fitted on fMRI images to localize areas using SPM99. | The right hemisphere is dominant for tasks requiring manipulation in space. It is possible that the activation of this region is linked with the spatial dimension of the writing. Right-sided PCx may play an important role in the elemental mechanism of writing. |
| Harrington et al., 2007 [47] | 11 healthy r-h adults (5 ♂, 6 ♀); age 37.1 yrs, range 22–61 yrs, of whom 6 (2 ♂, 4 ♀; age 39.0 yrs, range 26–54 yrs) participated in Experiment 2. | fMRI 1.5T, T1-weighted images EPI sequences analyzed through AFNI AlphaSim normalized on T&TSA; ROI approach. | Mental imagery of motion recruits the same Cx areas used in motor performance. The differences in activation reflect the use of distinct brain pathways for symbolic (writing) vs. visuo-spatial (drawing) processes. |
| Longcamp et al., 2008 [22] | 12 r-h prtcs (6 ♂, 6 ♀; age 26 ± 3 yrs). | fMRI 3T, T2*-weighted EPI sequence superimposed on T1-weighted images; analysis with SPM2 normalized on MNI template; significant clusters identified with T&TSA. | Better recognition of new characters when written compared with typed. Handwriting memory facilitated discrimination between characters and their mirror images for longer periods than typewriting memory. Via fMRI, different neural circuits were found to underlie differences in recognition performance between hand- and typewritten characters, supported by left-sided lateralization of activations. |
| Tremblay et al., 2008 [41] | ST: Six healthy r-h English speakers (four ♀, two ♂; age 26 ± 4.91 yrs). NST: Six healthy r-h English speakers (three ♀, three ♂; age 22.7 ± 2.27 yrs). | EEG 64-pin active Ag-AgCl electrodes, EMG of the orbicularis oris muscle with two flat electrodes placed above upper and lower lip. | β ERD patterns showed a closer relation to response selection. In contrast, α ERD patterns were more closely related to response onset, perhaps reflecting attentional demands overseeing execuction of motor response. ERD patterns for the speech and keyboard-pressing tasks were very similar across the frequency bands, suggesting that the orofacial and finger motor systems used similar underlying neural mechanisms. |
| Brownsett & Wise, 2010 [48] | Three healthy r-h (seven ♂, six ♀; age 51.7 yrs, range 40–70 yrs). | PET, H215O rCBF; images analyzed with SPM2. | Extensive parietal activity is associated with the planning, execution, and monitoring of writing, even for only a single letter. The PL contributes little to the planning, execution, and monitoring of articulation during normal spoken language production. Parietal activity for amodal linguistic or mnemonic processing is confined to the left AG. |
| Shah et al., 2013 [28] | 28 r-h prtcs (14 ♀, 14 ♂; age 24.0 ± 1.9 yrs) in-experienced in creative writing, native German speakers. | fMRI 3T, BOLD, T1-weighted, EPI, images analysed using SPM5 run on MATLAB 7.4 normalized to MNI. | Creative writing and copying shared many brain activations and correlated with creativity indexes; hence, copying is not devoid of creative processes and their related brain activations, mostly the SFG–temporopolar focus. Brainstorming involved novel and original idea generation and composition of the story concept through recruiting fronto-parieto-temporal circuits. Some PreMCx activity may have been involved in preparatory processes for writing activity. Creative writing combines handwriting with cognitive processes associated with writing, such as episodic memory, semantic integration, and text production, linked to activation of a left fronto-temporal network including left IFG (BA45) and left temporopolar Cx (BA38). |
| Bisio et al., 2016 [49] | Enrolled 44 r-h prtcs, 22 (12 ♀, 10 ♂; age 25 ± 5.6 years) in Ex-periment 1, 22 (14 ♀, 8 ♂; age 24.2 ± 6.1 years) in Experiment 2, of whom 7 underwent fMRI scanning (sex and age not specified). | fMRI 1.5T, T2*-weighted sin-gle-shot EPI sequences normal-ised on MNI template analysed with SPM12. | The tablet reliably captured kinematic data across time, including fairly predictably under constrained MR conditions. fMRI identified activations in brain regions purportedly associated with handwriting in healthy people. |
| Palmis et al., 2019 [23] | 25 r-h native French speakers ( age 24, range 19–37); sex not specified. | fMRI 3T, T2* BOLD and T1 MR superimposition analyzed through SPM12. ROI. | Core regions of the written language network, like left IFG and FuG, are sensitive to orthographic irregularities during writing. Motor-related regions, like SFG and SPL, exhibited ↑ activation for irregular words. Findings support dynamic interactive and parallel orthographic and motor processes during handwriting. |
| Boyraz et al., 2021 [37] | 26 r-h ♀ with MDD, age range 18–50 yrs. | rs-fMRI 1.5T, T1-weighted MP-RAGE sequence for ROI analysis with CONN software using MATLAB-based SPM. | Adding NHE to standard treatment may be associated with modest improvements in symptoms of depression, despite significant differences between NHE and non-NHE in their FC changes. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marano, G.; Kotzalidis, G.D.; Lisci, F.M.; Anesini, M.B.; Rossi, S.; Barbonetti, S.; Cangini, A.; Ronsisvalle, A.; Artuso, L.; Falsini, C.; et al. The Neuroscience Behind Writing: Handwriting vs. Typing—Who Wins the Battle? Life 2025, 15, 345. https://doi.org/10.3390/life15030345
Marano G, Kotzalidis GD, Lisci FM, Anesini MB, Rossi S, Barbonetti S, Cangini A, Ronsisvalle A, Artuso L, Falsini C, et al. The Neuroscience Behind Writing: Handwriting vs. Typing—Who Wins the Battle? Life. 2025; 15(3):345. https://doi.org/10.3390/life15030345
Chicago/Turabian StyleMarano, Giuseppe, Georgios D. Kotzalidis, Francesco Maria Lisci, Maria Benedetta Anesini, Sara Rossi, Sara Barbonetti, Andrea Cangini, Alice Ronsisvalle, Laura Artuso, Cecilia Falsini, and et al. 2025. "The Neuroscience Behind Writing: Handwriting vs. Typing—Who Wins the Battle?" Life 15, no. 3: 345. https://doi.org/10.3390/life15030345
APA StyleMarano, G., Kotzalidis, G. D., Lisci, F. M., Anesini, M. B., Rossi, S., Barbonetti, S., Cangini, A., Ronsisvalle, A., Artuso, L., Falsini, C., Caso, R., Mandracchia, G., Brisi, C., Traversi, G., Mazza, O., Pola, R., Sani, G., Mercuri, E. M., Gaetani, E., & Mazza, M. (2025). The Neuroscience Behind Writing: Handwriting vs. Typing—Who Wins the Battle? Life, 15(3), 345. https://doi.org/10.3390/life15030345