
Mycobacterium avium subsp. paratuberculosis Antigens Elicit a Strong IgG4 Response in Patients with Multiple Sclerosis and Exacerbate Experimental Autoimmune Encephalomyelitis

 ,
,  ,
,  and
and
Abstract
1. Introduction
- To evaluate the humoral response against the peptide MAP_HSP70533–545 in Japanese and Italian patients with RR-MS through in vitro experiments.
- To characterize the clinical parameters associated with the IgG subclass response to MAP_HSP70533–545.
- To assess the cross-reactivity between EBNA1386–405 and MAP_HSP70533–545 peptides.
- To evaluate the impact of MAP_HSP70533–545 on neuroinflammation using an active EAE model.
2. Materials and Methods
2.1. Patients
2.2. Antigens
2.3. Peptide-Based Enzyme-Linked Immunosorbent Assays (ELISAs)
2.4. Indirect ELISA for Anti-Lipopentapeptide (MAP_Lp5) Antibodies
2.5. Inhibition ELISA
2.6. Animal and Mouse Immunization
2.7. Histological Analysis and T-Cell Proliferation Assay
2.8. Statistics
3. Results
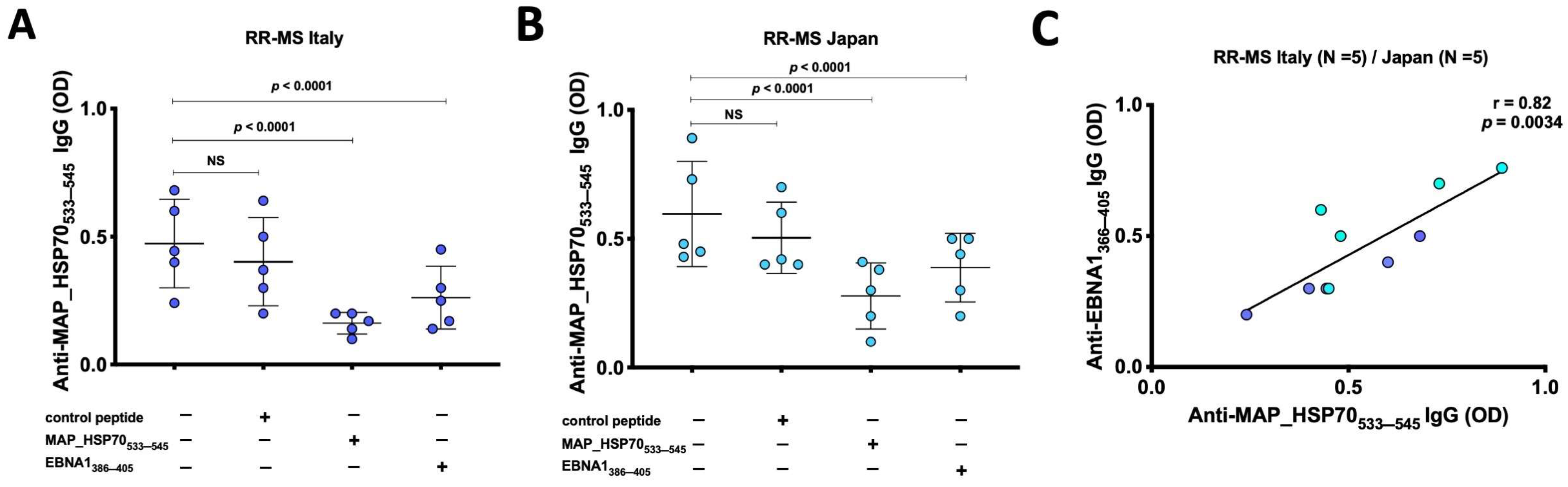
3.1. Anti-MAP_HSP70533–545 Antibodies Are Prevalent in Japanese and Italian RR-MS Patients
3.2. Cross Reactivity between HSP70 and EBNA
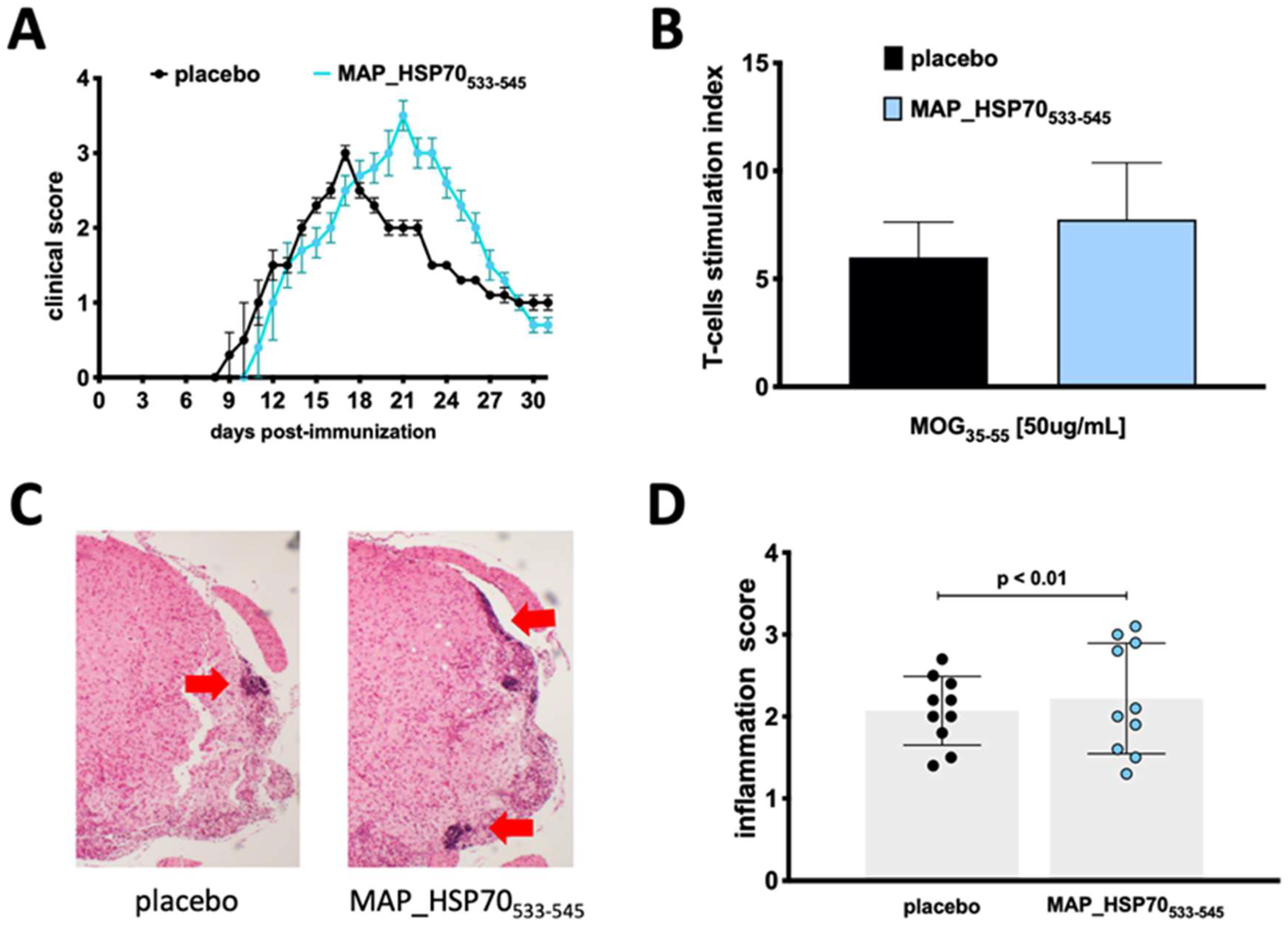
3.3. MAP_HSP70533–545 Immunization Exacerbates Active EAE
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cossu, D.; Yokoyama, K.; Hattori, N. Bacteria-Host Interactions in Multiple Sclerosis. Front. Microbiol. 2018, 9, 2966. [Google Scholar] [CrossRef] [PubMed]
- Goris, A.; Vandebergh, M.; McCauley, J.L.; Saarela, J.; Cotsapas, C. Genetics of multiple sclerosis: Lessons from polygenicity. Lancet Neurol. 2022, 21, 830–842. [Google Scholar] [CrossRef] [PubMed]
- Wasko, N.J.; Nichols, F.; Clark, R.B. Multiple sclerosis, the microbiome, TLR2, and the hygiene hypothesis. Autoimmun. Rev. 2020, 19, 102430. [Google Scholar] [CrossRef] [PubMed]
- Rousseau, B.A.; Bhaduri-McIntosh, S. Inflammation and Epstein-Barr Virus at the Crossroads of Multiple Sclerosis and Post-Acute Sequelae of COVID-19 Infection. Viruses 2023, 15, 949. [Google Scholar] [CrossRef]
- Meier, U.C.; Cipian, R.C.; Karimi, A.; Ramasamy, R.; Middeldorp, J.M. Cumulative Roles for Epstein-Barr Virus, Human Endogenous Retroviruses, and Human Herpes Virus-6 in Driving an Inflammatory Cascade Underlying MS Pathogenesis. Front. Immunol. 2021, 12, 757302. [Google Scholar] [CrossRef] [PubMed]
- Cossu, D.; Yokoyama, K.; Tomizawa, Y.; Momotani, E.; Hattori, N. Altered humoral immunity to mycobacterial antigens in Japanese patients affected by inflammatory demyelinating diseases of the central nervous system. Sci. Rep. 2017, 7, 3179. [Google Scholar] [CrossRef] [PubMed]
- Cossu, D.; Yokoyama, K.; Sechi, L.A.; Otsubo, S.; Tomizawa, Y.; Momotani, E.; Hattori, N. Humoral response against host-mimetic homologous epitopes of Mycobacterium avium subsp. paratuberculosis in Japanese multiple sclerosis patients. Sci. Rep. 2016, 6, 29227. [Google Scholar] [CrossRef]
- Hayashi, F.; Isobe, N.; Cossu, D.; Yokoyama, K.; Sakoda, A.; Matsushita, T.; Hattori, N.; Kira, J.I. Elevated mycobacterium avium subsp. paratuberculosis (MAP) antibody titer in Japanese multiple sclerosis. J. Neuroimmunol. 2021, 360, 577701. [Google Scholar] [CrossRef]
- Cossu, D.; Yokoyama, K.; Hattori, N. Conflicting Role of Mycobacterium Species in Multiple Sclerosis. Front. Neurol. 2017, 8, 216. [Google Scholar] [CrossRef]
- Kuenstner, L.; Kuenstner, J.T. Mycobacterium avium ssp. paratuberculosis in the Food Supply: A Public Health Issue. Front. Public Health 2021, 9, 647448. [Google Scholar] [CrossRef]
- Cossu, D.; Yokoyama, K.; Sakanishi, T.; Momotani, E.; Hattori, N. Adjuvant and antigenic properties of Mycobacterium avium subsp. paratuberculosis on experimental autoimmune encephalomyelitis. J. Neuroimmunol. 2019, 330, 174–177. [Google Scholar] [CrossRef]
- Cossu, D.; Yokoyama, K.; Sakanishi, T.; Kuwahara-Arai, K.; Momotani, E.; Hattori, N. A mucosal immune response induced by oral administration of heat-killed Mycobacterium avium subsp. paratuberculosis exacerbates EAE. J. Neuroimmunol. 2021, 352, 577477. [Google Scholar] [CrossRef]
- Mameli, G.; Cossu, D.; Cocco, E.; Masala, S.; Frau, J.; Marrosu, M.G.; Sechi, L.A. Epstein-Barr virus and Mycobacterium avium subsp. paratuberculosis peptides are cross recognized by anti-myelin basic protein antibodies in multiple sclerosis patients. J. Neuroimmunol. 2014, 270, 51–55. [Google Scholar] [CrossRef]
- Mameli, G.; Cocco, E.; Frau, J.; Marrosu, M.G.; Sechi, L.A. Epstein Barr Virus and Mycobacterium avium subsp. paratuberculosis peptides are recognized in sera and cerebrospinal fluid of MS patients. Sci. Rep. 2016, 6, 22401. [Google Scholar] [CrossRef]
- Lanz, T.V.; Brewer, R.C.; Ho, P.P.; Moon, J.S.; Jude, K.M.; Fernandez, D.; Fernandes, R.A.; Gomez, A.M.; Nadj, G.S.; Bartley, C.M.; et al. Clonally expanded B cells in multiple sclerosis bind EBV EBNA1 and GlialCAM. Nature 2022, 603, 321–327. [Google Scholar] [CrossRef]
- Mameli, G.; Madeddu, G.; Cossu, D.; Galleri, G.; Manetti, R.; Babudieri, S.; Mura, M.S.; Sechi, L.A. Immune response induced by Epstein-Barr virus and Mycobacterium avium subsp. paratuberculosis peptides in current and past infectious mononucleosis: A risk for multiple sclerosis? Eur. J. Neurol. 2016, 23, 140–147. [Google Scholar] [CrossRef]
- Cossu, D.; Masala, S.; Frau, J.; Cocco, E.; Marrosu, M.G.; Sechi, L.A. Anti Mycobacterium avium subsp. paratuberculosis heat shock protein 70 antibodies in the sera of Sardinian patients with multiple sclerosis. J. Neurol. Sci. 2013, 335, 131–133. [Google Scholar] [CrossRef] [PubMed]
- Thompson, A.J.; Banwell, B.L.; Barkhof, F.; Carroll, W.M.; Coetzee, T.; Comi, G.; Correale, J.; Fazekas, F.; Filippi, M.; Freedman, M.S.; et al. Diagnosis of multiple sclerosis: 2017 revisions of the McDonald criteria. Lancet Neurol. 2018, 17, 162–173. [Google Scholar] [CrossRef]
- Yokoyama, K.; Cossu, D.; Hoshino, Y.; Tomizawa, Y.; Momotani, E.; Hattori, N. Anti-Mycobacterial Antibodies in Paired Cerebrospinal Fluid and Serum Samples from Japanese Patients with Multiple Sclerosis or Neuromyelitis Optica Spectrum Disorder. J. Clin. Med. 2018, 7, 522. [Google Scholar] [CrossRef] [PubMed]
- Otsubo, S.; Cossu, D.; Eda, S.; Otsubo, Y.; Sechi, L.A.; Suzuki, T.; Iwao, Y.; Yamamoto, S.; Kuribayashi, T.; Momotani, E. Seroprevalence of IgG1 and IgG4 class antibodies against Mycobacterium avium subsp. paratuberculosis in Japanese population. Foodborne Pathog. Dis. 2015, 12, 851–856. [Google Scholar] [CrossRef] [PubMed]
- Cossu, D.; Yokoyama, K.; Sakanishi, T.; Sechi, L.A.; Hattori, N. Bacillus Calmette-Guerin Tokyo-172 vaccine provides age-related neuroprotection in actively induced and spontaneous experimental autoimmune encephalomyelitis models. Clin. Exp. Immunol. 2023, 212, 70–80. [Google Scholar] [CrossRef] [PubMed]
- Chiricosta, L.; Gugliandolo, A.; Bramanti, P.; Mazzon, E. Could the Heat Shock Proteins 70 Family Members Exacerbate the Immune Response in Multiple Sclerosis? An in Silico Study. Genes 2020, 11, 615. [Google Scholar] [CrossRef]
- Morales, J.E.; De, A.; Miller, A.A.; Chen, Z.; McCarty, J.H. Mlc1-Expressing Perivascular Astrocytes Promote Blood-Brain Barrier Integrity. J. Neurosci. 2022, 42, 1406–1416. [Google Scholar] [CrossRef]
- Arnedo, T.; Aiello, C.; Jeworutzki, E.; Dentici, M.L.; Uziel, G.; Simonati, A.; Pusch, M.; Bertini, E.; Estevez, R. Expanding the spectrum of megalencephalic leukoencephalopathy with subcortical cysts in two patients with GLIALCAM mutations. Neurogenetics 2014, 15, 41–48. [Google Scholar] [CrossRef]
- Pilzecker, B.; Jacobs, H. Mutating for Good: DNA Damage Responses During Somatic Hypermutation. Front. Immunol. 2019, 10, 438. [Google Scholar] [CrossRef] [PubMed]
- Beltran, E.; Obermeier, B.; Moser, M.; Coret, F.; Simo-Castello, M.; Bosca, I.; Perez-Miralles, F.; Villar, L.M.; Senel, M.; Tumani, H.; et al. Intrathecal somatic hypermutation of IgM in multiple sclerosis and neuroinflammation. Brain 2014, 137, 2703–2714. [Google Scholar] [CrossRef]
- Volodarsky, I.; Anton, A.; Zilberman, L.; Fugenfirov, I.; Neumark, E.; Malnick, S.; Levy, Y.; George, J.; Goland, S. Elevated Serum Levels of IgG4 in Patients with Heart Failure with Reduced Ejection Fraction: A Prospective Controlled Study. Biology 2022, 11, 1168. [Google Scholar] [CrossRef]
- Frau, J.; Coghe, G.; Lorefice, L.; Fenu, G.; Cocco, E. Infections and Multiple Sclerosis: From the World to Sardinia, From Sardinia to the World. Front. Immunol. 2021, 12, 728677. [Google Scholar] [CrossRef]
- Sechi, L.A.; Dow, C.T. Mycobacterium avium ss. paratuberculosis Zoonosis—The Hundred Year War—Beyond Crohn’s Disease. Front. Immunol. 2015, 6, 96. [Google Scholar] [CrossRef]
- Momotani, E. Epidemiological situation and control strategies for paratuberculosis in Japan. Jpn. J. Vet. Res. 2012, 60, S19–S29. [Google Scholar] [PubMed]
- Cossu, D.; Otsubo, S.; Otsubo, Y.; Eda, S.; Suzuki, T.; Iwao, Y.; Kuribayashi, T.; Yamamoto, S.; Sechi, L.A.; Momotani, E. Mycobacterium avium Subsp. paratuberculosis Induces Specific IgE Production in Japanese People with Allergies. Int. J. Inflam. 2017, 2017, 7959154. [Google Scholar] [CrossRef]
- Kumar, A.; Cocco, E.; Atzori, L.; Marrosu, M.G.; Pieroni, E. Structural and dynamical insights on HLA-DR2 complexes that confer susceptibility to multiple sclerosis in Sardinia: A molecular dynamics simulation study. PLoS ONE 2013, 8, e59711. [Google Scholar] [CrossRef] [PubMed]
- Yoshimura, S.; Isobe, N.; Matsushita, T.; Masaki, K.; Sato, S.; Kawano, Y.; Ochi, H.; Kira, J. Genetic and infectious profiles influence cerebrospinal fluid IgG abnormality in Japanese multiple sclerosis patients. PLoS ONE 2014, 9, e95367. [Google Scholar] [CrossRef] [PubMed]
- Kawa, S.; Ota, M.; Yoshizawa, K.; Horiuchi, A.; Hamano, H.; Ochi, Y.; Nakayama, K.; Tokutake, Y.; Katsuyama, Y.; Saito, S.; et al. HLA DRB10405-DQB10401 haplotype is associated with autoimmune pancreatitis in the Japanese population. Gastroenterology 2002, 122, 1264–1269. [Google Scholar] [CrossRef] [PubMed]
- Cheong, W.L.; Mohan, D.; Warren, N.; Reidpath, D.D. Multiple Sclerosis in the Asia Pacific Region: A Systematic Review of a Neglected Neurological Disease. Front. Neurol. 2018, 9, 432. [Google Scholar] [CrossRef] [PubMed]
- Vakrakou, A.G.; Tzanetakos, D.; Evangelopoulos, M.E.; Fragoulis, G.E.; Kazakou, P.; Lekka, E.; Kafasi, N.; Tzartos, J.S.; Andreadou, E.; Koutsis, G.; et al. IgG4-related autoimmune manifestations in Alemtuzumab-treated multiple sclerosis patients. J. Neuroimmunol. 2021, 361, 577759. [Google Scholar] [CrossRef]
- Korn, T.; Reddy, J.; Gao, W.; Bettelli, E.; Awasthi, A.; Petersen, T.R.; Backstrom, B.T.; Sobel, R.A.; Wucherpfennig, K.W.; Strom, T.B.; et al. Myelin-specific regulatory T cells accumulate in the CNS but fail to control autoimmune inflammation. Nat. Med. 2007, 13, 423–431. [Google Scholar] [CrossRef]
- Kolinski, T.; Marek-Trzonkowska, N.; Trzonkowski, P.; Siebert, J. Heat shock proteins (HSPs) in the homeostasis of regulatory T cells (Tregs). Cent. Eur. J. Immunol. 2016, 41, 317–323. [Google Scholar] [CrossRef]
- Cossu, D.; Yokoyama, K.; Sechi, L.A.; Hattori, N. Potential of PINK1 and PARKIN Proteins as Biomarkers for Active Multiple Sclerosis: A Japanese Cohort Study. Front. Immunol. 2021, 12, 681386. [Google Scholar] [CrossRef]
- Cossu, D.; Yokoyama, K.; Sato, S.; Noda, S.; Sakanishi, T.; Sechi, L.A.; Hattori, N. Age related immune modulation of experimental autoimmune encephalomyelitis in PINK1 knockout mice. Front. Immunol. 2022, 13, 1036680. [Google Scholar] [CrossRef]
- Souza, F.G.; Cavalcante, G.C. Mitochondria in Mycobacterium Infection: From the Immune System to Mitochondrial Haplogroups. Int. J. Mol. Sci. 2022, 23, 9511. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Japan | Italy | |||
|---|---|---|---|---|
| RR-MS (n = 65) | HCs (n = 65) | RR-MS (n = 65) | HCs (n = 65) | |
| Gender female/male | 45/20 | 40/25 | 46/19 | 47/18 |
| Age mean ± SD | 41 ± 14 | 43 ± 12 | 49 ± 9 | 45 ± 13 |
| Age at onset mean ± SD | 33 ± 10 | 0 | 38 ± 11 | 0 |
| EDSS 1 score median (range) | 2.5 (0–7) | 0 | 3 (0–7.5) | 0 |
| Oligoclonal bands ≥2 (%) | 57 (87) | 0 | 44 (68) | 0 |
| IgG index ≥0.7 (%) | 49 (75) | 0 | 49 (70) | 0 |
| Albumin quotient ≥7 × 10−3 (%) | 16 (25) | 0 | 16 (24) | 0 |
| Treatment 2 Weeks before EAE Induction | Mouse Strain (Age) | EAE Incidence | Mortality | Mean Day of Onset | Peak Clinical Score |
|---|---|---|---|---|---|
| Placebo | C57BL/6 (9 weeks) | 90% | 0% | 11 ± 0.5 * | 3.0 ± 0.5 |
| MAP_HSP70533–545 | C57BL/6 (9 weeks) | 90% | 0% | 10 ± 0.4 | 3.5 ± 0.4 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cossu, D.; Tomizawa, Y.; Yokoyama, K.; Sakanishi, T.; Momotani, E.; Sechi, L.A.; Hattori, N. Mycobacterium avium subsp. paratuberculosis Antigens Elicit a Strong IgG4 Response in Patients with Multiple Sclerosis and Exacerbate Experimental Autoimmune Encephalomyelitis. Life 2023, 13, 1437. https://doi.org/10.3390/life13071437
Cossu D, Tomizawa Y, Yokoyama K, Sakanishi T, Momotani E, Sechi LA, Hattori N. Mycobacterium avium subsp. paratuberculosis Antigens Elicit a Strong IgG4 Response in Patients with Multiple Sclerosis and Exacerbate Experimental Autoimmune Encephalomyelitis. Life. 2023; 13(7):1437. https://doi.org/10.3390/life13071437
Chicago/Turabian StyleCossu, Davide, Yuji Tomizawa, Kazumasa Yokoyama, Tamami Sakanishi, Eiichi Momotani, Leonardo A. Sechi, and Nobutaka Hattori. 2023. "Mycobacterium avium subsp. paratuberculosis Antigens Elicit a Strong IgG4 Response in Patients with Multiple Sclerosis and Exacerbate Experimental Autoimmune Encephalomyelitis" Life 13, no. 7: 1437. https://doi.org/10.3390/life13071437
APA StyleCossu, D., Tomizawa, Y., Yokoyama, K., Sakanishi, T., Momotani, E., Sechi, L. A., & Hattori, N. (2023). Mycobacterium avium subsp. paratuberculosis Antigens Elicit a Strong IgG4 Response in Patients with Multiple Sclerosis and Exacerbate Experimental Autoimmune Encephalomyelitis. Life, 13(7), 1437. https://doi.org/10.3390/life13071437