Abstract
Copepods are proven nutritious food sources for the mariculture/larviculture industry, however, unreliable methods for mass production of copepods are a major bottleneck. In this study, we modified a previously reported inorganic fertilization method (N: 700 μg L−1 and P: 100 μg L−1) by the addition of iron (Fe: 10 μg L−1, using FeSO4·7H2O) (+Fe treatment) and compared its suitability for copepod culture (Pseudodiaptomus annandalei) to the original method (control). The experiment was conducted outdoors in 1000 L tanks for 15 days. The addition of iron prolonged the growth phase of the phytoplankton and resulted in the production of significantly more small phytoplankton (0.45–20 μm, average 2.01 ± 0.52 vs. 9.03 ± 4.17 µg L−1 in control and +Fe, respectively) and adult copepods (control: 195 ± 35, +Fe: 431 ± 109 ind L−1), whereas copepodid-stage was similar between treatments (control: 511 ± 107 vs. +Fe: 502 ± 68 ind L−1). Although adding iron increased the cost of production by 23% compared to the control, the estimated net profit was 97% greater. We concluded that inorganic fertilization, with the addition of iron (Fe: 10 μg L−1), could be an effective method for the mass production of copepods for larviculture.
1. Introduction
Larviculture is the first step in aquaculture. Over the decades, billions of food-fish larvae belonging to only a small number of species have been produced, while the larvae of the majority of other attempted fish species have failed to thrive [1]. Among the numerous difficulties faced in fish larviculture, the primary challenge is to provide live food of suitable size and nutritional quality to the first-feeding larvae [2,3,4,5]. Traditionally, rotifers, Artemia nauplii, and copepods have been provided as prey for fish larvae [1,6,7]. Although rotifers and Artemia are relatively easy to culture, their culture vessels must often be enriched with microalgae to increase their nutritional value, especially with respect to HUFA (highly unsaturated fatty acids), DHA (docosahexaenoic acid, 22:6n-3), and EPA (eicosapentaenoic acid, 20:5n-3), which are essential for the survival and growth of fish larvae [8,9]. Copepods, on the other hand, have a fatty acid composition already dominated by DHA, EPA, and saturated fatty acids [10,11], and many studies have confirmed their nutritional value for larval fish [9,12,13].
Copepods are one of the more common live feeds used in aquaculture [14,15], such as Acartia tonsa [16], and Cyclopina kasignete [17] species. The majority of commercial copepod products are collected from extensive outdoor culture ponds or as a by-product of fish ponds [18,19,20,21]. To enhance copepod production in outdoor culture ponds, various organic materials such as animal manure [22], soybean meal [23], alfalfa meal [24], and fish solubles [25] have been used traditionally to fertilize the ponds. Through the decomposition of the organic matter by bacteria, nutrients such as nitrate and phosphorus are released into the water, thereby enhancing phytoplankton growth, and the microalgae are then consumed by the copepods [22,26]. Production of this type is not reliable in terms of both copepod quantity and quality [18] since the nutrient contents and the lengths of decomposition of various organic fertilizers vary. Large-scale use of organic fertilizers leads to negative side effects such as the accumulation of nitrogenous wastes [27,28,29] causing the proliferation of harmful algae, eutrophication of water bodies [30,31], deteriorating water quality due to low oxygen [32], and high pH [25], and other extreme conditions in the ponds. In addition, different types of fish pathogens such as the microsporidian parasite Enterocytozoon hepatopenaei and the bacterium Vibrio parahaemolyticus can grow in such conditions [25], which might cause mass mortality of larval fishes when fed with those products. The commercial products grown in extensive outdoor culture ponds are not pure culture but are mixtures of different groups of crustaceans such as cyclopoids, calanoids, cladocerans, and ostracods [25]. These mixtures of zooplankton are not the ideal food for a lot of larval fish with very small mouth gapes [4,25,33]. Therefore, alternative applications of inorganic nutrients need to be explored in the mass production of copepods.
Inorganic fertilization has been widely used in agriculture on various crops but is relatively little used in aquaculture [32,33,34,35,36,37]. Previous studies have shown that compared to the traditional organic fertilization methods, the addition of inorganic nitrogen and phosphorus to aquaculture ponds results in fewer filamentous blue-green algae, but enhances more of the small unicellular algae that serve as the main food source for different sizes of zooplankton [4,35,36,37,38,39]. Using the inorganic fertilization method in culturing coral reef fish larvae has revealed that unicellular diatoms such as Coscinodiscus spp., Navicula spp., Nitzschia seriata, and Nitzschia spp. could be enhanced, and more small size zooplankton could be produced [4,33]. In order to mass produce unicellular phytoplankton for zooplankton, pure cultures of several algal species, such as Isochrysis galbana, Nannochloropsis oculata, and Tetraselmis chui have been developed [1]. The monoculture protocol of these algal species has been applied in many studies. Laboratory-scale attempts to augment the production of copepods using algal culture media such as F/2 medium [40] have been used in culturing Bestiolina similis [41], Schmackeria poplesia [42], and A. grani [43]. The results were encouraging, but using the algal culture media would not be cost-effective for industrial-scale mass rearing of copepods [44]. Moreover, monoculture of different algal species may occupy large space in an aquaculture facility, thus reducing the capacity to culture the intended copepod.
To date, only a few efforts at mass production of copepods using inorganic fertilization have been reported. Mischke and Zimba [26] found that applying only inorganic fertilizer to channel catfish nursery ponds at an initial concentration of ∼20 kg/ha N and 2 kg/ha P, followed by twice a week applications of half the initial quantity for 3–4 weeks, increased the concentration of zooplankton suitable as feed for fish with larger mouth gapes. Hong and Tew [25] evaluated the feasibility of using 700 µg L−1 N and 100 µg L−1 P for mass production of the calanoid copepod Pseudodiaptomus annandalei in 1000 L culture tanks, with the results that pathogen-free copepodites and adult copepods were significantly higher than with organic fertilization. Other benefits such as no development of filamentous algae in the tanks and lower total operating costs made it a profitable method for commercial copepod producers [25].
Iron (Fe) is involved in many physiological functions such as photosynthesis [45,46] and nitrogen absorption [47,48] and is a limiting nutrient for phytoplankton growth [49,50,51]. In addition to iron supplementation in aquaponic systems [52], dietary supplementation with inorganic iron has been tried in fish [53]. Adding iron has been shown to be effective in increasing the productivity of cultured phytoplankton [54], and also influences the fatty acid and sterol composition of phytoplankton [55]; this in turn affects the egg production rate of copepods that feed on the algae [55]. Until now, there has been no study on the effects of adding iron to aquaculture media. Since the inorganic fertilization method could potentially be very useful for the mass production of copepods [25], combining it with the addition of iron might further enhance copepod production.
The immediate goal of the present study was to assess how well the inorganic fertilization method we previously employed for the mass production of copepods [25] can be optimized by adding iron as culturing proceeds. The long-term goal is to improve the methodology for providing large quantities of larval and adult copepods for use in larviculture and thereby advance the large-scale rearing of additional fish species in aquaculture in the future.
2. Materials and Methods
2.1. Experimental Setup
We conducted the experiments outdoors at the National Museum of Marine Biology & Aquarium, Taiwan, in June, using ten 1000 L round fiberglass tanks that were filled with unfiltered natural seawater. After filling the tanks, no additional water changes or flows would be made for the entire experiment period. The inorganic nitrogen and phosphorus concentrations of each tank were measured daily and maintained at N: 700 μg L−1 and P: 100 μg L−1 for all tanks. Among them, five served as controls (Control, N = 5) while the iron was added to the other five at a concentration of 10 μg L−1 daily (+Fe treatment, N = 5). The sources of N and P were NH4NO3 and H3PO4 (SIGMA-ALDRICH, St. Louis, MO, USA) [25], respectively, whereas iron was added as FeSO4·7H2O (J.T. Baker, Radnor, PA, USA) [56]. Adult calanoid copepods, P. annandalei, which is commonly found in the coastal areas in Taiwan and is the dominant copepod species in the waters adjacent to the museum [25], were inoculated in each tank at a density of 10 ind L−1 on day 1. Previous studies have confirmed that the species is suitable prey for larval fishes [1,57,58,59] and pose no threat to their predator. The experiment lasted for 15 days since our objective is to mass produce the copepod in a short period of time and harvest them at the highest density possible (around day-15 from previous experiments) and restart the culturing process. We did not inoculate monoculture algae, therefore the algal compositions grown in the tanks were those from the natural seawater at the start of the experiment.
2.2. Physicochemical Analyses
The temperature, pH, dissolved oxygen (DO), and salinity were measured daily at 1100~1300 h with a handheld multi-parameter meter (YSI Professional Plus, YSI, Yellow Springs, OH, USA) during the entire experiment, same procedures as in [25].
The dissolved inorganic nitrogen (NH3-N, NO2-N, and NO3-N), phosphorus (PO4-P), and total Fe concentrations of water samples that had been filtered through 0.45 μm filter paper were analyzed daily for each tank. We used HACH water analysis products (HACH, Loveland, CO, USA) such as the ammonia kit (salicylate method 8155), nitrite kit (diazotization method 8507), phosphorus kit (ascorbic acid method 8048), and total iron kit (FerroVer® method 8008) in the analyses, and measured the nutrient concentrations with a spectrophotometer (Synergy H4 Hybrid Reader, BioTek Instruments, Winooski, VT, USA) [25]. Nitrate (NO3−) was reduced to NO2− [60] before using the HACH nitrite kit.
2.3. Biological Analyses
Chl a-bearing algae were separated into two size categories: 0.45–20 μm and >20 μm, by filtering 200 mL of seawater taken daily from each tank through filter paper (Advantec, Tokyo, Japan) of two pore sizes, 0.45 μm, and 20 μm, respectively. A 0.45–20 μm fraction was calculated by subtracting the amount of Chl a on 20 μm filter paper from those on 0.45 μm filter paper. A previous study found that culture tanks could become overgrown with filamentous algae [25]; therefore, in the present experiment, we collected benthic algae from an area of 7.5 cm × 2.5 cm of the wall (30 cm below the surface) and bottom (150 cm below the surface) of each tank on day 15. The phytoplankton and benthic algae were then extracted with acetone [4,33], and the Chl a concentrations of the extracts were measured with a spectrophotometer (Hitachi U-5100, Hitachi, Tokyo, Japan). The taxonomical composition and density of the phytoplankton were counted and identified under a light microscope to the nearest taxon possible [4,33,35].
To assess the zooplankton, a 1 L water sample was taken daily from each tank after using a stirrer to vigorously disturb the tanks for 30 s. Samples were fixed using 5% formaldehyde, then the water was filtered through a 25 μm mesh, and all the zooplankton (non-copepod, copepod nauplii, and copepodids + adult copepods) were identified and counted under a compound microscope. We dried the copepods from each sample at 70 °C for 3 days in an oven to obtain the total copepod dry weight, and measured them using a 6-digit digital analytical microbalance (XP2U Ultra Micro Balance, Mettler-Toledo, Columbus, OH, USA), and used the formulae from Blanda, et al. [61] and Rayner, et al. [62] to calculate the total daily dried weights.
The population growth rates (r) for total copepod between control and +Fe treatments were calculated from the exponential phase of each population growth using the equation: r = ln(Nt+1 − Nt)/t, where Nt = population density at time t, and Nt+1 = population density after time t+1, and t is the time interval (d) [63].
The cost per unit dry weight of copepods was calculated for each treatment by dividing the total cost of the fertilizers (N, P, and Fe) by the total dry weight of copepods produced during the experimental period. We also calculated the net income over the 15-day period, assuming the sale at the average market price (2.20 NT$ g−1) [25] of the total quantity of copepods produced per tank, minus the total cost per tank.
2.4. Statistical Analysis
We used one-way repeated measures analysis of variance (RM-ANOVA) to determine the effects of adding iron on various biological and physicochemical parameters, with sampling date treated as the repeated factor. Benthic algal Chl a concentrations, total cost, and net income of copepods were compared between treatments by using t-tests. In order to meet the assumptions of normality and homogeneity of variance, we transformed all the data when necessary. We used SigmaPlot 12.5 (SPSS 1997) in all statistical analyses, and α = 0.05 was considered statistically significant [25].
3. Results
During the experiment, the temperature, pH, and DO were significantly higher in the +Fe treatment tanks than in the control tanks, while the salinity was similar between the two (Figure 1).
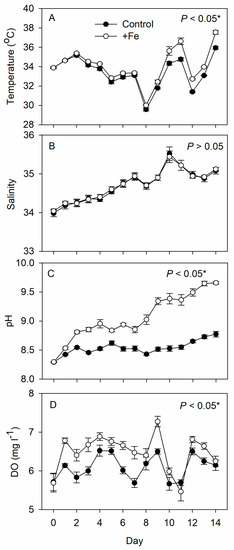
Figure 1.
Daily measurements of (A) temperature, (B) salinity, (C) pH, and (D) dissolved oxygen in the control (N = 5) and +Fe (N = 5) treatment tanks during the 14-day experimental period (mean ± SD). The p-value in each panel indicates the significance level (*, α = 0.05) of the treatment effect based on repeated measures ANOVA.
The NO3-N and PO4-P concentrations were significantly lower in the +Fe treatment tanks than in the controls, while the NH3-N and NO2-N concentrations were similar between the two (Figure 2). Although 10 μg L−1 Fe was added daily to each +Fe treatment tank (i.e., 0.05 g FeSO4·7H2O per tank), the concentration of iron in these tanks always remained below the HACH iron kit’s detection limit.
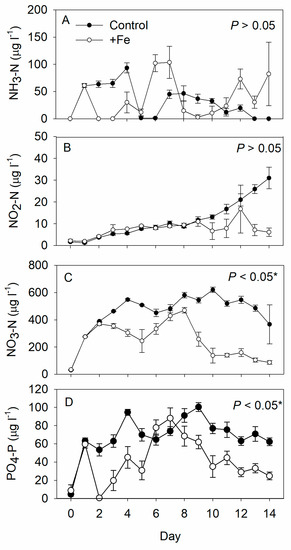
Figure 2.
Daily measurements of (A) ammonium-nitrogen (NH3-N), (B) nitrite-nitrogen (NO2-N), (C) nitrate-nitrogen (NO3-N), and (D) phosphate (PO4-P) concentrations in the control (N = 5), and +Fe (N = 5) treatment tanks during the 14-day experimental period records (mean ± SD). The p-value in each panel indicates the significance level (*, α = 0.05) of the treatment effect based on repeated measures ANOVA.
The Chl a concentration of smaller phytoplankton (0.45–20 μm) was significantly higher in the +Fe treatment tanks than in the control tanks (Figure 3A), while that of the larger phytoplankton (>20 μm) was similar between the two (Figure 3B). Phytoplankton of both sizes began to grow on day 2 in both treatments (Figure 3). At the beginning of the experiment (Day 0~2), about 95% of the phytoplankton were diatom Chaetoceros spp. and shifted to green algae Chlamydomonas spp. and dinoflagellates Gymnodinium spp. in both treatments. Chlamydomonas spp. remained at about 74~78%, while Gymnodinium spp. at 18~24% between Day 4~12 in the control tanks; whereas in +Fe treatment tanks, Chlamydomonas spp. composed of more than 95% of the total phytoplankton densities from Day 8 until the end of the experiment. Although no visible benthic filamentous algae were found with either treatment, the benthic algae on the wall and bottom in the +Fe treatment tanks (2.83 ± 0.63 and 9.43 ± 1.27 µg cm−2, respectively) were significantly higher than in the control tanks (0.25 ± 0.15 and 3.88 ± 0.54 µg cm−2, respectively).
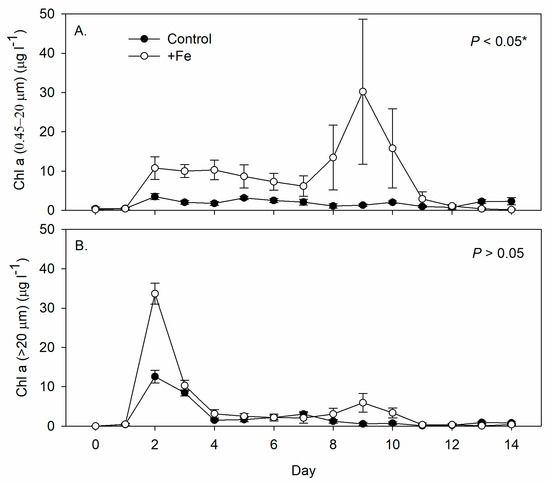
Figure 3.
Daily measurements of (A) smaller phytoplankton chlorophyll a concentration (0.45–20 µm, µg L−1), and (B) larger phytoplankton chlorophyll a concentration (20 µm, µg L−1) in the control (N = 5) and +Fe (N = 5) treatment tanks during the 14-day experimental period (mean ± SD). The p-value in each panel indicates the significance level (*, α = 0.05) of the treatment effect based on repeated measures ANOVA.
Copepod nauplii started to appear 4 days after fertilization, peaked after 8 days, and declined thereafter under both treatments (Figure 4A), with no significant difference in abundance between the control and +Fe tanks (p > 0.05). Adult copepod abundance (including copepodids) was significantly higher in the +Fe treatment tanks, especially after day 8, and remained so until the end of the experiment (Figure 4B). Total copepod dry weight (Figure 4C) followed the same trend as for adults and copepodids and was significantly higher in the +Fe treatment tanks than in the controls (p < 0.05).
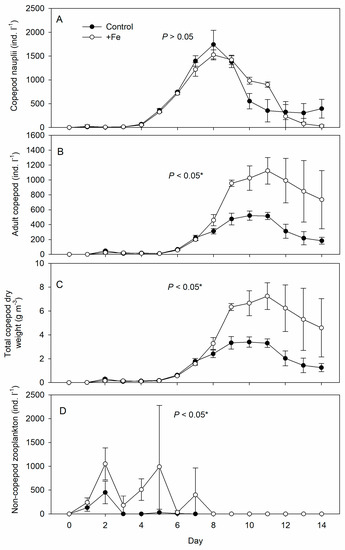
Figure 4.
Daily measurements of abundance (density) of (A) copepod nauplii, (B) adult copepod, (C) total copepod dry weight, and (D) non-copepod zooplankton in the control (N = 5) and +Fe (N = 5) treatment tanks during the 14-day experimental period (mean ± SD). The p-value on each panel indicates the significance level (*, α = 0.05) of the treatment effect based on repeated measures ANOVA.
Non-copepod zooplankton, mainly ciliates: tintinnids and Strombidium spp., appeared early during the experiment but decreased as copepods became dominant (Figure 4D). Their density fluctuated and was significantly higher in +Fe treatment tanks than in the controls (p < 0.05).
The population growth rates from the exponential phase of population growth (Day 4–9) were 0.6008x + 2.7036 and 0.6762x + 2.1586, for control and +Fe treatment, respectively (Figure 5).
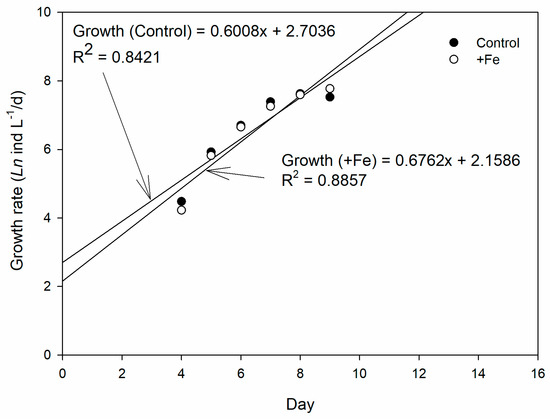
Figure 5.
The growth rate of copepod in the control (N = 5) and +Fe (N = 5) treatment tanks during the 14-day experimental period.
The calculation of total costs and net profits showed that the cost of producing copepods was 23% higher with the +Fe treatment (NT$0.69 ± 0.10 g−1 dry weight) than in the controls (NT$0.56 ± 0.07 g−1 dry weight), but the expected net profit was also significantly (97%) higher with the +Fe treatment (NT$11.04 ± 2.49 g−1 dry weight) than for the controls (NT$5.61 ± 0.83 g−1 dry weight) (t-test, p < 0.05).
4. Discussion
Copepods are a proven live feed for newly hatched fish larvae, especially for some larvae of coral reef fish with small mouth gapes [9,37,64,65,66]. They can enhance the survival and growth rate of fish larvae and reduce the incidence of deformities. Access to a reliable supply of copepods on a commercial scale has been a bottleneck faced by aquaculturists hoping to use copepods in their operations on a daily basis. To overcome this shortfall, several methods, as well as specialized equipment for the large-scale production of copepods, have been devised [18,25,67,68]. For example, Prado-Cabrero, Herena-Garcia, and Nolan [68] designed a monoalgal bioreactor that uses the microalga Tetraselmis chui to continuously produce the harpacticoid copepod Tigriopus californicus [68]. Sarkisian, Lemus, Apeitos, Blaylock, and Saillant [67] developed an indoor batch-culture system for intensive production of the calanoid copepod A. tonsa, featuring integrated grow-out and egg-production units that can produce nauplii daily. Hong and Tew [25] did a pilot study adding a precise ratio of inorganic nitrogen and phosphorus fertilizers to culture tanks in order to mass-produce the calanoid copepod P. annandalei. All of these efforts were aimed at helping to regularize the mass production of copepods to supply the aquafeed industry with marine larval fish culture.
Our previous study [25] showed that inorganic fertilization of culture tanks could produce pathogen-free copepods at a cheaper cost per unit output than the commonly used organic fertilization method. However, the Chl a concentration of phytoplankton usually declined rapidly in the tanks even when nitrogen and phosphorus were still abundant [4,25], thus affecting copepod production. Since iron is involved in phytoplankton photosynthesis [45], and is a trace element necessary for phytoplankton growth [49,50,51], we modified our inorganic fertilization method by adding iron into the copepod culture tanks. The concentration we employed (10 μg L−1 Fe) was higher than that employed by previous researchers: 0.16 μg L−1 Fe by Watson et al. [54] to stimulate limnetic phytoplankton, 0.06 μg L−1 Fe by Takeda [48] to raise oceanic diatoms, and 5.60 μg L−1 Fe by Chen et al. [55] to enhance marine phytoplankton. Since we cannot be certain of the concentration of Fe that will adequately enhance phytoplankton productivity, we chose a much higher but economically feasible concentration. Different iron concentrations should be tried in future studies to lower the cost of copepod production.
In this study, instead of inoculating mono-species culture, the random microalgae from natural seawater were induced by inorganic fertilizer. Since the microalgae composition in the coastal water is commonly seasonal- and regional-specific, one might suspect that the outcomes from using the inorganic fertilization method will vary at different places and seasons, making it unreliable in producing copepods. To the contrary, several larviculture experiments using this method have shown that the phytoplankton and zooplankton productions were consistent and reliable, and higher survival rates of coral reef fish larvae were achieved, irrespective of when and where it was applied [4,33,37,69]. Earlier studies showed unicellular diatoms such as Coscinodiscus spp., Navicula spp., and Nitzschia spp. out-competed filamentous diatom Chaetoceros spp. under inorganic fertilization treatment [4,33], whereas in the present study, the unicellular green algae Chlamydomonas spp. succeeded Chaetoceros spp. The taxonomical compositions of algae were different during different experiments, but it was always smaller unicellular phytoplankton that prevailed. By controlling the ratio and concentration of nitrogen to phosphorus in the water column, we would expect to have phytoplankton compositions of desired sizes, albeit different species in different places or seasons. This method will liberate aquaculturists worldwide from having to monoculture certain algal species in order to feed their monoculture zooplankton, which is then fed to the fish larvae.
During the present experiment, the tanks were set up outdoors and were thus subject to various environmental stresses. As a result of exposure to direct sunlight, for example, the water temperature at 1100~1300 h could reach 38 °C. Fitness trade-offs have been reported for the intertidal copepod T. californicus under extremely high temperatures [70], but for other copepod species such conditions can be ameliorated by providing sufficient food, similar to what was found by Hammock, et al. [71] in that low food availability narrows the tolerance of the copepod Eurytemora affinis to salinity [71]. The high Chl a concentration in our study indicates that there was a high abundance of phytoplankton serving as food for the copepods, which likely offset any temperature stress experienced by the copepods in this study. Moreover, as the temperature goes up, dissolved oxygen tends to be reduced to the detriment of many aquatic organisms [72]. In our study, the mean dissolved oxygen was always 5.00 mg L−1 or higher, thus, above the level at which low oxygen negatively affects the survival and egg production of copepods [73]. This was also likely due to the presence of a high density of phytoplankton in the culture tanks and the resulting production of oxygen as a product of photosynthesis. A high pH can result in a cessation of egg production or an increased nauplii mortality in some species of copepod [74], and our previous study of organic fertilization of culture tanks showed that the growth of nauplii into copepodites and adult copepods was constrained at pH 9.5 [25]. The latter effect was, however, negligible in the present study when iron was added along with inorganic nitrogen and phosphorus. Since nutrient deficiency may reduce the efficiency of energy transfer at the base of food webs by altering the chemical composition of phytoplankton [75], the high quantity of the phytoplankton in our study may have alleviated the otherwise negative impact of environmental stresses on the copepod population.
The concentrations of nutrients such as NO3-N and PO4-P were significantly lower overall in the +Fe treatment tanks than in the control tanks, with the beginning of their decline co-occurring with or immediately preceding the peaks on day 9 in Chl a concentration attributable to smaller-sized algae and in copepod density. Based on the results, we would suggest applying higher concentrations of nitrogen and phosphorus in the future study along with the addition of iron to further enhance the phytoplankton growth. Although excess phosphate could enhance the massive growth of benthic filamentous algae, such as Tribonema spp. [25,76,77], and suppress the growth of phytoplankton which adversely affect the growth of copepod larvae and adults, however, with or without the addition of iron, we observed no significant growth of filamentous algae during the present study, but only unicellular algae. This further strengthens our argument that precise adjustment of the nutrient concentrations and N:P ratio, as enabled by the inorganic fertilization method, provides better conditions for the growth of copepod nauplii than organic fertilization does.
5. Conclusions
Commercially available copepod products are produced by applying various organic materials as fertilizer to extensive outdoor culture ponds or as a by-product of fish ponds. As a result, the quantity and quality of the products are inconsistent. Our previous study found that using inorganic fertilizer was better as compared to the organic method in terms of quantity and quality of copepod production. In this study, we further found that by adding iron as part of an inorganic fertilization protocol for rearing copepods in large culture tanks, the growth of phytoplankton lasted much longer, and a significantly higher density of copepodids and adult copepods could be obtained. Adding iron increased the cost, but the net expected profit was almost twice as high with the addition of iron than without. We contend that inorganic fertilization with the addition of iron, if conducted at a large enough scale, could be an effective method for the mass production of copepods as live feed for use in fish larviculture.
Author Contributions
G.-K.H.: Data curation, Formal analysis, Writing-Original draft preparation, Visualization, Investigation. J.K.: Data analysis and validation. K.S.T.: Conceptualization, Methodology, Supervision, Validation, Writing- Reviewing and Editing, Funding acquisition. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Ministry of Science and Technology of Taiwan (MOST 109-2611-M-291-001 and MOST 110-2611-M-291-001) to KST.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data will be provided whenever requested.
Acknowledgments
This work was supported by grants from Taiwan’s Ministry of Science and Technology (MOST) to KST (MOST 109-2611-M-291-001 and MOST 110-2611-M-291-001). The authors would like to thank Mark J. Grygier for careful English proofreading and suggestions on the manuscript.
Conflicts of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
- Liao, I.C.; Su, H.M.; Chang, E.Y. Techniques in finfish larviculture in Taiwan. Aquaculture 2001, 200, 1–31. [Google Scholar] [CrossRef]
- de F Côrtes, G.; Tsuzuki, M.Y. Effect of different live food on survival and growth of first feeding barber goby, Elacatinus figaro (Sazima, Moura & Rosa 1997) larvae. Aquac. Res. 2012, 43, 831–834. [Google Scholar]
- Shan, X.; Lin, M. Effects of algae and live food density on the feeding ability, growth and survival of miiuy croaker during early development. Aquaculture 2014, 428, 284–289. [Google Scholar] [CrossRef]
- Tew, K.S.; Chang, Y.C.; Meng, P.J.; Leu, M.Y.; Glover, D.C. Towards sustainable exhibits-application of an inorganic fertilization method in coral reef fish larviculture in an aquarium. Aquac. Res. 2016, 47, 2748–2756. [Google Scholar] [CrossRef]
- Divya, K.R.; AkbarAli, I.; Schmidt, B.V.; John, E.M.; Sureshkumar, S.; Thazhakot Vasunambesan, S. Improvement of nutritional quality of live feed for aquaculture: An overview. Aquac. Res. 2020, 51, 1–17. [Google Scholar]
- Hagiwara, A.; Marcial, H.S. The use of non-Brachionus plicatilis species complex rotifer in larviculture. Hydrobiologia 2019, 844, 163–172. [Google Scholar] [CrossRef]
- Lim, L.C.; Dhert, P.; Sorgeloos, P. Recent developments in the application of live feeds in the freshwater ornamental fish culture. Aquaculture 2003, 227, 319–331. [Google Scholar] [CrossRef]
- Whyte, J.N.; Nagata, W.D. Carbohydrate and fatty acid composition of the rotifer, Brachionus plicatilis, fed monospecific diets of yeast or phytoplankton. Aquaculture 1990, 89, 263–272. [Google Scholar] [CrossRef]
- Jepsen, P.M.; van Someren Gréve, H.; Jørgensen, K.N.; Kjær, K.G.; Hansen, B.W. Evaluation of high-density tank cultivation of the live-feed cyclopoid copepod Apocyclops royi (Lindberg 1940). Aquaculture 2021, 533, 736125. [Google Scholar] [CrossRef]
- Olivotto, I.; Buttino, I.; Borroni, M.; Piccinetti, C.; Malzone, M.; Carnevali, O. The use of the Mediterranean calanoid copepod Centropages typicus in Yellowtail clownfish (Amphiprion clarkii) larviculture. Aquaculture 2008, 284, 211–216. [Google Scholar] [CrossRef]
- Olivotto, I.; Capriotti, F.; Buttino, I.; Avella, A.; Vitiello, V.; Maradonna, F.; Carnevali, O. The use of harpacticoid copepods as live prey for Amphiprion clarkii larviculture: Effects on larval survival and growth. Aquaculture 2008, 274, 347–352. [Google Scholar] [CrossRef]
- Lavens, P.; Sorgeloos, P. Manual on the Production and Use of Live Food for Aquaculture; Food and Agriculture Organization (FAO): Rome, Italy, 1996. [Google Scholar]
- Shields, R.J.; Bell, J.G.; Luizi, F.S.; Gara, B.; Bromage, N.R.; Sargent, J.R. Natural copepods are superior to enriched Artemia nauplii as feed for halibut larvae (Hippoglossus hippoglossus) in terms of survival, pigmentation and retinal morphology: Relation to dietary essential fatty acids. J. Nutr. 1999, 129, 1186–1194. [Google Scholar] [CrossRef]
- Piasecki, W.; Goodwin, A.E.; Eiras, J.C.; Nowak, B.F. Importance of Copepoda in freshwater aquaculture. Zool. Stud. 2004, 43, 193–205. [Google Scholar]
- Støttrup, J. The elusive copepods: Their production and suitability in marine aquaculture. Aquac. Res. 2000, 31, 703–711. [Google Scholar] [CrossRef]
- Franco, S.C.; Augustin, C.B.; Geffen, A.J.; Dinis, M.T. Growth, egg production and hatching success of Acartia tonsa cultured at high densities. Aquaculture 2017, 468, 569–578. [Google Scholar] [CrossRef]
- Rasdi, N.W.; Qin, J.G. Copepod supplementation as a live food improved growth and survival of Asian seabass Lates calcarifer larvae. Aquac. Res. 2018, 49, 3606–3613. [Google Scholar] [CrossRef]
- Blanda, E.; Drillet, G.; Huang, C.C.; Hwang, J.S.; Højgaard, J.K.; Jakobsen, H.H.; Rayner, T.A.; Su, H.M.; Hansen, B.W. An analysis of how to improve production of copepods as live feed from tropical Taiwanese outdoor aquaculture ponds. Aquaculture 2017, 479, 432–441. [Google Scholar] [CrossRef]
- Luo, X.; Li, C.; Huang, X. Effect of diet on the development, survival, and reproduction of the calanoid copepod Pseudodiaptomus dubia. J. Oceanol. Limnol. 2019, 37, 1756–1767. [Google Scholar] [CrossRef]
- Li, C.; Luo, X.; Huang, X.; Gu, B. Influences of temperature on development and survival, reproduction and growth of a calanoid copepod (Pseudodiaptomus dubia). Sci. World J. 2009, 9, 866–879. [Google Scholar] [CrossRef]
- Lindley, L.C.; Phelps, R.P. Production and collection of copepod nauplii from brackish water ponds. J. Appl. Aquac. 2009, 21, 96–109. [Google Scholar] [CrossRef]
- Kang’ombe, J.; Brown, J.A.; Halfyard, L.C. Effect of using different types of organic animal manure on plankton abundance, and on growth and survival of Tilapia rendalli (Boulenger) in ponds. Aquac. Res. 2006, 37, 1360–1371. [Google Scholar] [CrossRef]
- Adewumi, A. The growth and gonadal maturation of the African catfish, Clarias gariepinus (Burchell) broodstock fed differently heated soybean-based diets. Aquac. Nutr. 2006, 12, 267–274. [Google Scholar] [CrossRef]
- Kaatz, S.E.; Morris, J.E.; Rudacille, J.B.; Johnson, J.A.; Clayton, R.D. Role of organic fertilizers in walleye (Sander vitreus) production in plastic-lined culture ponds. Aquac. Res. 2011, 42, 490–498. [Google Scholar] [CrossRef]
- Hong, G.K.; Tew, K.S. The Advantages of inorganic fertilization for the mass production of copepods as food for fish larvae in aquaculture. Life 2022, 12, 441. [Google Scholar] [CrossRef]
- Mischke, C.C.; Zimba, P.V. Plankton community responses in earthen channel catfish nursery ponds under various fertilization regimes. Aquaculture 2004, 233, 219–235. [Google Scholar] [CrossRef]
- Zeng, C.; Romano, N. Nitrogenous wastes: Often overlooked pollutants in aquatic environments. J. Mar. Sci. Res. Dev. 2013, 3, 1. [Google Scholar] [CrossRef]
- Hargreaves, J.A. Nitrogen biogeochemistry of aquaculture ponds. Aquaculture 1998, 166, 181–212. [Google Scholar] [CrossRef]
- Tomasso, J. Toxicity of nitrogenous wastes to aquaculture animals. Rev. Fish. Sci. 1994, 2, 291–314. [Google Scholar] [CrossRef]
- Ayoub, A.T. Fertilizers and the environment. Nutr. Cycl. Agroecosystems 1999, 55, 117–121. [Google Scholar] [CrossRef]
- Glibert, P.M.; Maranger, R.; Sobota, D.J.; Bouwman, L. The Haber Bosch–harmful algal bloom (HB–HAB) link. Environ. Res. Lett. 2014, 9, 105001. [Google Scholar] [CrossRef]
- Qin, J.; Culver, D.A. The survival and growth of larval walleye, Stizostedion vitreum, and trophic dynamics in fertilized ponds. Aquaculture 1992, 108, 257–276. [Google Scholar] [CrossRef]
- Tew, K.S.; Meng, P.J.; Lin, H.S.; Chen, J.H.; Leu, M.Y. Experimental evaluation of inorganic fertilization in larval giant grouper (Epinephelus lanceolatus Bloch) production. Aquac. Res. 2013, 44, 439–450. [Google Scholar] [CrossRef]
- Jacob, A.P.; Culver, D.A. Experimental evaluation of the impacts of reduced inorganic phosphorus fertilization rates on juvenile saugeye production. Aquaculture 2010, 304, 22–33. [Google Scholar] [CrossRef]
- Tew, K.S.; Conroy, J.D.; Culver, D.A. Effects of lowered inorganic phosphorus fertilization rates on pond production of percid fingerlings. Aquaculture 2006, 255, 436–446. [Google Scholar] [CrossRef]
- Culver, D.A. Effects of the N: P ratio in fertilizer for fish hatchery ponds. Int. Ver. Für Theor. Und Angew. Limnol. Verh. 1991, 24, 1503–1507. [Google Scholar] [CrossRef]
- Kuo, J.; Chen, C.Y.; Han, C.C.; Ju, Y.M.; Tew, K.S. Analyses of diet preference of larval orange-spotted grouper (Epinephelus coioides) grown under inorganic fertilization method using next-generation sequencing. Aquaculture 2021, 531, 735916. [Google Scholar] [CrossRef]
- Culver, D.A.; Madon, S.P.; Qin, J. Percid pond production techniques: Timing, enrichment, and stocking density manipulation. J. Appl. Aquac. 1994, 2, 9–32. [Google Scholar] [CrossRef]
- Qin, J.; Madon, S.P.; Culver, D.A. Effect of larval walleye (Stizostedion vitreum) and fertilization on the plankton community: Implications for larval fish culture. Aquaculture 1995, 130, 51–65. [Google Scholar] [CrossRef]
- Guillard, R.R.; Ryther, J.H. Cyclotella nana Hustedt, and Detonula confervacea (Cleve) Gran. Can. J. Microbiol. 1962, 8, 229–239. [Google Scholar]
- Camus, T.; Zeng, C.; McKinnon, A.D. Egg production, egg hatching success and population increase of the tropical paracalanid copepod, Bestiolina similis (Calanoida: Paracalanidae) fed different microalgal diets. Aquaculture 2009, 297, 169–175. [Google Scholar] [CrossRef]
- Liu, G.-X.; Xu, D.-H. Feeding, egg production and laboratory culture of Schmackeria poplesia Shen (Copepoda: Calanoida). Aquac. Res. 2010, 41, 1817–1826. [Google Scholar] [CrossRef]
- Nogueira, N.; Sumares, B.; Andrade, C.A.P.; Afonso, A. The effects of temperature and photoperiod on egg hatching success, egg production and population growth of the calanoid copepod, Acartia grani (Calanoida: Acartiidae). Aquac. Res. 2018, 49, 93–103. [Google Scholar] [CrossRef]
- Abate, T.G.; Nielsen, R.; Nielsen, M.; Drillet, G.; Jepsen, P.M.; Hansen, B.W. Economic feasibility of copepod production for commercial use: Result from a prototype production facility. Aquaculture 2015, 436, 72–79. [Google Scholar] [CrossRef]
- Geider, R.J.; La Roche, J. The role of iron in phytoplankton photosynthesis, and the potential for iron-limitation of primary productivity in the sea. Photosynth. Res. 1994, 39, 275–301. [Google Scholar] [CrossRef] [PubMed]
- Roche, J.L.; Geider, R.J.; Graziano, L.M.; Murray, H.; Lewis, K. Induction of specific proteins in eukaryotic algae grown under iron-, phosphorus-, or nitrogen-deficient conditions. J. Phycol. 1993, 29, 767–776. [Google Scholar] [CrossRef]
- Rueler, J.G.; Ades, D.R. The role of iron nutrition in photosynthesis and nitrogen assimilation in Scenedesmus quadricauda (Chlorophyceae). J. Phycol. 1987, 23, 452–457. [Google Scholar] [CrossRef]
- Takeda, S. Influence of iron availability on nutrient consumption ratio of diatoms in oceanic waters. Nature 1998, 393, 774–777. [Google Scholar] [CrossRef]
- Martin, J.H.; Gordon, M.; Fitzwater, S.E. The case for iron. Limnol. Oceanogr. 1991, 36, 1793–1802. [Google Scholar] [CrossRef]
- Hutchins, D.; Hare, C.; Weaver, R.; Zhang, Y.; Firme, G.; DiTullio, G.; Alm, M.; Riseman, S.; Maucher, J.; Geesey, M. Phytoplankton iron limitation in the Humboldt Current and Peru Upwelling. Limnol. Oceanogr. 2002, 47, 997–1011. [Google Scholar] [CrossRef]
- Hutchins, D.A.; Bruland, K.W. Iron-limited diatom growth and Si: N uptake ratios in a coastal upwelling regime. Nature 1998, 393, 561–564. [Google Scholar] [CrossRef]
- Kasozi, N.; Tandlich, R.; Fick, M.; Kaiser, H.; Wilhelmi, B. Iron supplementation and management in aquaponic systems: A review. Aquac. Rep. 2019, 15, 100221. [Google Scholar] [CrossRef]
- Afshari, A.; Sourinejad, I.; Gharaei, A.; Johari, S.A.; Ghasemi, Z. The effects of diet supplementation with inorganic and nanoparticulate iron and copper on growth performance, blood biochemical parameters, antioxidant response and immune function of snow trout Schizothorax zarudnyi (Nikolskii, 1897). Aquaculture 2021, 539, 736638. [Google Scholar] [CrossRef]
- Watson, A.; Liss, P.; Duce, R. Design of a small-scale in situ iron fertilization experiment. Limnol. Oceanogr. 1991, 36, 1960–1965. [Google Scholar] [CrossRef]
- Chen, X.; Wakeham, S.G.; Fisher, N.S. Influence of iron on fatty acid and sterol composition of marine phytoplankton and copepod consumers. Limnol. Oceanogr. 2011, 56, 716–724. [Google Scholar] [CrossRef]
- De Oliveira, M.; Monteiro, M.; Robbs, P.; Leite, S. Growth and chemical composition of Spirulina Maxima and Spirulina Platensis biomass at different temperatures. Aquac. Int. 1999, 7, 261–275. [Google Scholar] [CrossRef]
- Doi, M.; Toledo, J.D.; Golez MS, N.; de los Santos, M.; Ohno, A. Preliminary investigation of feeding performance of larvae of early red-spotted grouper, Epinephelus coioides, reared with mixed zooplankton. In Live Food in Aquaculture; Springer: Dordrecht, The Netherlands, 1997; pp. 259–263. [Google Scholar]
- Lee, C.H.; Dahms, H.U.; Cheng, S.H.; Souissi, S.; Schmitt, F.G.; Kumar, R.; Hwang, J.S. Predation of Pseudodiaptomus annandalei (Copepoda: Calanoida) by the grouper fish fry Epinephelus coioides under different hydrodynamic conditions. J. Exp. Mar. Biol. Ecol. 2010, 393, 17–22. [Google Scholar] [CrossRef]
- Chen, Q.; Sheng, J.; Lin, Q.; Gao, Y.; Lv, J. Effect of salinity on reproduction and survival of the copepod Pseudodiaptomus annandalei Sewell, 1919. Aquaculture 2006, 258, 575–582. [Google Scholar] [CrossRef]
- Pai, S.C.; Riley, J.P. Determination of nitrate in the presence of nitrite in natural waters by flow injection analysis with a non-quantitative on-line cadmium redactor. Int. J. Environ. Anal. Chem. 1994, 57, 263–277. [Google Scholar] [CrossRef]
- Blanda, E.; Drillet, G.; Huang, C.C.; Hwang, J.S.; Jakobsen, H.H.; Rayner, T.A.; Su, H.M.; Wu, C.H.; Hansen, B.W. Trophic interactions and productivity of copepods as live feed from tropical Taiwanese outdoor aquaculture ponds. Aquaculture 2015, 445, 11–21. [Google Scholar] [CrossRef]
- Rayner, T.A.; Jørgensen, N.O.; Blanda, E.; Wu, C.H.; Huang, C.C.; Mortensen, J.; Hwang, J.S.; Hansen, B.W. Biochemical composition of the promising live feed tropical calanoid copepod Pseudodiaptomus annandalei (Sewell 1919) cultured in Taiwanese outdoor aquaculture ponds. Aquaculture 2015, 441, 25–34. [Google Scholar] [CrossRef]
- Kumari, S.; Dhanker, R.; Kumar, R. Does maternal exposure to diatom polyunsaturated aldehyde enhance offspring fitness in limnetic zooplankton Echinisca triserialis (Crustacea: Claodcera)? Fundam. Appl. Limnol. 2022, 195, 325–341. [Google Scholar] [CrossRef]
- Ajiboye, O.; Yakubu, A.; Adams, T.; Olaji, E.; Nwogu, N. A review of the use of copepods in marine fish larviculture. Rev. Fish Biol. Fish. 2011, 21, 225–246. [Google Scholar] [CrossRef]
- Chen, J.Y.; Zeng, C. The effects of live prey and greenwater on the early larval rearing of orchid dottyback Pseudochromis fridmani. Aquaculture 2021, 543, 737008. [Google Scholar] [CrossRef]
- Zeng, C.; Shao, L.; Ricketts, A.; Moorhead, J. The importance of copepods as live feed for larval rearing of the green mandarin fish Synchiropus splendidus. Aquaculture 2018, 491, 65–71. [Google Scholar] [CrossRef]
- Sarkisian, B.L.; Lemus, J.T.; Apeitos, A.; Blaylock, R.B.; Saillant, E.A. An intensive, large-scale batch culture system to produce the calanoid copepod, Acartia tonsa. Aquaculture 2019, 501, 272–278. [Google Scholar] [CrossRef]
- Prado-Cabrero, A.; Herena-Garcia, R.; Nolan, J.M. Intensive production of the harpacticoid copepod Tigriopus californicus in a zero-effluent ‘green water’ bioreactor. Sci. Rep. 2022, 12, 466. [Google Scholar] [CrossRef]
- Leu, M.Y.; Hsu, Y.C.; Tu, Y.H.; Chiu, P.S.; Yu, B.H.; Wang, J.B.; Tew, K.S.; Meng, P.J. Natural spawning, early development and first successful hatchery production of the bluestreak cleaner wrasse, Labroides dimidiatus (Valenciennes, 1839), with application of an inorganic fertilization method in larviculture. Aquaculture 2022, 553, 738056. [Google Scholar] [CrossRef]
- Willett, C.S. Potential fitness trade-offs for thermal tolerance in the intertidal copepod Tigriopus californicus. Evol. Int. J. Org. Evol. 2010, 64, 2521–2534. [Google Scholar] [CrossRef]
- Hammock, B.G.; Lesmeister, S.; Flores, I.; Bradburd, G.S.; Hammock, F.H.; Teh, S.J. Low food availability narrows the tolerance of the copepod Eurytemora affinis to salinity, but not to temperature. Estuaries Coasts 2016, 39, 189–200. [Google Scholar] [CrossRef]
- Ringler, N.H.; Hall, J.D. Effects of logging on water temperature, and dissolved oxygen in spawning beds. Trans. Am. Fish. Soc. 1975, 104, 111–121. [Google Scholar] [CrossRef]
- Sedlacek, C.; Marcus, N.H. Egg production of the copepod Acartia tonsa: The influence of hypoxia and food concentration. J. Exp. Mar. Biol. Ecology. 2005, 318, 183–190. [Google Scholar] [CrossRef]
- Hansen, B.W.; Hansen, P.J.; Nielsen, T.G.; Jepsen, P.M. Effects of elevated pH on marine copepods in mass cultivation systems: Practical implications. J. Plankton Res. 2017, 39, 984–993. [Google Scholar] [CrossRef]
- Bi, R.; Sommer, U. Food quantity and quality interactions at phytoplankton–zooplankton interface: Chemical and reproductive responses in a calanoid copepod. Front. Mar. Science. 2020, 7, 274. [Google Scholar] [CrossRef]
- Davis, A.K.; Anderson, R.S.; Spierling, R.; Leader, S.; Lesne, C.; Mahan, K.; Lundquist, T.; Benemann, J.R.; Lane, T.; Polle, J.E.W. Characterization of a novel strain of Tribonema minus demonstrating high biomass productivity in outdoor raceway ponds. Bioresour. Technol. 2021, 331, 125007. [Google Scholar] [CrossRef]
- Cheng, P.; Cheng, J.J.; Cobb, K.; Zhou, C.; Zhou, N.; Addy, M.; Chen, P.; Yan, X.; Ruan, R. Tribonema sp. and Chlorella zofingiensis co-culture to treat swine wastewater diluted with fishery wastewater to facilitate harvest. Bioresour. Technol. 2020, 297, 122516. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).