
Microbiome Changes in Layer Pullets Reared in Floor Pens along the Growth Period

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Housing Conditions
2.2. Cecum and Ileum Sampling
2.3. DNA Extraction and Microbiome 16S rRNA Sequencing
2.4. Microbiome Diversity and Abundance Analyses
2.5. LEfSe Analysis
3. Results
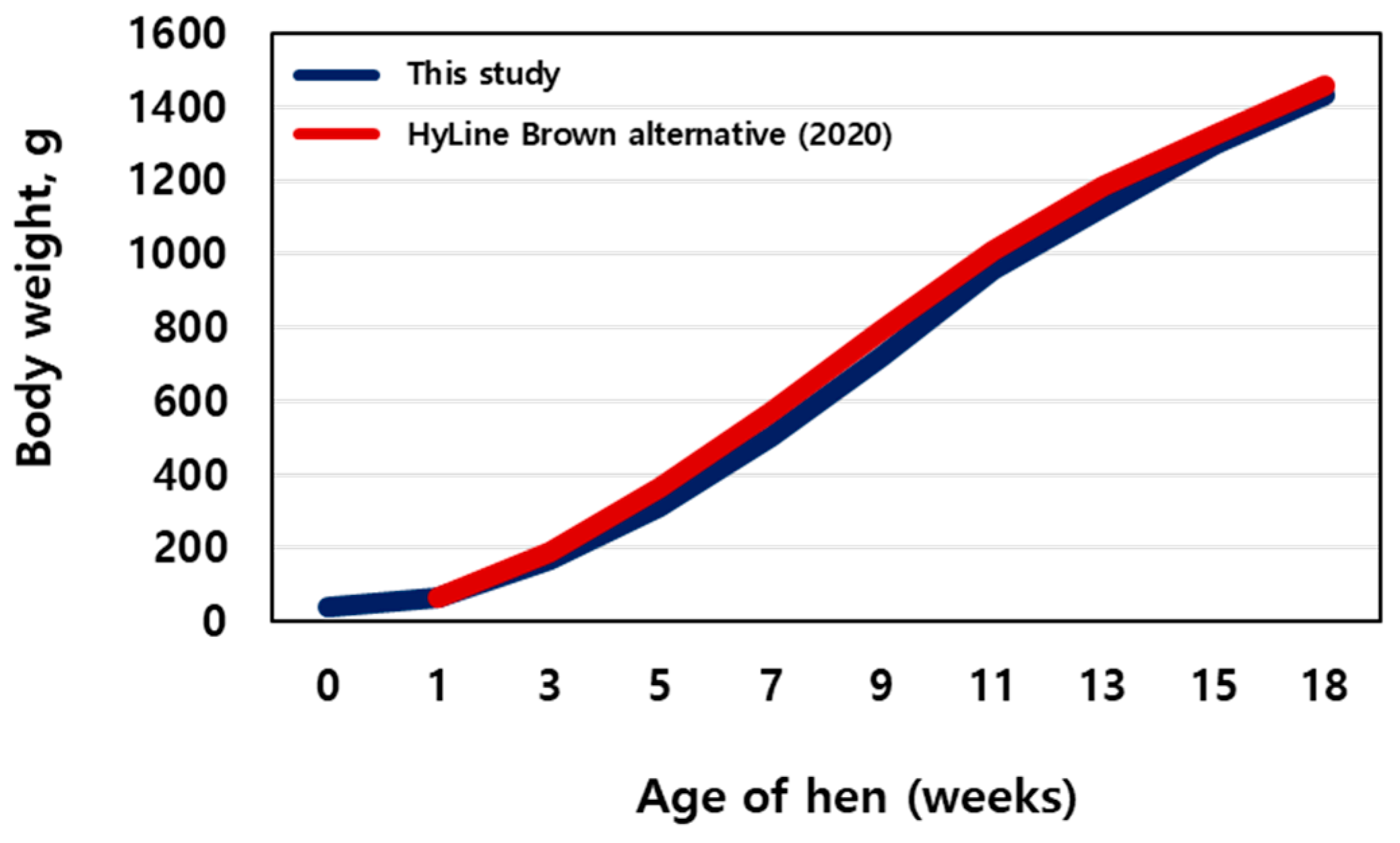
3.1. Weight
3.2. Alpha Diversity
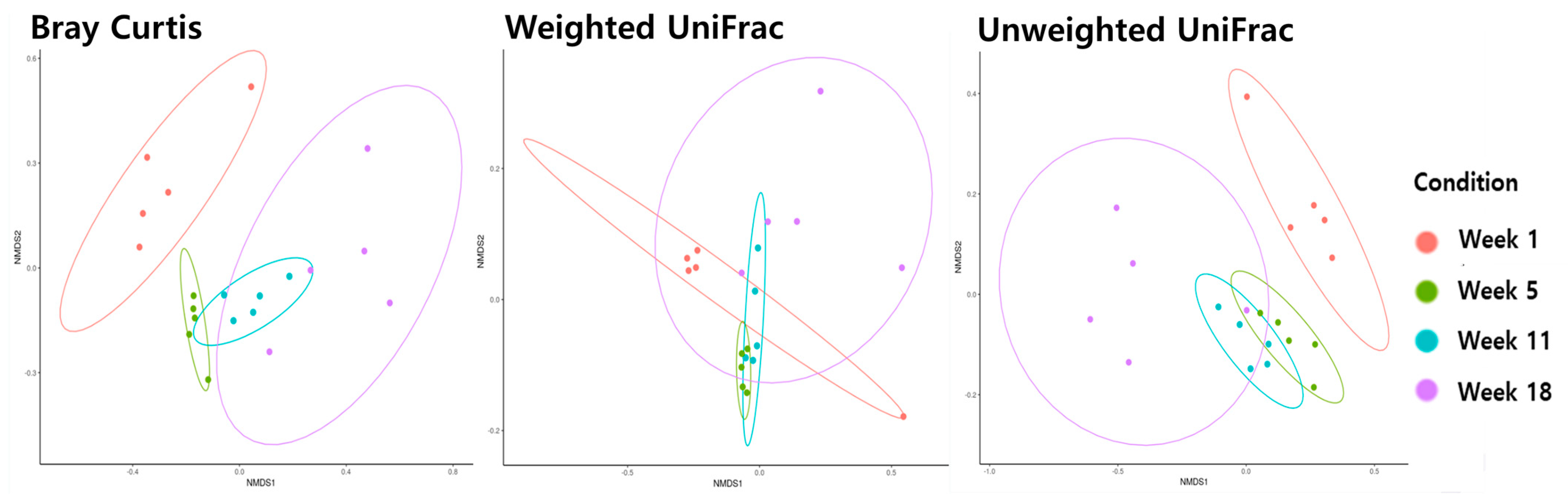
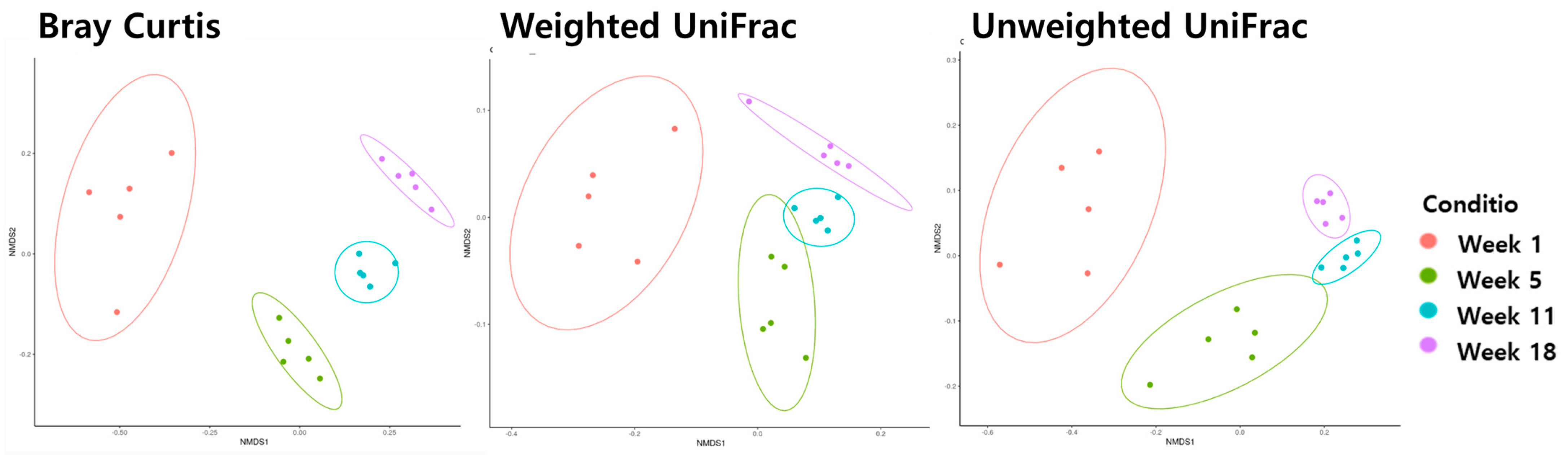
3.3. Beta Diversity
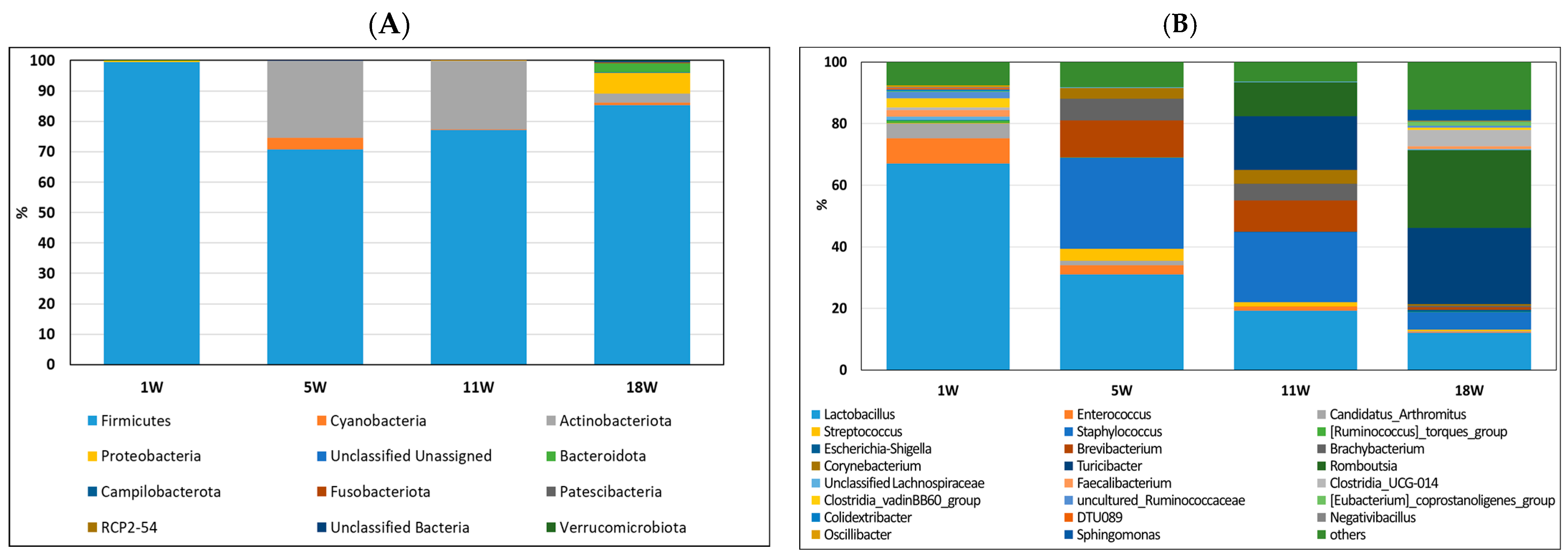
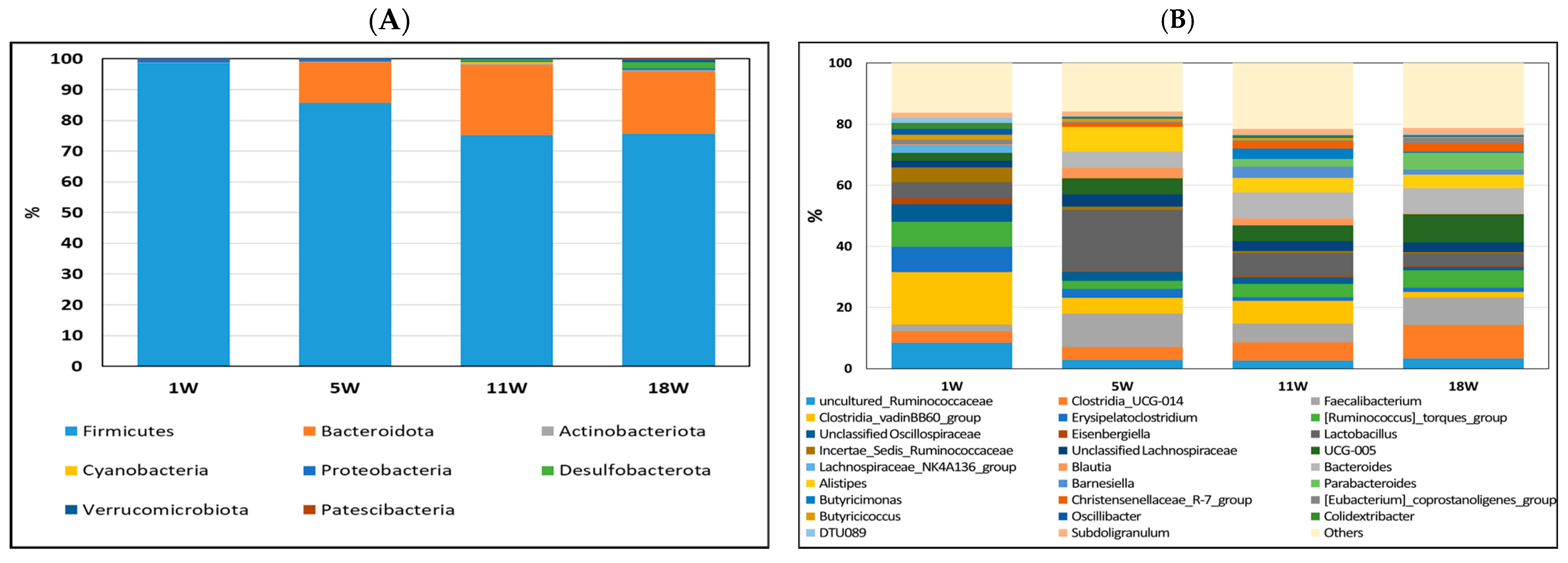
3.4. Composition of Gut Bacteria
3.5. LDA Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bryden, W.L.; Li, X.; Ruhnke, I.; Zhang, D.; Shini, S. Nutrition, feeding and laying hen welfare. Anim. Prod. Sci. 2021, 61, 893–914. [Google Scholar] [CrossRef]
- Liu, Y.; Yan, T.; Ren, Z.; Yang, X. Age-associated changes in caecal microbiome and their apparent correlations with growth performances of layer pullets. Anim. Nutr. 2021, 7, 841–848. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T. Regulation of intestinal epithelial permeability by tight junctions. Cell. Mol. Life Sci. 2013, 70, 631–659. [Google Scholar] [CrossRef] [PubMed]
- Shang, Y.; Kumar, S.; Oakley, B.; Kim, W.K. Chicken gut microbiota: Importance and detection technology. Front. Vet. Sci. 2018, 5, 254. [Google Scholar] [CrossRef] [PubMed]
- Adil, S.; Magray, S.N. Impact and manipulation of gut microflora in poultry: A review. J. Anim. Vet. Adv. 2012, 11, 873–877. [Google Scholar] [CrossRef]
- Lozupone, C.A.; Stombaugh, J.I.; Gordon, J.I.; Jansson, J.K.; Knight, R. Diversity, stability and resilience of the human gut microbiota. Nature 2012, 489, 220–230. [Google Scholar] [CrossRef]
- Antonissen, G.; Eeckhaut, V.; Van Driessche, K.; Onrust, L.; Haesebrouck, F.; Ducatelle, R.; Moore, R.J.; Van Immerseel, F. Microbial shifts associated with necrotic enteritis. Avian Pathol. 2016, 45, 308–312. [Google Scholar] [CrossRef]
- Yeoman, C.J.; Chia, N.; Jeraldo, P.; Sipos, M.; Goldenfeld, N.D.; White, B.A. The microbiome of the chicken gastrointestinal tract. Anim. Health Res. Rev. 2012, 13, 89–99. [Google Scholar] [CrossRef]
- Videnska, P.; Sedlar, K.; Lukac, M.; Faldynova, M.; Gerzova, L.; Cejkova, D.; Sisak, F.; Rychlik, I. Succession and replacement of bacterial populations in the caecum of egg laying hens over their whole life. PLoS ONE 2014, 9, e115142. [Google Scholar] [CrossRef]
- Bavananthasivam, J.; Astill, J.; Matsuyama-Kato, A.; Taha-Abdelaziz, K.; Shojadoost, B.; Sharif, S. Gut microbiota is associated with protection against Marek’s disease virus infection in chickens. Virology 2021, 553, 122–130. [Google Scholar] [CrossRef]
- Waite, D.W.; Taylor, M.W. Characterizing the avian gut microbiota: Membership, driving influences, and potential function. Front. Microbiol. 2014, 5, 223. [Google Scholar] [CrossRef]
- Diaz Carrasco, J.M.; Casanova, N.A.; Fernández Miyakawa, M.E. Microbiota, gut health and chicken productivity: What is the connection? Microorganisms 2019, 7, 374. [Google Scholar] [CrossRef] [PubMed]
- Pan, D.; Yu, Z. Intestinal microbiome of poultry and its interaction with host and diet. Gut Microbes 2014, 5, 108–119. [Google Scholar] [CrossRef] [PubMed]
- Borda-Molina, D.; Seifert, J.; Camarinha-Silva, A. Current perspectives of the chicken gastrointestinal tract and its microbiome. Comput. Struct. Biotechnol. J. 2018, 16, 131–139. [Google Scholar] [CrossRef]
- Dethlefsen, L.; McFall-Ngai, M.; Relman, D.A. An ecological and evolutionary perspective on human–microbe mutualism and disease. Nature 2017, 449, 811–818. [Google Scholar] [CrossRef] [PubMed]
- Faseleh Jahromi, M.; Wesam Altaher, Y.; Shokryazdan, P.; Ebrahimi, R.; Ebrahimi, M.; Idrus, Z.; Tufarelli, V.; Liang, J.B. Dietary supplementation of a mixture of Lactobacillus strains enhances performance of broiler chickens raised under heat stress conditions. Int. J. Biometeorol. 2016, 60, 1099–1110. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Yao, M.; Lv, L.; Ling, Z.; Li, L. The human microbiota in health and disease. Engineering 2017, 3, 71–82. [Google Scholar] [CrossRef]
- Yan, W.; Sun, C.; Yuan, J.; Yang, N. Gut metagenomic analysis reveals prominent roles of Lactobacillus and cecal microbiota in chicken feed efficiency. Sci. Rep. 2017, 7, 45308. [Google Scholar] [CrossRef]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome. Biol. 2011, 12, 1–18. [Google Scholar] [CrossRef]
- Coloe, P.J.; Bagust, T.J.; Ireland, L. Development of the normal gastrointestinal microflora of specific pathogen-free chickens. Epidemiol. Infect. 1984, 92, 79–87. [Google Scholar] [CrossRef]
- Lu, J.; Idris, U.; Harmon, B.; Hofacre, C.; Maurer, J.J.; Lee, M.D. Diversity and succession of the intestinal bacterial community of the maturing broiler chicken. Appl. Environ. Microbiol. 2003, 69, 6816–6824. [Google Scholar] [CrossRef]
- Hume, M.E.; Kubena, L.F.; Edrington, T.S.; Donskey, C.J.; Moore, R.W.; Ricke, S.C.; Nisbet, D.J. Poultry digestive microflora biodiversity as indicated by denaturing gradient gel electrophoresis. Poult. Sci. 2003, 82, 1100–1107. [Google Scholar] [CrossRef] [PubMed]
- Ngunjiri, J.M.; Taylor, K.J.; Abundo, M.C.; Jang, H.; Elaish, M.; Kc, M.; Ghorbani, A.; Wijeratne, S.; Weber, B.P.; Johnson, T.J.; et al. Farm stage, bird age, and body site dominantly affect the quantity, taxonomic composition, and dynamics of respiratory and gut microbiota of com-mercial layer chickens. Appl. Environ. Microbiol. 2019, 85, e03137-18. [Google Scholar] [CrossRef] [PubMed]
- Wielen PW, J.J.; Keuzenkamp, D.A.; Lipman, L.V.; Knapen, F.; Biesterveld, S. Spatial and temporal variation of the intestinal bacterial community in commercially raised broiler chickens during growth. Microb. Ecol. 2002, 44, 286–293. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wu, S.; Li, X.; Yan, T.; Duan, Y.; Yang, X.; Duan, Y.; Sun, Q.; Yang, X. Simultaneous supplementation of Bacillus subtilis and antibiotic growth promoters by stages improved intestinal function of pullets by altering gut microbiota. Front. Microbiol. 2018, 9, 2328. [Google Scholar] [CrossRef] [PubMed]
- Ballou, A.L.; Ali, R.A.; Mendoza, M.A.; Ellis, J.C.; Hassan, H.M.; Croom, W.J.; Koci, M.D. Development of the chick microbiome: How early exposure influences future microbial diversity. Front. Vet. Sci. 2016, 3, 2. [Google Scholar] [CrossRef]
- Dai, D.; Wu, S.G.; Zhang, H.J.; Qi, G.H.; Wang, J. Dynamic alterations in early intestinal development, microbiota and metabolome induced by in ovo feeding of L-arginine in a layer chick model. J. Anim. Sci. Biotechnol. 2020, 11, 19. [Google Scholar] [CrossRef]
- Zhao, L.; Wang, G.; Siegel, P.; He, C.; Wang, H.; Zhao, W.; Zhai, Z.; Tian, F.; Zhao, J.; Zhang, H.; et al. Quantitative genetic background of the host influences gut microbiomes in chickens. Sci. Rep. 2013, 3, 1163. [Google Scholar] [CrossRef]
- Nordentoft, S.; Mølbak, L.; Bjerrum, L.; De Vylder, J.; Van Immerseel, F.; Pedersen, K. The influence of the cage system and colonisation of Salmonella Enteritidis on the microbial gut flora of laying hens studied by T-RFLP and 454 py-rosequencing. BMC Microb. 2011, 11, 187. [Google Scholar] [CrossRef]
- Neijat, M.; Habtewold, J.; Shirley, R.B.; Welsher, A.; Barton, J.; Thiery, P.; Kiarie, E. Bacillus subtilis strain DSM 29784 modulates the cecal microbiome, concentration of short-chain fatty acids, and apparent retention of dietary components in shaver white chickens during grower, developer, and laying phases. Appl. Environ. Microbiol. 2019, 85, e00402-19. [Google Scholar] [CrossRef]
- Peng, L.; Li, Z.R.; Green, R.S.; Holzman, I.R.; Lin, J. Butyrate enhances the intestinal barrier by facilitating tight junction assembly via activation of AMP-activated protein kinase in Caco-2 cell monolayers. J. Nutr. 2009, 139, 1619–1625. [Google Scholar] [CrossRef]
- Zaiss, M.M.; Jones, R.M.; Schett, G.; Pacifici, R. The gut-bone axis: How bacterial metabolites bridge the distance. J. Clin. Investig. 2019, 129, 3018–3028. [Google Scholar] [CrossRef] [PubMed]
- Singh, K.M.; Shah, T.; Deshpande, S.; Jakhesara, S.J.; Koringa, P.G.; Rank, D.N.; Joshi, C.G. High through put 16S rRNA gene-based pyrosequencing analysis of the fecal microbiota of high FCR and low FCR broiler growers. Mol. Biol. Rep. 2012, 39, 10595–10602. [Google Scholar] [CrossRef]
- Renouf, M.; Hendrich, S. Bacteroides uniformis is a putative bacterial species associated with the degradation of the isoflavone genistein in human feces. J. Nutr. 2011, 141, 1120–1126. [Google Scholar] [CrossRef] [PubMed]
- Gauffin Cano, P.; Santacruz, A.; Moya, Á.; Sanz, Y. Bacteroides uniformis CECT 7771 ameliorates metabolic and immunological dysfunction in mice with high-fat-diet induced obesity. PLoS ONE 2012, 7, E41079. [Google Scholar] [CrossRef]
- Polansky, O.; Sekelova, Z.; Faldynova, M.; Sebkova, A.; Sisak, F.; Rychlik, I. Important metabolic pathways and biological processes expressed by chicken cecal microbiota. Appl. Environ. Microbiol. 2016, 82, 1569–1576. [Google Scholar] [CrossRef] [PubMed]
- Ndotono, E.W.; Khamis, F.M.; Bargul, J.L.; Tanga, C.M. Gut microbiota shift in layer pullets fed on black soldier fly larvae-based feeds towards enhancing healthy gut microbial community. Sci. Rep. 2022, 12, 16714. [Google Scholar] [CrossRef]
- Ayeni, F.A.; Adeniyi, B.A.; Ogunbanwo, S.T.; Tabasco, R.; Paarup, T.; Peláez, C.; Requena, T. Inhibition of uropathogens by lactic acid bacteria isolated from dairy foods and cow’s intestine in western Nigeria. Arch. Microbiol. 2009, 191, 639–648. [Google Scholar] [CrossRef]
- Vieco-Saiz, N.; Belguesmia, Y.; Raspoet, R.; Auclair, E.; Gancel, F.; Kempf, I.; Drider, D. Benefits and inputs from lactic acid bacteria and their bacteriocins as alternatives to antibiotic growth promoters during food-animal production. Front. Microbiol. 2019, 10, 57. [Google Scholar] [CrossRef]
- Chege, S.M. Opportunistic infections of avians. In The Rasputin Effect: When Commensals and Symbionts Become Parasitic; Springer: Cham, Switzerland, 2016; pp. 221–260. [Google Scholar]
- Xiao, S.; Mi, J.; Chen, Y.; Feng, K.; Mei, L.; Liao, X.; Liao, X.; Wu, Y.; Wang, Y. The abundance and diversity of antibiotic resistance genes in layer chicken ceca is associated with farm enviroment. Front. Microbiol. 2023, 14, 1177404. [Google Scholar] [CrossRef]
- Joat, N.; Bajagai, Y.S.; Van TT, H.; Stanley, D.; Chousalkar, K.; Moore, R.J. The temporal fluctuations and development of faecal microbiota in commercial layer flocks. Anim. Nutr. 2023, 15, 19–209. [Google Scholar] [CrossRef] [PubMed]
- Emami, N.K.; Dalloul, R.A. Centennial Review: Recent developments in host-pathogen interactions during necrotic enteritis in poultry. Poult. Sci. 2021, 100, 101330. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Week | Chao1 | Shannon Entropy | Simpson | Faith pd |
|---|---|---|---|---|
| 1 | 65.7 b | 3.44 | 0.81 | 3.52 b |
| 5 | 60.0 b | 4.28 | 0.92 | 3.80 b |
| 11 | 138.9 ab | 4.26 | 0.87 | 6.13 b |
| 18 | 191.7 a | 4.15 | 0.75 | 18.30 a |
| SEM | 28.63 | 0.561 | 0.053 | 2.232 |
| p value | 0.0139 | 0.6847 | 0.1825 | 0.0006 |
| Week | Chao1 | Shannon Entropy | Simpson | Faith pd |
|---|---|---|---|---|
| 1 | 155.1 b | 5.72 c | 0.96 b | 8.28 d |
| 5 | 286.9 b | 6.36 b | 0.97 b | 14.94 c |
| 11 | 667.5 a | 7.56 a | 0.99 a | 28.46 a |
| 18 | 569.8 a | 7.52 a | 0.99 a | 23.94 b |
| SEM | 45.40 | 0.135 | 0.003 | 1.324 |
| p value | <0.0001 | <0.0001 | <0.0001 | <0.0001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, H.-J.; Kim, H.-S.; Yun, Y.-S.; Shin, H.; Lee, W.-D.; Son, J.; Hong, E.-C.; Jeon, I.-S.; Kang, H.-K. Microbiome Changes in Layer Pullets Reared in Floor Pens along the Growth Period. Life 2023, 13, 2302. https://doi.org/10.3390/life13122302
Kim H-J, Kim H-S, Yun Y-S, Shin H, Lee W-D, Son J, Hong E-C, Jeon I-S, Kang H-K. Microbiome Changes in Layer Pullets Reared in Floor Pens along the Growth Period. Life. 2023; 13(12):2302. https://doi.org/10.3390/life13122302
Chicago/Turabian StyleKim, Hee-Jin, Hyun-Soo Kim, Yeon-Seo Yun, Hyekyoung Shin, Woo-Do Lee, Jiseon Son, Eui-Chul Hong, Ik-Soo Jeon, and Hwan-Ku Kang. 2023. "Microbiome Changes in Layer Pullets Reared in Floor Pens along the Growth Period" Life 13, no. 12: 2302. https://doi.org/10.3390/life13122302
APA StyleKim, H.-J., Kim, H.-S., Yun, Y.-S., Shin, H., Lee, W.-D., Son, J., Hong, E.-C., Jeon, I.-S., & Kang, H.-K. (2023). Microbiome Changes in Layer Pullets Reared in Floor Pens along the Growth Period. Life, 13(12), 2302. https://doi.org/10.3390/life13122302