

Why Does Your Uterus Become Malignant? The Impact of the Microbiome on Endometrial Carcinogenesis

 , ,
, , {kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Review Methodology
3. Endometrial Cancer Classification
4. Factors Affecting Endometrial Carcinogenesis
4.1. Factors Increasing Endometrial Cancer Risk
4.2. Factors Decreasing Endometrial Cancer Risk
5. Uterine Microbiome
5.1. Microbiome of a Healthy Uterus
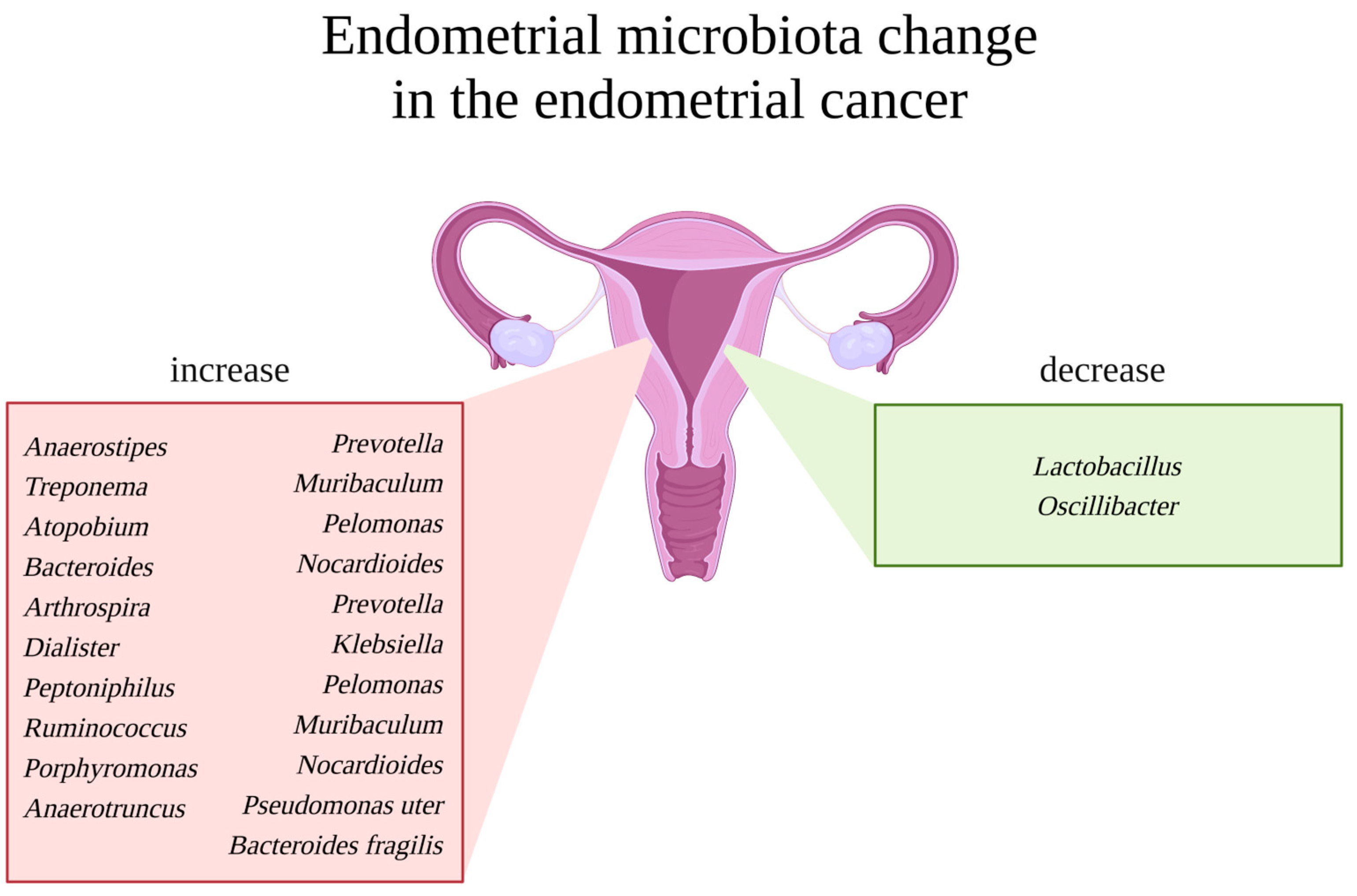
5.2. Microbiome of a Pathological Uterus
6. Implications of Microbiome
7. How Does the Endometrial Tissue Become Malignant?
8. How Does the Gut Influence Endometrial Tissue Carcinogenesis?
9. Limitation of Current Evidence
10. Conclusions
11. Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Endometrial Cancer Statistics|World Cancer Research Fund International. Available online: https://www.wcrf.org/cancer-trends/endometrial-cancer-statistics/ (accessed on 5 August 2022).
- Global Cancer Observatory; WHO. Cancer Today. Available online: https://gco.iarc.fr/today/home (accessed on 11 November 2022).
- Jemal, A.; Clegg, L.X.; Ward, E.; Ries, L.A.G.; Wu, X.; Jamison, P.M.; Wingo, P.A.; Howe, H.L.; Anderson, R.N.; Edwards, B.K. Annual Report to the Nation on the Status of Cancer, 1975–2001, with a Special Feature Regarding Survival. Cancer 2004, 101, 3–27. [Google Scholar] [CrossRef] [PubMed]
- The National Cancer Data Base Report on Endometrial Carcinoma in African-American Women—Hicks—1998—Cancer—Wiley Online Library. Available online: https://acsjournals.onlinelibrary.wiley.com/doi/10.1002/(SICI)1097-0142(19981215)83:12%3C2629::AID-CNCR30%3E3.0.CO;2-2 (accessed on 15 December 2022).
- Endometrial Cancer. Available online: https://www.cancer.org/cancer/endometrial-cancer.html (accessed on 15 December 2022).
- Agostinis, C.; Mangogna, A.; Bossi, F.; Ricci, G.; Kishore, U.; Bulla, R. Uterine Immunity and Microbiota: A Shifting Paradigm. Front. Immunol. 2019, 10, 2387. [Google Scholar] [CrossRef] [PubMed]
- Zhu, N.; Yang, X.; Liu, Q.; Chen, Y.; Wang, X.; Li, H.; Gao, H. “Iron Triangle” of Regulating the Uterine Microecology: Endometrial Microbiota, Immunity and Endometrium. Front. Immunol. 2022, 13, 928475. [Google Scholar] [CrossRef] [PubMed]
- Pioli, P.A.; Weaver, L.K.; Schaefer, T.M.; Wright, J.A.; Wira, C.R.; Guyre, P.M. Lipopolysaccharide-Induced IL-1 Beta Production by Human Uterine Macrophages up-Regulates Uterine Epithelial Cell Expression of Human Beta-Defensin 2. J. Immunol. 2006, 176, 6647–6655. [Google Scholar] [CrossRef] [PubMed]
- Cassetta, L.; Pollard, J.W. Targeting Macrophages: Therapeutic Approaches in Cancer. Nat. Rev. Drug Discov. 2018, 17, 887–904. [Google Scholar] [CrossRef]
- Nakamura, A.; Ooga, T.; Matsumoto, M. Intestinal Luminal Putrescine Is Produced by Collective Biosynthetic Pathways of the Commensal Microbiome. Gut Microbes 2019, 10, 159–171. [Google Scholar] [CrossRef]
- Baker, J.M.; Al-Nakkash, L.; Herbst-Kralovetz, M.M. Estrogen-Gut Microbiome Axis: Physiological and Clinical Implications. Maturitas 2017, 103, 45–53. [Google Scholar] [CrossRef]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 Statement: An Updated Guideline for Reporting Systematic Reviews. BMJ 2021, 372, 105906. [Google Scholar] [CrossRef]
- Page, M.J.; Moher, D.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. PRISMA 2020 Explanation and Elaboration: Updated Guidance and Exemplars for Reporting Systematic Reviews. BMJ 2021, 372, n160. [Google Scholar] [CrossRef]
- Huvila, J.; Pors, J.; Thompson, E.F.; Gilks, C.B. Endometrial Carcinoma: Molecular Subtypes, Precursors and the Role of Pathology in Early Diagnosis. J. Pathol. 2021, 253, 355–365. [Google Scholar] [CrossRef]
- Alexa, M.; Hasenburg, A.; Battista, M.J. The TCGA Molecular Classification of Endometrial Cancer and Its Possible Impact on Adjuvant Treatment Decisions. Cancers 2021, 13, 1478. [Google Scholar] [CrossRef] [PubMed]
- Di Donato, V.; Giannini, A.; Bogani, G. Recent Advances in Endometrial Cancer Management. J. Clin. Med. 2023, 12, 2241. [Google Scholar] [CrossRef] [PubMed]
- Golia D’Augè, T.; Cuccu, I.; Santangelo, G.; Muzii, L.; Giannini, A.; Bogani, G.; Di Donato, V. Novel Insights into Molecular Mechanisms of Endometrial Diseases. Biomolecules 2023, 13, 499. [Google Scholar] [CrossRef]
- World Cancer Research Fund. American Institute for Cancer Research Analysing Research on Cancer Prevention and Survival 2013 Diet, Nutrition, Physical Activity and Endometrial Cancer; World Cancer Research Fund: London, UK, 2018. [Google Scholar]
- Samulak, D.; Wilczak, M.; Englert-Golon, M.; Michalska, M.M. The Diagnostic Value of Evaluating the Maximum Velocity of Blood Flow in the Uterine Arteries of Women with Postmenopausal Bleeding. Arch. Gynecol. Obstet. 2011, 284, 1175–1178. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Hursting, S.D.; Lavigne, J.A.; Berrigan, D.; Perkins, S.N.; Barrett, J.C. Calorie Restriction, Aging, and Cancer Prevention: Mechanisms of Action and Applicability to Humans. Annu. Rev. Med. 2003, 54, 131–152. [Google Scholar] [CrossRef] [PubMed]
- Calle, E.E.; Kaaks, R. Overweight, Obesity and Cancer: Epidemiological Evidence and Proposed Mechanisms. Nat. Rev. Cancer 2004, 4, 579–591. [Google Scholar] [CrossRef]
- Westley, R.L.; May, F.E.B. A Twenty-First Century Cancer Epidemic Caused by Obesity: The Involvement of Insulin, Diabetes, and Insulin-like Growth Factors. Int. J. Endocrinol. 2013, 2013, 632461. [Google Scholar] [CrossRef]
- Key, T.J.; Appleby, P.; Barnes, I.; Reeves, G.; Dorgan, J.F.; Longcope, C.; Franz, C.; Stanczyk, F.Z.; Chang, L.C.; Stephenson, H.E.; et al. Endogenous Sex Hormones and Breast Cancer in Postmenopausal Women: Reanalysis of Nine Prospective Studies. J. Natl. Cancer Inst. 2002, 94, 606–616. [Google Scholar] [CrossRef]
- Wellen, K.E.; Hotamisligil, G.S. Obesity-Induced Inflammatory Changes in Adipose Tissue. J. Clin. Investig. 2003, 112, 1785–1788. [Google Scholar] [CrossRef]
- Rexrode, K.M.; Pradhan, A.; Manson, J.E.; Buring, J.E.; Ridker, P.M. Relationship of Total and Abdominal Adiposity with CRP and IL-6 in Women. Ann. Epidemiol. 2003, 13, 674–682. [Google Scholar] [CrossRef]
- Nagle, C.M.; Olsen, C.M.; Ibiebele, T.I.; Spurdle, A.B.; Webb, P.M. Glycemic Index, Glycemic Load and Endometrial Cancer Risk: Results from the Australian National Endometrial Cancer Study and an Updated Systematic Review and Meta-Analysis. Eur. J. Nutr. 2013, 52, 705–715. [Google Scholar] [CrossRef] [PubMed]
- Mulholland, H.G.; Murray, L.J.; Cardwell, C.R.; Cantwell, M.M. Dietary Glycaemic Index, Glycaemic Load and Endometrial and Ovarian Cancer Risk: A Systematic Review and Meta-Analysis. Br. J. Cancer 2008, 99, 434–441. [Google Scholar] [CrossRef]
- le Roith, D.; Bondy, C.; Yakar, S.; Liu, J.L.; Butler, A. The Somatomedin Hypothesis: 2001. Endocr. Rev. 2001, 22, 53–74. [Google Scholar] [CrossRef] [PubMed]
- Clinical Usefulness of Evaluation of Quality Parameters of Blood Flow: Pulsation Index and Resistance Index in the Uterine Arteries in the Initial Differential Diagnostics of Pathology within the Endometrium. Available online: https://pubmed.ncbi.nlm.nih.gov/20882889/ (accessed on 28 February 2023).
- Westerterp, K.R. Physical Activity and Physical Activity Induced Energy Expenditure in Humans: Measurement, Determinants, and Effects. Front. Physiol. 2013, 4, 90. [Google Scholar] [CrossRef]
- McTiernan, A. Mechanisms Linking Physical Activity with Cancer. Nat. Rev. Cancer 2008, 8, 205–211. [Google Scholar] [CrossRef]
- Moore, S.C.; Gierach, G.L.; Schatzkin, A.; Matthews, C.E. Physical Activity, Sedentary Behaviours, and the Prevention of Endometrial Cancer. Br. J. Cancer 2010, 103, 933–938. [Google Scholar] [CrossRef] [PubMed]
- Giri, A.; Hartmann, K. Re: A Prospective Investigation of Coffee Drinking and Endometrial Cancer Incidence. Int. J. Cancer 2012, 131, 2210–2211. [Google Scholar] [CrossRef]
- Je, Y.; Giovannucci, E. Coffee Consumption and Risk of Endometrial Cancer: Findings from a Large up-to-Date Meta-Analysis. Int. J. Cancer 2012, 131, 1700–1710. [Google Scholar] [CrossRef]
- Gunter, M.J.; Schaub, J.A.; Xue, X.; Freedman, N.D.; Gaudet, M.M.; Rohan, T.E.; Hollenbeck, A.R.; Sinha, R. A Prospective Investigation of Coffee Drinking and Endometrial Cancer Incidence. Int. J. Cancer 2012, 131, E530–E536. [Google Scholar] [CrossRef]
- Sobstyl, M.; Brecht, P.; Sobstyl, A.; Mertowska, P.; Grywalska, E. The Role of Microbiota in the Immunopathogenesis of Endometrial Cancer. Int. J. Mol. Sci. 2022, 23, 5756. [Google Scholar] [CrossRef]
- Turnbaugh, P.J.; Ley, R.E.; Hamady, M.; Fraser-Liggett, C.M.; Knight, R.; Gordon, J.I. The Human Microbiome Project. Nature 2007, 449, 804–810. [Google Scholar] [CrossRef] [PubMed]
- Sender, R.; Fuchs, S.; Milo, R. Revised Estimates for the Number of Human and Bacteria Cells in the Body. PLoS Biol. 2016, 14, e1002533. [Google Scholar] [CrossRef] [PubMed]
- Verstraelen, H.; Vilchez-Vargas, R.; Desimpel, F.; Jauregui, R.; Vankeirsbilck, N.; Weyers, S.; Verhelst, R.; De Sutter, P.; Pieper, D.H.; Van De Wiele, T. Characterisation of the Human Uterine Microbiome in Non-Pregnant Women through Deep Sequencing of the V1-2 Region of the 16S RRNA Gene. PeerJ 2016, 4, e1602. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Song, X.; Wei, W.; Zhong, H.; Dai, J.; Lan, Z.; Li, F.; Yu, X.; Feng, Q.; Wang, Z.; et al. The Microbiota Continuum along the Female Reproductive Tract and Its Relation to Uterine-Related Diseases. Nat. Commun. 2017, 8, 875. [Google Scholar] [CrossRef] [PubMed]
- Winters, A.D.; Romero, R.; Gervasi, M.T.; Gomez-Lopez, N.; Tran, M.R.; Garcia-Flores, V.; Pacora, P.; Jung, E.; Hassan, S.S.; Hsu, C.D.; et al. Does the Endometrial Cavity Have a Molecular Microbial Signature? Sci. Rep. 2019, 9, 9905. [Google Scholar] [CrossRef] [PubMed]
- Hansen, L.K.; Becher, N.; Bastholm, S.; Glavind, J.; Ramsing, M.; Kim, C.J.; Romero, R.; Jensen, J.S.; Uldbjerg, N. The Cervical Mucus Plug Inhibits, but Does Not Block, the Passage of Ascending Bacteria from the Vagina during Pregnancy. Acta Obstet. Gynecol. Scand. 2014, 93, 102–108. [Google Scholar] [CrossRef]
- Altmäe, S.; Franasiak, J.M.; Mändar, R. The Seminal Microbiome in Health and Disease. Nat. Rev. Urol. 2019, 16, 703–721. [Google Scholar] [CrossRef]
- Suarez, S.S.; Pacey, A.A. Sperm Transport in the Female Reproductive Tract. Hum. Reprod. Update 2006, 12, 23–37. [Google Scholar] [CrossRef]
- Zervomanolakis, I.; Ott, H.W.; Hadziomerovic, D.; Mattle, V.; Seeber, B.E.; Virgolini, I.; Heute, D.; Kissler, S.; Leyendecker, G.; Wildt, L. Physiology of Upward Transport in the Human Female Genital Tract. Ann. N. Y. Acad. Sci. 2007, 1101, 1–20. [Google Scholar] [CrossRef]
- Baker, J.M.; Chase, D.M.; Herbst-Kralovetz, M.M. Uterine Microbiota: Residents, Tourists, or Invaders? Front. Immunol. 2018, 9, 208. [Google Scholar] [CrossRef]
- Jeon, S.J.; Cunha, F.; Vieira-Neto, A.; Bicalho, R.C.; Lima, S.; Bicalho, M.L.; Galvão, K.N. Blood as a Route of Transmission of Uterine Pathogens from the Gut to the Uterus in Cows. Microbiome 2017, 5, 109. [Google Scholar] [CrossRef] [PubMed]
- Aagaard, K.; Ma, J.; Antony, K.M.; Ganu, R.; Petrosino, J.; Versalovic, J. The Placenta Harbors a Unique Microbiome. Sci. Transl. Med. 2014, 6, 237ra65. [Google Scholar] [CrossRef] [PubMed]
- Lindheim, L.; Bashir, M.; Münzker, J.; Trummer, C.; Zachhuber, V.; Leber, B.; Horvath, A.; Pieber, T.R.; Gorkiewicz, G.; Stadlbauer, V.; et al. Alterations in Gut Microbiome Composition and Barrier Function Are Associated with Reproductive and Metabolic Defects in Women with Polycystic Ovary Syndrome (PCOS): A Pilot Study. PLoS ONE 2017, 12, e0168390. [Google Scholar] [CrossRef] [PubMed]
- Pereira, N.; Hutchinson, A.P.; Lekovich, J.P.; Hobeika, E.; Elias, R.T. Antibiotic Prophylaxis for Gynecologic Procedures Prior to and during the Utilization of Assisted Reproductive Technologies: A Systematic Review. J. Pathog. 2016, 2016, 4698314. [Google Scholar] [CrossRef] [PubMed]
- Łaniewski, P.; Ilhan, Z.E.; Herbst-Kralovetz, M.M. The Microbiome and Gynaecological Cancer Development, Prevention and Therapy. Nat. Rev. Urol. 2020, 17, 232–250. [Google Scholar] [CrossRef] [PubMed]
- Riganelli, L.; Iebba, V.; Piccioni, M.; Illuminati, I.; Bonfiglio, G.; Neroni, B.; Calvo, L.; Gagliardi, A.; Levrero, M.; Merlino, L.; et al. Structural Variations of Vaginal and Endometrial Microbiota: Hints on Female Infertility. Front. Cell. Infect. Microbiol. 2020, 10, 350. [Google Scholar] [CrossRef] [PubMed]
- Fuhler, G.M. The Immune System and Microbiome in Pregnancy. Best Pract. Res. Clin. Gastroenterol. 2020, 44–45, 101671. [Google Scholar] [CrossRef] [PubMed]
- Robertson, S.A.; Chin, P.Y.; Glynn, D.J.; Thompson, J.G. Peri-Conceptual Cytokines—Setting the Trajectory for Embryo Implantation, Pregnancy and Beyond. Am. J. Reprod. Immunol. 2011, 66, 2–10. [Google Scholar] [CrossRef]
- Park, D.W.; Yang, K.M. Hormonal Regulation of Uterine Chemokines and Immune Cells. Clin. Exp. Reprod. Med. 2011, 38, 179–185. [Google Scholar] [CrossRef]
- Ravel, J.; Gajer, P.; Abdo, Z.; Schneider, G.M.; Koenig, S.S.K.; McCulle, S.L.; Karlebach, S.; Gorle, R.; Russell, J.; Tacket, C.O.; et al. Vaginal Microbiome of Reproductive-Age Women. Proc. Natl. Acad. Sci. USA 2011, 108, 4680–4687. [Google Scholar] [CrossRef]
- Grossman, J.H., III; Adams, R.L.; Hierholzer, W.J.J.; Andriole, V.T. Endometrial and Vaginal Cuff Bacteria Recovered at Elective Hysterectomy during a Trial of Antibiotic Prophylaxis. Am. J. Obstet. Gynecol. 1978, 130, 312–316. [Google Scholar] [PubMed]
- Pezzlo, M.T.; Hesser, J.W.; Morgan, T.; Valter, P.J.; Thrupp, L.D. Improved Laboratory Efficiency and Diagnostic Accuracy with New Double-Lumen-Protected Swab for for Endometrial Specimens. J. Clin. Microbiol. 1979, 9, 56. [Google Scholar] [CrossRef] [PubMed]
- Mishell, D.R.; Bell, J.H.; Good, R.G.; Moyer, D.L. The Intrauterine Device: A Bacteriologic Study of the Endometrial Cavity. Am. J. Obstet. Gynecol. 1966, 96, 119–126. [Google Scholar] [CrossRef]
- Bollinger, C.C. Bacterial Flora of the Nonpregnant Uterus: A New Culture Technic. Obstet. Gynecol. 1964, 23, 251–255. [Google Scholar] [PubMed]
- Butler, B. Value of Endometrial Cultures in Sterility Investigation. Fertil. Steril. 1958, 9, 269–273. [Google Scholar] [CrossRef]
- Møller, B.R.; Kristiansen, F.V.; Thorsen, P.; Frost, L.; Mogensen, S.C. Sterility of the Uterine Cavity. Acta Obstet. Gynecol. Scand. 1995, 74, 216–219. [Google Scholar] [CrossRef]
- Cowling, P.; McCoy, D.R.; Marshall, R.J.; Padfield, C.J.H.; Reeves, D.S. Bacterial Colonization of the Non-Pregnant Uterus: A Study of Pre-Menopausal Abdominal Hysterectomy Specimens. Eur. J. Clin. Microbiol. Infect. Dis. 1992, 11, 204–205. [Google Scholar] [CrossRef]
- Teisala, K. Endometrial Microbial Flora of Hysterectomy Specimens. Eur. J. Obstet. Gynecol. Reprod. Biol. 1987, 26, 151–155. [Google Scholar] [CrossRef]
- Eschenbach, D.A.; Rosene, K.; Tompkins, L.S.; Watkins, H.; Gravett, M.G. Endometrial Cultures Obtained by a Triple-Lumen Method from Afebrile and Febrile Postpartum Women. J. Infect. Dis. 1986, 153, 1038–1045. [Google Scholar] [CrossRef]
- Sparks, R.A.; Purrier, B.G.; Watt, P.J.; Elstein, M. Bacteriological Colonisation of Uterine Cavity: Role of Tailed Intrauterine Contraceptive Device. Br. Med. J. Clin. Res. Ed. 1981, 282, 1189–1191. [Google Scholar] [CrossRef]
- Hemsell, D.L.; Obregon, V.L.; Heard, M.C.; Nobles, B.J. Endometrial Bacteria in Asymptomatic, Nonpregnant Women. J. Reprod. Med. 1989, 34, 872–874. [Google Scholar]
- Walther-António, M.R.S.; Chen, J.; Multinu, F.; Hokenstad, A.; Distad, T.J.; Cheek, E.H.; Keeney, G.L.; Creedon, D.J.; Nelson, H.; Mariani, A.; et al. Potential Contribution of the Uterine Microbiome in the Development of Endometrial Cancer. Genome Med. 2016, 8, 122. [Google Scholar] [CrossRef]
- Moreno, I.; Codoñer, F.M.; Vilella, F.; Valbuena, D.; Martinez-Blanch, J.F.; Jimenez-Almazán, J.; Alonso, R.; Alamá, P.; Remohí, J.; Pellicer, A.; et al. Evidence That the Endometrial Microbiota Has an Effect on Implantation Success or Failure. Am. J. Obstet. Gynecol. 2016, 215, 684–703. [Google Scholar] [CrossRef]
- Mitchell, C.M.; Haick, A.; Nkwopara, E.; Garcia, R.; Rendi, M.; Agnew, K.; Fredricks, D.N.; Eschenbach, D. Colonization of the Upper Genital Tract by Vaginal Bacterial Species in Non-Pregnant Women. Am. J. Obstet. Gynecol. 2015, 212, 611.e1. [Google Scholar] [CrossRef]
- Lu, W.; He, F.; Lin, Z.; Liu, S.; Tang, L.; Huang, Y.; Hu, Z. Dysbiosis of the Endometrial Microbiota and Its Association with Inflammatory Cytokines in Endometrial Cancer. Int. J. Cancer 2021, 148, 1708–1716. [Google Scholar] [CrossRef]
- Davenport, E.R.; Sanders, J.G.; Song, S.J.; Amato, K.R.; Clark, A.G.; Knight, R. The Human Microbiome in Evolution. BMC Biol. 2017, 15, 127. [Google Scholar] [CrossRef]
- Miller, M.B.; Bassler, B.L. Quorum Sensing in Bacteria. Annu. Rev. Microbiol. 2001, 55, 165–199. [Google Scholar] [CrossRef]
- Critchley, H.O.D.; Maybin, J.A.; Armstrong, G.M.; Williams, A.R.W. Physiology of the Endometrium and Regulation of Menstruation. Physiol. Rev. 2020, 100, 1149–1179. [Google Scholar] [CrossRef]
- Shen, H.H.; Zhang, T.; Yang, H.L.; Lai, Z.Z.; Zhou, W.J.; Mei, J.; Shi, J.W.; Zhu, R.; Xu, F.Y.; Li, D.J.; et al. Ovarian Hormones-Autophagy-Immunity Axis in Menstruation and Endometriosis. Theranostics 2021, 11, 3512. [Google Scholar] [CrossRef]
- Critchley, H.O.D.; Babayev, E.; Bulun, S.E.; Clark, S.; Garcia-Grau, I.; Gregersen, P.K.; Kilcoyne, A.; Kim, J.Y.J.; Lavender, M.; Marsh, E.E.; et al. Menstruation: Science and Society. Am. J. Obstet. Gynecol. 2020, 223, 624–664. [Google Scholar] [CrossRef]
- Moreno, I.; Simon, C. Relevance of Assessing the Uterine Microbiota in Infertility. Fertil. Steril. 2018, 110, 337–343. [Google Scholar] [CrossRef]
- Moore, D.E.; Soules, M.R.; Klein, N.A.; Fujimoto, V.Y.; Agnew, K.J.; Eschenbach, D.A. Bacteria in the Transfer Catheter Tip Influence the Live-Birth Rate after in Vitro Fertilization. Fertil. Steril. 2000, 74, 1118–1124. [Google Scholar] [CrossRef]
- Ojosnegros, S.; Seriola, A.; Godeau, A.L.; Veiga, A. Embryo Implantation in the Laboratory: An Update on Current Techniques. Hum. Reprod. Update 2021, 27, 501–530. [Google Scholar] [CrossRef]
- Vallvé-Juanico, J.; Houshdaran, S.; Giudice, L.C. The Endometrial Immune Environment of Women with Endometriosis. Hum. Reprod. Update 2019, 25, 565–592. [Google Scholar] [CrossRef]
- Li, H.; Zang, Y.; Wang, C.; Li, H.; Fan, A.; Han, C.; Xue, F. The Interaction Between Microorganisms, Metabolites, and Immune System in the Female Genital Tract Microenvironment. Front. Cell. Infect. Microbiol. 2020, 10, 609488. [Google Scholar] [CrossRef]
- Jiang, X.-F.; Tang, Q.-L.; Shen, X.-M.; Li, H.-G.; Chen, L.-H.; Wang, X.-Y.; Luo, X.; Lin, Z.-Q.; Jiang, G.-Y. Tumor-Associated Macrophages, Epidermal Growth Factor Receptor Correlated with the Triple Negative Phenotype in Endometrial Endometrioid Adenocarcinoma. Pathol. Res. Pract. 2012, 208, 730–735. [Google Scholar] [CrossRef]
- Jiang, X.F.; Tang, Q.L.; Li, H.G.; Shen, X.M.; Luo, X.; Wang, X.Y.; Lin, Z.Q. Tumor-Associated Macrophages Correlate with Progesterone Receptor Loss in Endometrial Endometrioid Adenocarcinoma. J. Obstet. Gynaecol. Res. 2013, 39, 855–863. [Google Scholar] [CrossRef]
- Kondratiev, S.; Sabo, E.; Yakirevich, E.; Lavie, O.; Resnick, M.B. Intratumoral CD8+ T Lymphocytes as a Prognostic Factor of Survival in Endometrial Carcinoma. Clin. Cancer Res. 2004, 10, 4450–4456. [Google Scholar] [CrossRef]
- ben Yahia, H.; Babay, W.; Bortolotti, D.; Boujelbene, N.; Laaribi, A.B.; Zidi, N.; Kehila, M.; Chelbi, H.; Boudabous, A.; Mrad, K.; et al. Increased Plasmatic Soluble HLA-G Levels in Endometrial Cancer. Mol. Immunol. 2018, 99, 82–86. [Google Scholar] [CrossRef]
- Kurman, R.J.; Carcangiu, M.L.; Herrington, C.S.; Young, R.H. WHO Classification of Tumours of Female Reproductive Organs. 2014. Available online: http://kubalibri.cz/ (accessed on 11 November 2022).
- Androutsopoulos, G.; Styliara, I.; Zarogianni, E.; Lazurko, N.; Valasoulis, G.; Michail, G.; Adonakis, G. The ErbB Signaling Network and Its Potential Role in Endometrial Cancer. Epigenomes 2023, 7, 24. [Google Scholar] [CrossRef]
- Srinivasan, R.; Benton, E.; McCormick, F.; Thomas, H.; Gullick, W.J. Expression of the C-ErbB-3/HER-3 and c-ErbB-4/HER-4 Growth Factor Receptors and Their Ligands, Neuregulin-1 α, Neuregulin-1 β, and Betacellulin, in Normal Endometrium and Endometrial Cancer. Clin. Cancer Res. 1999, 5, 2877–2883. [Google Scholar] [PubMed]
- Ejskjær, K.; Sørensen, B.S.; Poulsen, S.S.; Forman, A.; Nexø, E.; Mogensen, O. Expression of the Epidermal Growth Factor System in Endometrioid Endometrial Cancer. Gynecol. Oncol. 2007, 104, 158–167. [Google Scholar] [CrossRef] [PubMed]
- Reyes, H.D.; Thiel, K.W.; Carlson, M.J.; Meng, X.; Yang, S.; Stephan, J.M.; Leslie, K.K. Comprehensive Profiling of EGFR/HER Receptors for Personalized Treatment of Gynecologic Cancers. Mol. Diagn. Ther. 2014, 18, 137–151. [Google Scholar] [CrossRef] [PubMed]
- Slomovitz, B.M.; Broaddus, R.R.; Burke, T.W.; Sneige, N.; Soliman, P.T.; Wu, W.; Sun, C.C.; Munsell, M.F.; Gershenson, D.M.; Lu, K.H. Her-2/Neu Overexpression and Amplification in Uterine Papillary Serous Carcinoma. J. Clin. Oncol. 2004, 22, 3126–3132. [Google Scholar] [CrossRef]
- Androutsopoulos, G.; Adonakis, G.; Liava, A.; Ravazoula, P.; Decavalas, G. Expression and Potential Role of ErbB Receptors in Type II Endometrial Cancer. Eur. J. Obstet. Gynecol. Reprod. Biol. 2013, 168, 204–208. [Google Scholar] [CrossRef] [PubMed]
- Borella, F.; Carosso, A.R.; Cosma, S.; Preti, M.; Collemi, G.; Cassoni, P.; Bertero, L.; Benedetto, C. Gut Microbiota and Gynecological Cancers: A Summary of Pathogenetic Mechanisms and Future Directions. ACS Infect. Dis. 2021, 7, 987–1009. [Google Scholar] [CrossRef]
- Amabebe, E.; Anumba, D.O.C. Female Gut and Genital Tract Microbiota-Induced Crosstalk and Differential Effects of Short-Chain Fatty Acids on Immune Sequelae. Front. Immunol. 2020, 11, 2184. [Google Scholar] [CrossRef]
- Koh, A.; de Vadder, F.; Kovatcheva-Datchary, P.; Bäckhed, F. From Dietary Fiber to Host Physiology: Short-Chain Fatty Acids as Key Bacterial Metabolites. Cell 2016, 165, 1332–1345. [Google Scholar] [CrossRef]
- Bilotta, A.J.; Cong, Y. Gut Microbiota Metabolite Regulation of Host Defenses at Mucosal Surfaces: Implication in Precision Medicine. Precis Clin. Med. 2019, 2, 110–119. [Google Scholar] [CrossRef]
- Venegas, D.P.; de La Fuente, M.K.; Landskron, G.; González, M.J.; Quera, R.; Dijkstra, G.; Harmsen, H.J.M.; Faber, K.N.; Hermoso, M.A. Short Chain Fatty Acids (SCFAs)Mediated Gut Epithelial and Immune Regulation and Its Relevance for Inflammatory Bowel Diseases. Front. Immunol. 2019, 10, 277. [Google Scholar] [CrossRef]
- Rivière, A.; Selak, M.; Lantin, D.; Leroy, F.; de Vuyst, L. Bifidobacteria and Butyrate-Producing Colon Bacteria: Importance and Strategies for Their Stimulation in the Human Gut. Front. Microbiol. 2016, 7, 979. [Google Scholar] [CrossRef] [PubMed]
- Amabebe, E.; Robert, F.O.; Agbalalah, T.; Orubu, E.S.F. Microbial Dysbiosis-Induced Obesity: Role of Gut Microbiota in Homoeostasis of Energy Metabolism. Br. J. Nutr. 2020, 123, 1127–1137. [Google Scholar] [CrossRef] [PubMed]
- Fattahi, Y.; Heidari, H.R.; Khosroushahi, A.Y. Review of Short-Chain Fatty Acids Effects on the Immune System and Cancer. Food Biosci. 2020, 38, 100793. [Google Scholar] [CrossRef]
- Belizário, J.E.; Faintuch, J. Microbiome and Gut Dysbiosis. Exp. Suppl. 2018, 109, 459–476. [Google Scholar] [PubMed]
- Frosali, S.; Pagliari, D.; Gambassi, G.; Landolfi, R.; Pandolfi, F.; Cianci, R. How the Intricate Interaction among Toll-Like Receptors, Microbiota, and Intestinal Immunity Can Influence Gastrointestinal Pathology. J. Immunol. Res. 2015, 2015, 489821. [Google Scholar] [CrossRef] [PubMed]
- Rinninella, E.; Raoul, P.; Cintoni, M.; Franceschi, F.; Miggiano, G.A.D.; Gasbarrini, A.; Mele, M.C. What Is the Healthy Gut Microbiota Composition? A Changing Ecosystem across Age, Environment, Diet, and Diseases. Microorganisms 2019, 7, 14. [Google Scholar] [CrossRef] [PubMed]
- Dalile, B.; van Oudenhove, L.; Vervliet, B.; Verbeke, K. The Role of Short-Chain Fatty Acids in Microbiota–Gut–Brain Communication. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 461–478. [Google Scholar] [CrossRef]
- Cerf-Bensussan, N.; Gaboriau-Routhiau, V. The Immune System and the Gut Microbiota: Friends or Foes? Nat. Rev. Immunol. 2010, 10, 735–744. [Google Scholar] [CrossRef]
- Wallace, A.E.; Gibson, D.A.; Saunders, P.T.K.; Jabbour, H.N. Inflammatory Events in Endometrial Adenocarcinoma. J. Endocrinol. 2010, 206, 141–157. [Google Scholar] [CrossRef]
- Casaburi, I.; Avena, P.; Lanzino, M.; Sisci, D.; Giordano, F.; Maris, P.; Catalano, S.; Morelli, C.; Andò, S. Chenodeoxycholic Acid through a TGR5-Dependent CREB Signaling Activation Enhances Cyclin D1 Expression and Promotes Human Endometrial Cancer Cell Proliferation. Cell Cycle 2012, 11, 2699–2710. [Google Scholar] [CrossRef]
- Takai, N.; Narahara, H.; Takai, N.; Narahara, H. Human Endometrial and Ovarian Cancer Cells: Histone Deacetylase Inhibitors Exhibit Antiproliferative Activity, Potently Induce Cell Cycle Arrest, and Stimulate Apoptosis. Curr. Med. Chem. 2007, 14, 2548–2553. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.I.; Schultz, C.R.; Buras, A.L.; Friedman, E.; Fedorko, A.; Seamon, L.; Chandramouli, G.V.R.; Maxwell, G.L.; Bachmann, A.S.; Risinger, J.I. Ornithine Decarboxylase as a Therapeutic Target for Endometrial Cancer. PLoS ONE 2017, 12, e0189044. [Google Scholar] [CrossRef] [PubMed]
- Graham, M.E.; Herbert, W.G.; Song, S.D.; Raman, H.N.; Zhu, J.E.; Gonzalez, P.E.; Walther-António, M.R.S.; Tetel, M.J. Gut and Vaginal Microbiomes on Steroids: Implications for Women’s Health. Trends Endocrinol. Metab. 2021, 32, 554–565. [Google Scholar] [CrossRef] [PubMed]
- Perez-Muñoz, M.E.; Arrieta, M.C.; Ramer-Tait, A.E.; Walter, J. A Critical Assessment of the “Sterile Womb” and “in Utero Colonization” Hypotheses: Implications for Research on the Pioneer Infant Microbiome. Microbiome 2017, 5, 48. [Google Scholar] [CrossRef]
- Karstens, L.; Asquith, M.; Caruso, V.; Rosenbaum, J.T.; Fair, D.A.; Braun, J.; Gregory, W.T.; Nardos, R.; McWeeney, S.K. Community Profiling of the Urinary Microbiota: Considerations for Low-Biomass Samples. Nat. Rev. Urol. 2018, 15, 735–749. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morańska, K.; Englert-Golon, M.; Durda-Masny, M.; Sajdak, S.; Grabowska, M.; Szwed, A. Why Does Your Uterus Become Malignant? The Impact of the Microbiome on Endometrial Carcinogenesis. Life 2023, 13, 2269. https://doi.org/10.3390/life13122269
Morańska K, Englert-Golon M, Durda-Masny M, Sajdak S, Grabowska M, Szwed A. Why Does Your Uterus Become Malignant? The Impact of the Microbiome on Endometrial Carcinogenesis. Life. 2023; 13(12):2269. https://doi.org/10.3390/life13122269
Chicago/Turabian StyleMorańska, Katarzyna, Monika Englert-Golon, Magdalena Durda-Masny, Stefan Sajdak, Marlena Grabowska, and Anita Szwed. 2023. "Why Does Your Uterus Become Malignant? The Impact of the Microbiome on Endometrial Carcinogenesis" Life 13, no. 12: 2269. https://doi.org/10.3390/life13122269
APA StyleMorańska, K., Englert-Golon, M., Durda-Masny, M., Sajdak, S., Grabowska, M., & Szwed, A. (2023). Why Does Your Uterus Become Malignant? The Impact of the Microbiome on Endometrial Carcinogenesis. Life, 13(12), 2269. https://doi.org/10.3390/life13122269