
In Vitro Evaluation of Weizmannia coagulans Strain LMG S-31876 Isolated from Fermented Rice for Potential Probiotic Properties, Safety Assessment and Technological Properties


Abstract
:Simple Summary
Abstract

1. Introduction
2. Materials and Methods
2.1. Isolation, Identification and Safe Deposit of Bacteria
2.1.1. Isolation of Bacteria
2.1.2. Morphological and Biochemical Characterisation
2.1.3. Molecular Identification
2.1.4. Safe Deposit
2.2. Evaluation of Probiotic Properties
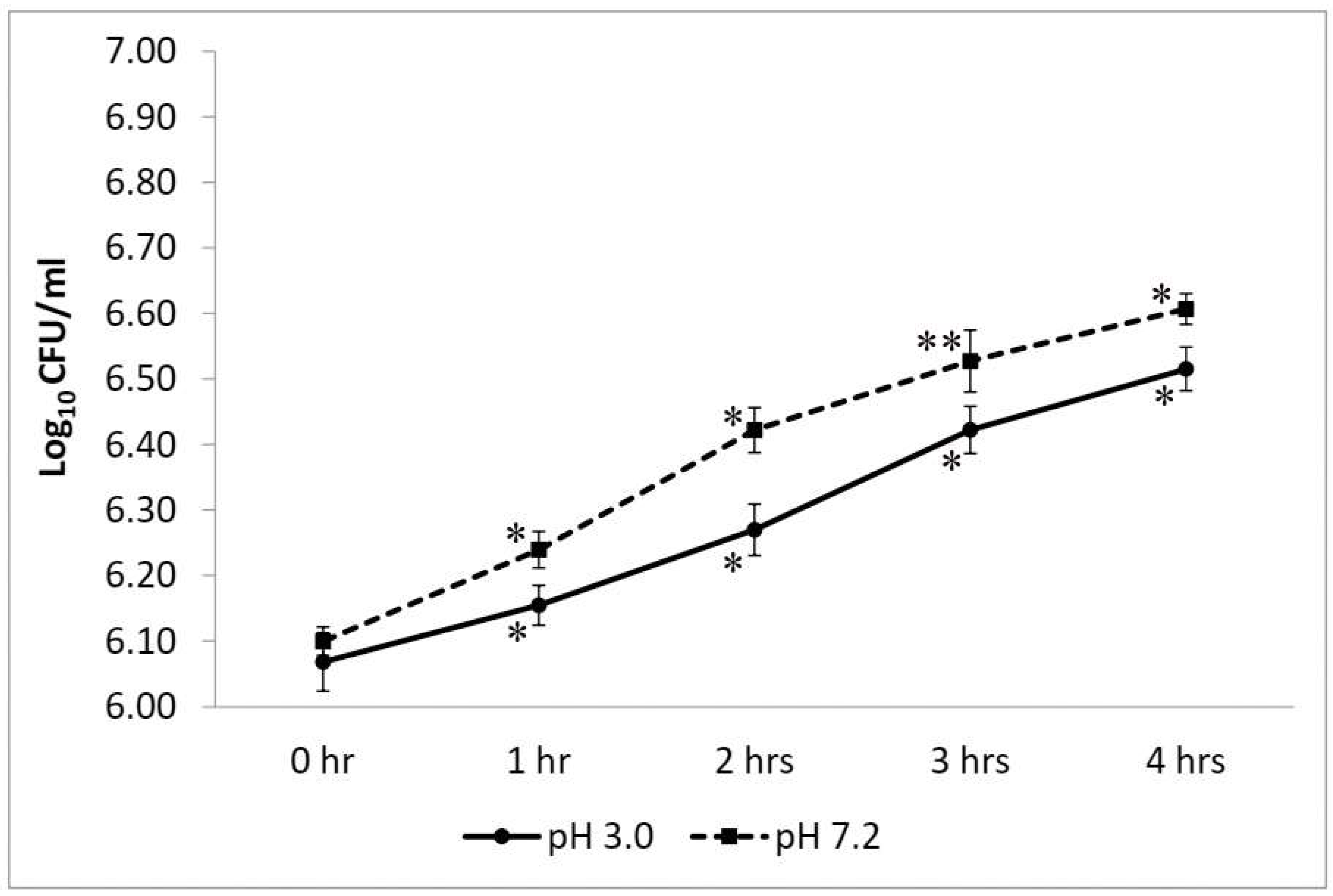
2.2.1. Test for Resistance to Low pH
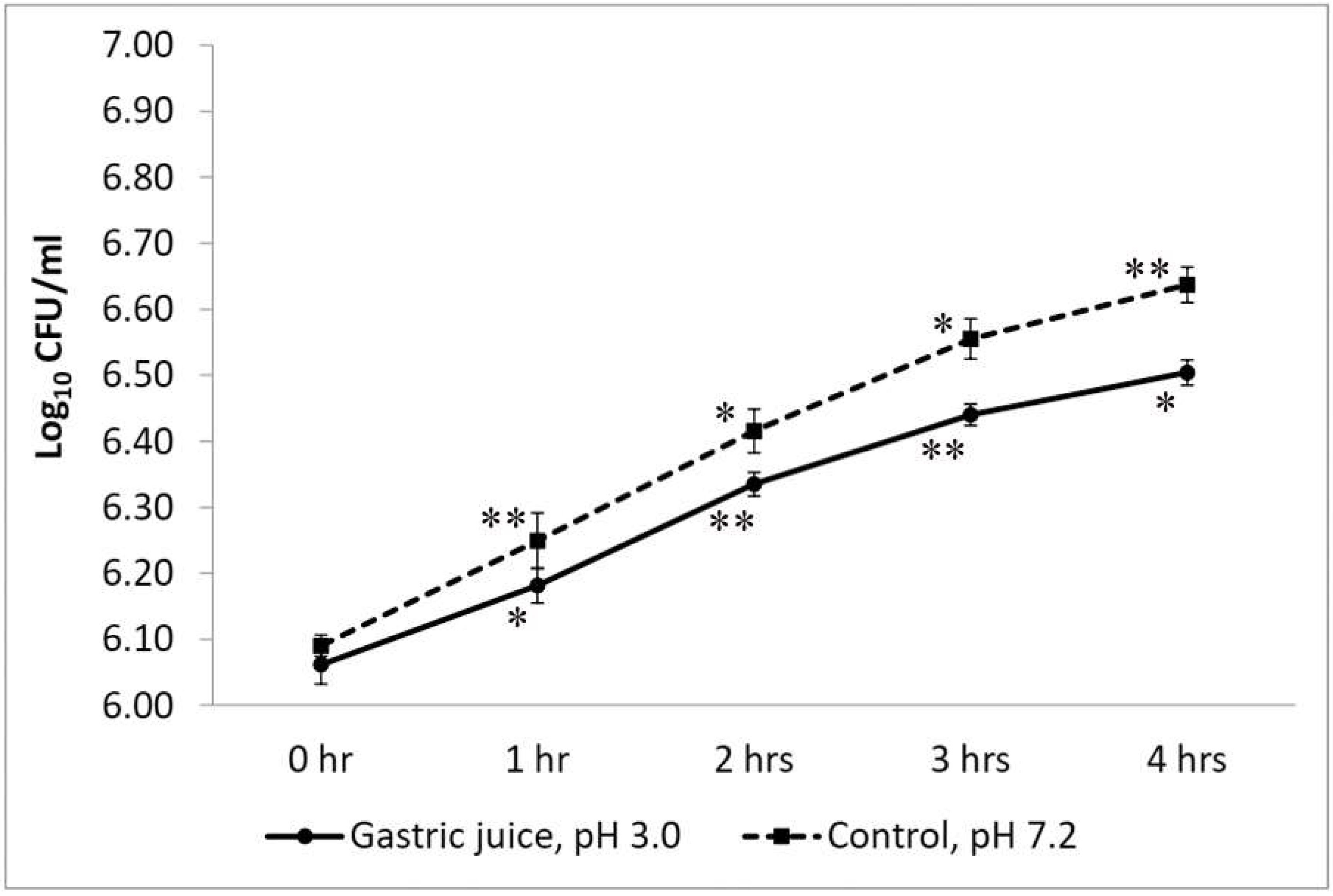
2.2.2. Simulated Gastric Juice Tolerance Test
2.2.3. Bile Tolerance Test
2.2.4. Pancreatin Tolerance Test
2.2.5. Lysozyme Tolerance
2.2.6. Cell Surface Hydrophobicity
2.2.7. Cellular Autoaggregation
2.2.8. NaCl Tolerance Test
2.3. Safety Assessment
2.3.1. Haemolytic Activity
2.3.2. DNase Activity
2.3.3. Antibiotic Resistance
2.3.4. Extraction of Antibacterial Agents and Evaluation of Their Antagonistic Activity
2.4. Evaluation of Technological Properties
2.4.1. Proteolytic Activity
2.4.2. Lipolytic Activity
2.4.3. Amylolytic Activity
2.5. Statistical Analysis
3. Results
3.1. Identification and Safe Deposit of Bacteria
3.1.1. Morphological and Biochemical Characteristics
3.1.2. Molecular Characterisation and Phylogenetic Analysis
3.1.3. Safe Deposit and Accession Number
3.2. Evaluation of Probable Probiotic Properties
3.2.1. Test for Resistance to Low pH
3.2.2. Simulated Gastric Juice Tolerance Test
3.2.3. Bile Tolerance Test
3.2.4. Pancreatin Tolerance Test
3.2.5. Lysozyme Tolerance Test
3.2.6. Cell Surface Hydrophobicity
3.2.7. Cellular Autoaggregation
3.2.8. NaCl Tolerance Test
3.3. Safety Assessment
3.3.1. Haemolytic Activity
3.3.2. DNase Activity
3.3.3. Antibiotic Sensitivity and Resistance Pattern
3.3.4. Extraction of Antibacterial Agents and Evaluation of Their Antagonistic Activity
3.4. Evaluation of Technological Properties
3.4.1. Proteolytic Activity
3.4.2. Lipolytic Activity
3.4.3. Amylolytic Activity
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bergey, D.H.; Breed, R.S.; Murray, E.G.D.; Hitchens, A.P. Manual of Determinative Bacteriology, 5th ed.; Bailliere, Tindall & Cox: London, UK, 1939. [Google Scholar]
- Breed, B.R.; Murray, E.G.D.; Smith, N.R. Bergey’s. Manual of Determinative Bacteriology, 7th ed.; Williams and Wilkins Co.: Baltimore, MD, USA, 1957. [Google Scholar]
- Gupta, R.S.; Patel, S.; Saini, N.; Chen, S. Robust demarcation of 17 distinct Bacillus species clades, proposed as novel Bacillaceae genera, by phylogenomics and comparative genomic analyses: Description of Robertmurraya kyonggiensis sp. nov. and proposal for an emended genus Bacillus limiting it only to the members of the Subtilis and Cereus clades of species. Int. J. Syst. Evol. Microbiol. 2020, 70, 5753–5798. [Google Scholar]
- Shudong, P.; Guo, C.; Wu, S.; Cui, H.; Suo, H.; Duan, Z. Bioactivity and metabolomics changes of plant-based drink fermented by Bacillus coagulans VHProbi C08. LWT 2022, 156, 113030. [Google Scholar] [CrossRef]
- Lee, B.; Lee, H.; Jeong, D.-W.; Lee, J.-H. A rapid isolation method for Bacillus coagulans from Rice Straw. Microbiol. Biotechnol. Lett. 2015, 43, 401–404. [Google Scholar] [CrossRef]
- Konuray, G.; Erginkaya, Z. Potential use of Bacillus coagulans in the food industry. Foods 2018, 7, 92. [Google Scholar] [CrossRef] [PubMed]
- Bang, W.Y.; Ban, O.-H.; Lee, B.S.; Oh, S.; Park, C.; Park, M.-K.; Jung, S.K.; Yang, J.; Jung, Y.H. Genomic-, phenotypic-, and toxicity-based safety assessment and probiotic potency of Bacillus coagulans IDCC 1201 isolated from green malt. J. Ind. Microbiol. Biotechnol. 2021, 48, kuab026. [Google Scholar] [CrossRef] [PubMed]
- Majeed, M.; Nagabhushanam, K.; Arumugam, S.; Majeed, S.; Ali, F. Bacillus coagulans MTCC 5856 for the management of major depression with irritable bowel syndrome: A randomised, double-blind, placebo controlled, multi-centre, pilot clinical study. Food Nutr. Res. 2018, 62, 1218. [Google Scholar] [CrossRef]
- Majeed, M.; Nagabhushanam, K.; Natarajan, S.; Sivakumar, A.; Ali, F.; Pande, A.; Majeed, S.; Karri, S.K. Bacillus coagulans MTCC 5856 supplementation in the management of diarrhea predominant Irritable Bowel Syndrome: A double blind randomized placebo controlled pilot clinical study. Nutr. J. 2016, 15, 21. [Google Scholar] [CrossRef]
- Hun, L. Bacillus coagulans significantly improved abdominal pain and bloating in patients with IBS. Postgrad. Med. 2009, 121, 119–124. [Google Scholar] [CrossRef]
- Maity, C.; Gupta, A.K.; Saroj, D.B.; Biyani, A.; Bagkar, P.; Kulkarni, J.; Kulkarni, J. Impact of a gastrointestinal stable probiotic supplement Bacillus coagulans LBSC on human gut microbiome modulation. J. Diet. Suppl. 2021, 18, 577–596. [Google Scholar] [CrossRef]
- Jäger, R.; Shields, K.A.; Lowery, R.P.; De Souza, E.O.; Partl, J.M.; Hollmer, C.; Purpura, M.; Wilson, J.M. Probiotic Bacillus coagulans GBI-30, 6086 reduces exercise-induced muscle damage and increases recovery. PeerJ 2016, 4, e2276. [Google Scholar] [CrossRef]
- Anaya-Loyola, M.A.; Enciso-Moreno, J.A.; López-Ramos, J.E.; García-Marín, G.; Orozco Álvarez, M.Y.; Vega-García, A.M.; Mosqueda, J.; García-Gutiérrez, D.G.; Keller, D.; Pérez-Ramírez, I.F. Bacillus coagulans GBI-30, 6068 decreases upper respiratory and gastrointestinal tract symptoms in healthy Mexican scholar-aged children by modulating immune-related proteins. Food Res. Int. 2019, 125, 108567. [Google Scholar] [CrossRef] [PubMed]
- Ratna Sudha, M.; Yelikar, K.A.; Deshpande, S. Clinical study of Bacillus coagulans unique IS-2 (ATCC PTA-11748) in the treatment of patients with bacterial vaginosis. Indian J. Microbiol. 2012, 52, 396–399. [Google Scholar] [CrossRef] [PubMed]
- Dolati, M.; Tafvizi, F.; Salehipour, M.; Movahed, T.K.; Jafari, P. Inhibitory effects of probiotic Bacillus coagulans against MCF7 breast cancer cells. Iran. J. Microbiol. 2021, 13, 839–847. [Google Scholar] [CrossRef]
- Baron, M. A patented strain of Bacillus coagulans increased immune response to viral challenge. Postgrad. Med. 2009, 121, 114–118. [Google Scholar] [CrossRef]
- Majeed, M.; Nagabhushanam, K.; Natarajan, S.; Arumugam, S.; Pande, A.; Majeed, S.; Ali, F. A double-blind, placebo-controlled, parallel study evaluating the safety of Bacillus coagulans MTCC 5856 in healthy individuals. J. Clin. Toxicol. 2016, 6, 1000283. [Google Scholar] [CrossRef]
- Tamang, J.P.; Shin, D.-H.; Jung, S.-J.; Chae, S.-W. Functional properties of microorganisms in fermented foods. Front. Microbiol. 2016, 7, 578. [Google Scholar] [CrossRef]
- Cao, J.; Yu, Z.; Liu, W.; Zhao, J.; Zhang, H.; Zhai, Q.; Chen, W. Probiotic characteristics of Bacillus coagulans and associated implications for human health and diseases. J. Funct. Foods 2020, 64, 103643. [Google Scholar] [CrossRef]
- Keller, D.; Verbruggen, S.; Cash, H.; Farmer, S.; Venema, K. Spores of Bacillus coagulans GBI-30, 6086 show high germination, survival and enzyme activity in a dynamic, computer-controlled in vitro model of the gastrointestinal tract. Benef. Microbes 2019, 10, 77–87. [Google Scholar] [CrossRef] [PubMed]
- Schillinger, U.; Guigas, C.; Holzapfel, W.H. In vitro adherence and other properties of lactobacilli used in probiotic yoghurt-like products. Int. Dairy J. 2005, 15, 1289–1297. [Google Scholar] [CrossRef]
- Salvetti, E.; Orrù, L.; Capozzi, V.; Martina, A.; Lamontanara, A.; Keller, D.; Cash, H.; Felis, G.E.; Cattivelli, L.; Torriani, S.; et al. Integrate genome-based assessment of safety for probiotic strains: Bacillus coagulans GBI-30, 6086 as a case study. Appl. Microbiol. Biotechnol. 2016, 100, 4595–4605. [Google Scholar] [CrossRef]
- Nath, S.; Roy, M.; Sikidar, J.; Deb, B.; Sharma, I.; Guha, A. Characterization and in-vitro screening of probiotic potential of novel Weissella confusa strain GCC_19R1 isolated from fermented sour rice. Curr. Res. Biotechnol. 2021, 3, 99–108. [Google Scholar] [CrossRef]
- Khagwal, N.; Sharma, P.K.; Sharma, D.C. Screening and evaluation of Lactobacillus spp. for the development of potential probiotics. Afr. J. Microbiol. Res. 2014, 8, 1573–1579. [Google Scholar]
- Bora, P.S.; Puri, V.; Bansal, A.K. Physicochemical properties and excipient compatibility studies of probiotic Bacillus coagulans spores. Sci. Pharm. 2009, 77, 625–638. [Google Scholar] [CrossRef]
- Cappuccino, J.G.; Sherman, N. Microbiology: A Laboratory Manual; Pearson: London, UK, 1996. [Google Scholar]
- Green, M.R.; Sambrook, J. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2012; p. 1890. [Google Scholar]
- Nath, S.; Deb, B.; Sharma, I. Isolation of toxic metal-tolerant bacteria from soil and examination of their bioaugmentation potentiality by pot studies in cadmium-and lead-contaminated soil. Int. Microbiol. 2018, 21, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Archer, A.C.; Halami, P.M. Probiotic attributes of Lactobacillus fermentum isolated from human feces and dairy products. Appl. Microbiol. Biotechnol. 2015, 99, 8113–8123. [Google Scholar] [CrossRef] [PubMed]
- Nath, S.; Sikidar, J.; Roy, M.; Deb, B. In vitro screening of probiotic properties of Lactobacillus plantarum isolated from fermented milk product. Food Qual. Saf. 2020, 4, 213–223. [Google Scholar] [CrossRef]
- Kõll, P.; Mändar, R.; Smidt, I.; Hütt, P.; Truusalu, K.; Mikelsaar, R.-H.; Shchepetova, J.; Krogh-Andersen, K.; Marcotte, H.; Hammarström, L.; et al. Screening and evaluation of human intestinal lactobacilli for the development of novel gastrointestinal probiotics. Curr. Microbiol. 2010, 61, 560–566. [Google Scholar] [CrossRef]
- Shivangi, S.; Devi, P.B.; Ragul, K.; Shetty, P.H. Probiotic potential of Bacillus strains isolated from an acidic fermented food Idli. Probiotics Antimicrob. Proteins 2020, 12, 1502–1513. [Google Scholar] [CrossRef]
- Pan, W.-H.; Li, P.-L.; Liu, Z. The correlation between surface hydrophobicity and adherence of Bifidobacterium strains from centenarians’ faeces. Anaerobe 2006, 12, 148–152. [Google Scholar] [CrossRef]
- Shangpliang, H.N.J.; Sharma, S.; Rai, R.; Tamang, J.P. Some technological properties of lactic acid bacteria isolated from Dahi and Datshi, naturally fermented milk products of Bhutan. Front. Microbiol. 2017, 8, 116. [Google Scholar] [CrossRef]
- Xu, H.; Jeong, H.S.; Lee, H.Y.; Ahn, J. Assessment of cell surface properties and adhesion potential of selected probiotic strains. Lett. Appl. Microbiol. 2009, 49, 434–442. [Google Scholar] [CrossRef] [PubMed]
- Rastogi, S.; Mittal, V.; Singh, A. In vitro evaluation of probiotic potential and safety assessment of Lactobacillus mucosae strains isolated from Donkey’s lactation. Probiotics Antimicrob. Proteins 2019, 12, 1045–1056. [Google Scholar] [CrossRef]
- Baccer, R.; Kirby, W.M.; Kirby, W.M.; Turck, M. Antibiotic susceptibility testing by standard single disc diffusion method. Am. J. Clin. Pathol. 1966, 45, 493–496. [Google Scholar]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing; Clinical Lab Standards Institute: Wayne, PA, USA, 2021; Volume 31. [Google Scholar]
- Hussein, E.I.; Jacob, J.H.; Shakhatreh, M.A.K.; Al-Razaq, M.A.A.; Juhmani, A.-S.F.; Cornelison, C.T. Detection of antibiotic-producing Actinobacteria in the sediment and water of Ma’in thermal springs (Jordan). Germs 2018, 8, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Nath, S.; Sinha, A.; Singha, Y.S.; Dey, A.; Bhattacharjee, N.; Deb, B. Prevalence of antibiotic-resistant, toxic metal-tolerant and biofilm-forming bacteria in hospital surroundings. Environ. Anal. Health Toxicol. 2020, 35, e2020018. [Google Scholar] [CrossRef] [PubMed]
- Raveschot, C.; Cudennec, B.; Deracinois, B.; Frémont, M.; Vaeremans, M.; Dugersuren, J.; Demberel, S.; Drider, D.; Dhulster, P.; Coutte, F.; et al. Proteolytic activity of Lactobacillus strains isolated from Mongolian traditional dairy products: A multiparametric analysis. Food Chem. 2020, 304, 125415. [Google Scholar] [CrossRef] [PubMed]
- Aspri, M.; Bozoudi, D.; Tsaltas, D.; Hill., C.; Papademas., P. Raw donkey milk as a source of Enterococcus diversity: Assessment of their technological properties and safety characteristics. Food Control. 2017, 73, 8–90. [Google Scholar] [CrossRef]
- do Espirito-Santo, A.P.; Mouquet-Rivier, C.; Humblot, C.; Cazevieille, C.; Icard-Vernière, C.; Soccol, C.R.; Guyot, J.-P. Influence of cofermentation by amylolytic Lactobacillus strains and probiotic bacteria on the fermentation process, viscosity and microstructure of gruels made of rice, soy milk and passion fruit fiber. Food Res. Int. 2014, 57, 104–113. [Google Scholar] [CrossRef]
- Abdhul, K.; Ganesh, M.; Shanmughapriya, S.; Vanithamani, S.; Kanagavel, M.; Anbarasu, K.; Natarajaseenivasan, K. Bacteriocinogenic potential of a probiotic strain Bacillus coagulans [BDU3] from Ngari. Int. J. Biol. Macromol. 2015, 79, 800–806. [Google Scholar] [CrossRef]
- Altun, G.K.; Erginkaya, Z. Identification and characterization of Bacillus coagulans strains for probiotic activity and safety. LWT 2021, 151, 112233. [Google Scholar] [CrossRef]
- Singhal, N.; Singh, N.S.; Mohanty, S.; Singh, P.; Virdi, J.S. Evaluation of Probiotic Characteristics of Lactic Acid Bacteria Isolated from Two Commercial Preparations Available in Indian Market. Indian J. Microbiol. 2019, 59, 112–115. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Kandasamy, S.; Kavitake, D.; Shetty, P.H. Probiotic characterization and antioxidant properties of Weissella confusa KR780676, isolated from an Indian fermented food. LWT 2018, 97, 53–60. [Google Scholar] [CrossRef]
- Mortuza, T. Isolation and Identification of Microbes from Various Fruit Juices Made and Sold for Immediate Consumption at Home and in the Market of Dhaka City; BRAC University: Dhaka, Bangladesh, 2016. [Google Scholar]
- Shinde, T.; Vemuri, R.; Shastri, M.D.; Perera, A.P.; Tristram, S.; Stanley, R.; Eri, R. Probiotic Bacillus coagulans MTCC 5856 spores exhibit excellent in-vitro functional efficacy in simulated gastric survival, mucosal adhesion and immunomodulation. J. Funct. Foods 2019, 52, 100–108. [Google Scholar] [CrossRef]
- Ruiz, L.; Margolles, A.; Sánchez, B. Bile resistance mechanisms in Lactobacillus and Bifidobacterium. Front. Microbiol. 2013, 4, 396. [Google Scholar] [CrossRef] [PubMed]
- Bernet-Camard, M.F.; Coconnier, M.H.; Hudault, S.; Servin, A.L. Differentiation-associated antimicrobial functions in human colon adenocarcinoma cell lines. Exp. Cell Res. 1996, 226, 80–89. [Google Scholar] [CrossRef]
- Ragul, K.; Syiem, I.; Sundar, K.; Shetty, P.H. Characterization of probiotic potential of Bacillus species isolated from a traditional brine pickle. J. Food Sci. Technol. 2017, 54, 4473–4483. [Google Scholar] [CrossRef]
- de Souza, B.M.S.; Borgonovi, T.F.; Casarotti, S.; Todorov, S. Lactobacillus casei and Lactobacillus fermentum strains isolated from mozzarella cheese: Probiotic potential, safety, acidifying kinetic parameters and viability under gastrointestinal tract conditions. Probiotics Antimicrob. Proteins 2019, 11, 382–396. [Google Scholar] [CrossRef]
- Shakirova, L.; Grube, M.; Gavare, M.; Auzina, L.; Zikmanis, P. Lactobacillus acidophilus La5 and Bifidobacterium lactis Bb12 cell surface hydrophobicity and survival of the cells under adverse environmental conditions. J. Ind. Microbiol. Biotechnol. 2013, 40, 85–93. [Google Scholar] [CrossRef]
- Krausova, G.; Hyrslova, I.; Hynstova, I. In vitro evaluation of adhesion capacity, hydrophobicity, and auto-aggregation of newly isolated potential probiotic strains. Fermentation 2019, 5, 100. [Google Scholar] [CrossRef]
- Soni, R.; Keharia, H.; Dunlap, C.; Pandit, N.; Doshi, J. Functional annotation unravels probiotic properties of a poultry isolate, Bacillus velezensis CGS1. 1. LWT 2022, 153, 112471. [Google Scholar] [CrossRef]
- Anandharaj, M.; Sivasankari, B. Isolation of potential probiotic Lactobacillus oris HMI68 from mother’s milk with cholesterol-reducing property. J. Biosci. 2014, 118, 153–159. [Google Scholar] [CrossRef] [PubMed]
- Saroj, D.B.; Gupta, A.K. Genome based safety assessment for Bacillus coagulans strain LBSC (DSM 17654) for probiotic application. Int. J. Food Microbiol. 2020, 318, 108523. [Google Scholar] [CrossRef]
- Sui, L.; Zhu, X.; Wu, D.; Ma, T.; Tuo, Y.; Jiang, S.; Qian, F.; Mu, G. In vitro assessment of probiotic and functional properties of Bacillus coagulans T242. Food Biosci. 2020, 36, 100675. [Google Scholar] [CrossRef]
- Kim, Y.-S.; Lee, J.; Heo, S.; Lee, J.-H.; Jeong, D.-W. Technology and safety evaluation of Bacillus coagulans exhibiting antimicrobial activity for starter development. LWT 2021, 137, 110464. [Google Scholar] [CrossRef]
- Choudhury, P.; Bhunia, B. Industrial application of lipase: A review. Biopharm. J. 2015, 1, 41–47. [Google Scholar]
- Ray, A. Application of lipase in industry. Asian J. Pharm. Technol. 2012, 2, 33–37. [Google Scholar]
- Wang, Y.; Cao, W.; Luo, J.; Qi, B.; Wan, Y. One step open fermentation for lactic acid production from inedible starchy biomass by thermophilic Bacillus coagulans IPE22. Bioresour. Technol. 2019, 272, 398–406. [Google Scholar] [CrossRef]
- Bailon-Salas, A.M.; Ordaz-Díaz, L.A.; Valle-Cervantes, S.; López-Miranda, J.; Urtiz-Estrada, U.; Páez-Lerma, J.B.; Rojas-Contreras, J.A. Characterization of Culturable Bacteria from Pulp and Paper Industry Wastewater, with the Potential for Degradation of Cellulose, Starch, and Lipids. BioResources 2018, 13, 5052–5064. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Observations |
|---|---|
| General Characteristics | |
| MRS broth | Turbid |
| MRS agar | Elevated, small-medium-sized colonies |
| Colony colour | White and creamy |
| Pigmentation | No |
| Growth temperature | 40 °C |
| Growth condition | Facultative anaerobe |
| Cell arrangement | Single or in-chain |
| Cell shape | Bacillus |
| Gram staining | Gram-positive |
| Endospore test | Positive |
| Biochemical Characteristics | |
| Indole test | Positive |
| Methyl red test | Positive |
| Voges-Proskauer test | Positive |
| Citrate utilisation test | Negative |
| Catalase Test | Positive |
| Oxidase test | Positive |
| Starch hydrolysis | Positive |
| Sl. No. | Antibiotic | Concentration (in µg) | Zone of Inhibition (in mm) |
|---|---|---|---|
| 1. | Polymyxin-B | PB-300 | 30 |
| 2. | Amoxyclav | AMC-30 | 42 |
| 3. | Rifampicin | RIF-5 | 35 |
| 4. | Tetracycline | TE-30 | 44 |
| 5. | Oxacillin | OX-5 | 35 |
| 6. | Amikacin | AK-30 | 27 |
| 7. | Cefoxitin | CX-30 | 31 |
| 8. | Cefepime | CPM-30 | 33 |
| 9 | Ceftazidime | CAZ-30 | 24 |
| 10. | Cefotaxime | CTX-30 | 25 |
| 11. | Chloramphenicol | C-30 | 34 |
| 12. | Cefdinir | CDR-5 | 37 |
| 13. | Penicillin g | P-10 | 18 |
| 14. | Moxifloxacin | MO-5 | 35 |
| 15. | Ampicillin | AMP-10 | 19 |
| 16. | Vancomycin | VA-30 | 22 |
| 17. | Ceftriaxone | CTR-30 | 30 |
| 18. | Neomycin | N-10 | 24 |
| 19. | Ofloxacin | OF-5 | 35 |
| 20. | Norfloxacin | NX-10 | 33 |
| 21. | Kanamycin | K-30 | 30 |
| 22. | Bacitracin | B-10 | 24 |
| 23. | Co-Trimoxazole | COT-25 | 13 |
| 24. | Methicillin | MET-10 | 37 |
| 25. | Streptomycin | S-10 | 22 |
| 26. | Levofloxacin | LE-5 | 35 |
| 27. | Erythromycin | E-15 | 11 |
| 28. | Clindamycin | CD-2 | 29 |
| 29. | Gentamycin | HLG-120 | 36 |
| 30. | Sterile disc | Control | 0 |
| Sl. No. | Test Pathogens | Zone of Inhibition (in mm) |
|---|---|---|
| 1. | Staphylococcus aureus strain GCC_20MS | 21 |
| 2. | Mammaliicoccus sciuri strain GCC_20RS | 18 |
| 3 | Bacillus cereus strain GCC_21R1 | 10 |
| 4. | Bacillus nealsonii strain GCC_21R8 | 14 |
| 5. | Bacillus megaterium strain GCC-SO1 | 14 |
| 6. | Enterobacter bugandensis strain GCC_21R10 | 18 |
| 7. | Pseudomonas aeruginosa strain GCC_19W1 | 20 |
| 8. | Stenotrophomonas maltophilia strain GCC_19W2 | 18 |
| 9. | Achromobacter spanius strain GCCSB1 | 20 |
| 10. | Acinetobacter johnsonii strain SB_SK | 17 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sreenadh, M.; Kumar, K.R.; Nath, S. In Vitro Evaluation of Weizmannia coagulans Strain LMG S-31876 Isolated from Fermented Rice for Potential Probiotic Properties, Safety Assessment and Technological Properties. Life 2022, 12, 1388. https://doi.org/10.3390/life12091388
Sreenadh M, Kumar KR, Nath S. In Vitro Evaluation of Weizmannia coagulans Strain LMG S-31876 Isolated from Fermented Rice for Potential Probiotic Properties, Safety Assessment and Technological Properties. Life. 2022; 12(9):1388. https://doi.org/10.3390/life12091388
Chicago/Turabian StyleSreenadh, Madapati, Kallur Ranjith Kumar, and Soumitra Nath. 2022. "In Vitro Evaluation of Weizmannia coagulans Strain LMG S-31876 Isolated from Fermented Rice for Potential Probiotic Properties, Safety Assessment and Technological Properties" Life 12, no. 9: 1388. https://doi.org/10.3390/life12091388
APA StyleSreenadh, M., Kumar, K. R., & Nath, S. (2022). In Vitro Evaluation of Weizmannia coagulans Strain LMG S-31876 Isolated from Fermented Rice for Potential Probiotic Properties, Safety Assessment and Technological Properties. Life, 12(9), 1388. https://doi.org/10.3390/life12091388