
A Review with Updated Perspectives on the Antiviral Potentials of Traditional Medicinal Plants and Their Prospects in Antiviral Therapy

 , , , , , , and
, , , , , , and
Abstract
1. Introduction
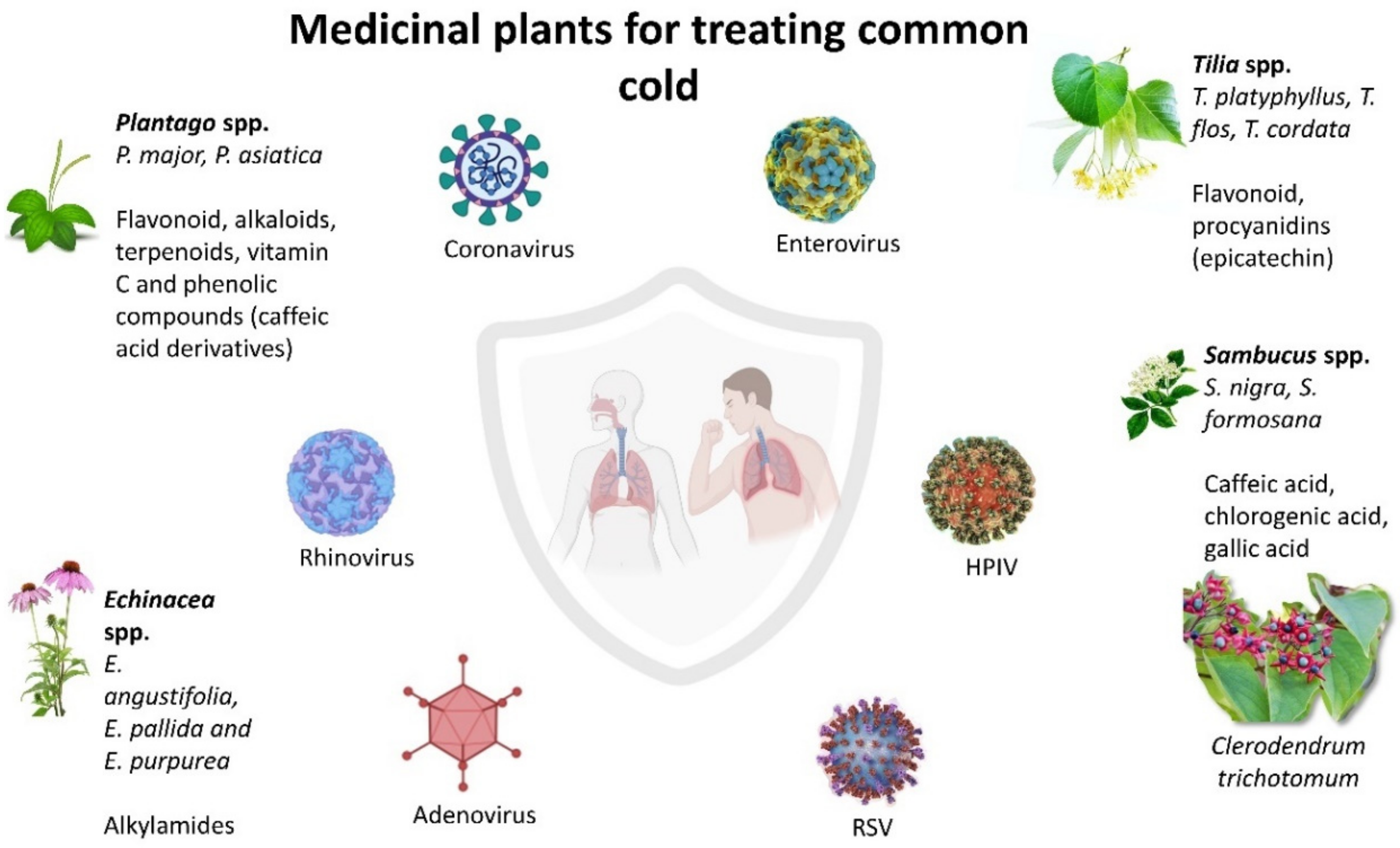
2. Medicinal Plants for Treating Common Cold
2.1. Echinacea spp.
2.2. Sambucus spp.
2.3. Plantago spp.
2.4. Tilia spp.
2.5. Clerodendrum trichotomum
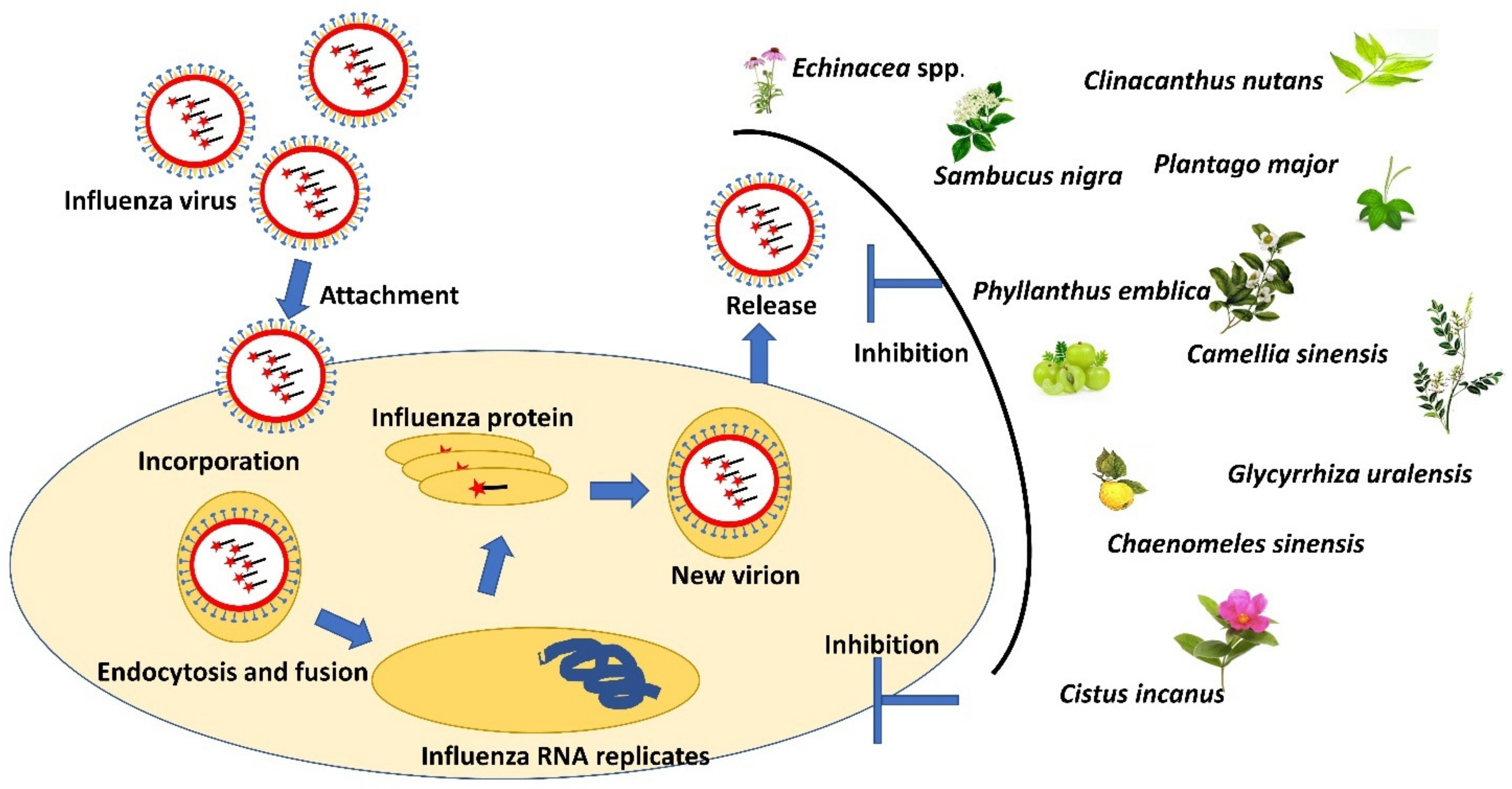
3. Medicinal Plant for Treating Influenza
3.1. Punica granatum
3.2. Geranium sanguineum
3.3. Echinacea spp.
3.4. Cistus incanus
3.5. Glycyrrhiza spp.
3.6. Chaenomeles sinensis
3.7. Sambucus nigra
3.8. Phyllanthus emblica
3.9. Camellia sinensis (Green Tea)
4. Medicinal Plants for Antiviral Treatment of Sexually Transmitted Diseases
4.1. Clinacanthus nutans and Clinacanthus siamensis
4.2. Cistus incanus
4.3. Plantago major
4.4. Polygonum minus
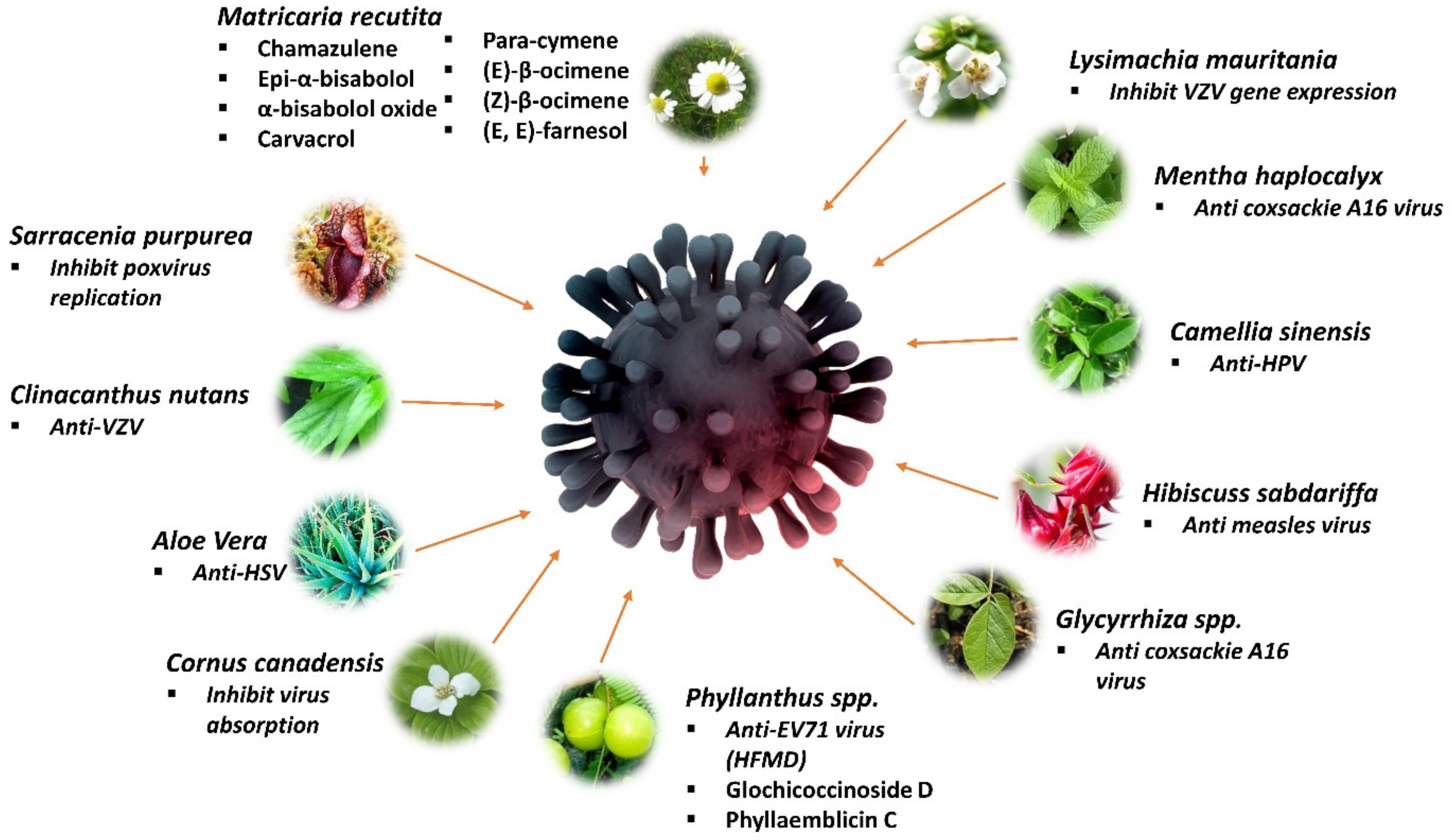
5. Medicinal Plant for Treating Dermatology-Related Viruses
5.1. Sarracenia purpurea
5.2. Clinacanthus nutans (Lindau)
5.3. Matricaria recutita
5.4. Aloe vera
5.5. Cornus spp.
5.6. Lysimachia mauritiana
5.7. Mentha haplocalyx
5.8. Camellia sinensis
5.9. Hibiscuss sabdariffa
5.10. Glycyrrhiza glabra and Glycyrrhiza uralensis
5.11. Phyllanthus spp.
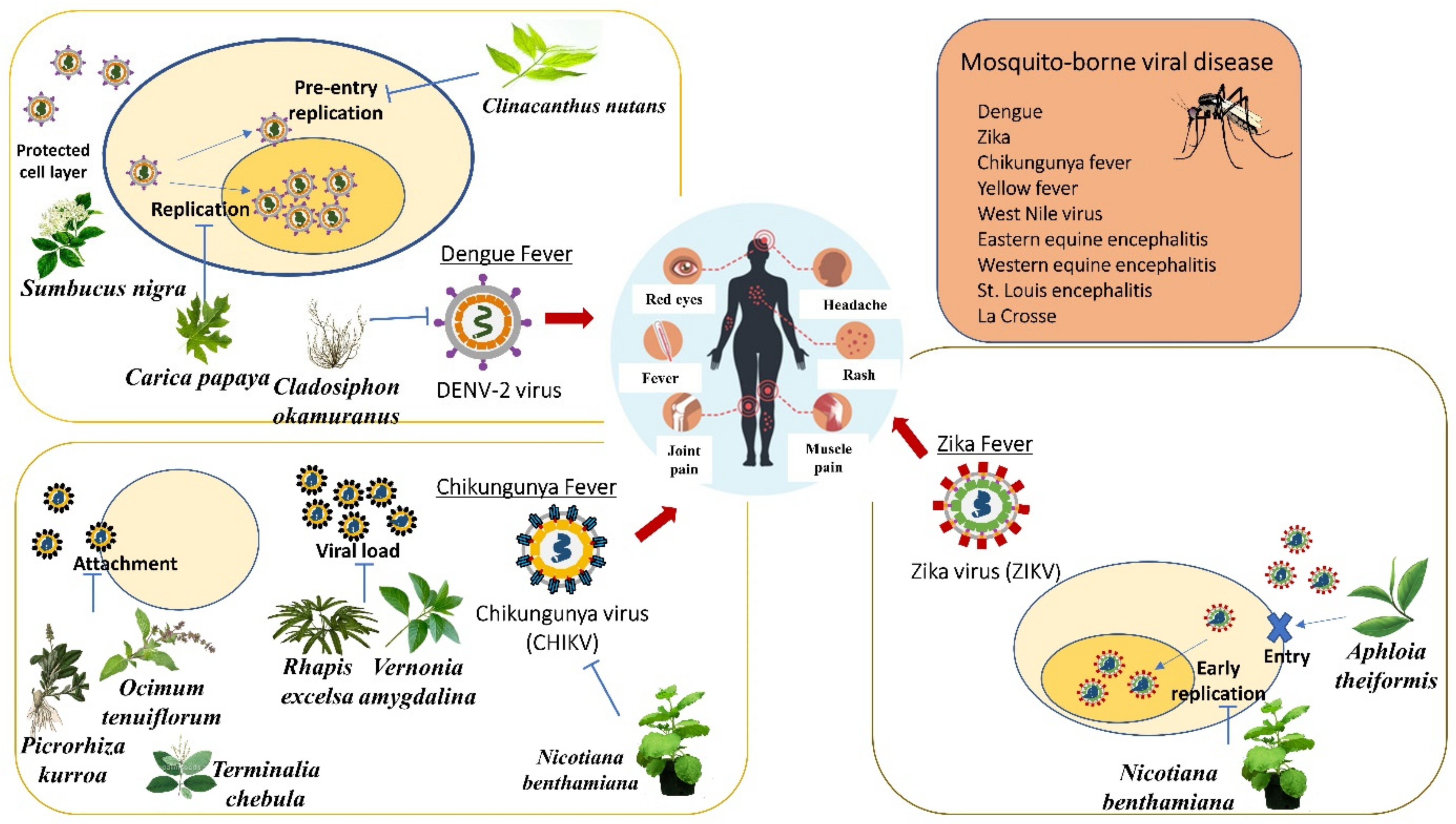
6. Medicinal Plants for Treating Mosquito-Borne Viral Disease
6.1. Anti-Dengue Medicinal Plants
6.1.1. Alternanthera philoxeroids
6.1.2. Cladosiphon okamuranus
6.1.3. Cladogynos orientalis
6.1.4. Clinacanthus nutans
6.1.5. Sambucus nigra
6.1.6. Carica papaya
6.2. Anti-Chikungunya Medicinal Plants
6.2.1. Picrorhiza kurroa, Ocimum tenuiflorum, and Terminalia chebula
6.2.2. Rhapis excelsa and Vernonia amygdalina
6.2.3. Nicotiana benthamiana
6.3. Anti-Zika Medicinal Plants
6.3.1. Aphloia theiformis
6.3.2. Nicotiana benthamiana
6.3.3. Psiloxylon mauritianum
7. Discussion
8. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| IC50 | 50% inhibitory concentration |
| EC50 | Half maximal effective concentration |
| CC50 | 50% Cytotoxic concentration |
| ED50 | Median effective dose |
| TD50 | Median toxic dose |
| MIC100 | Minimum inhibitory concentration at 100 µm |
| MOI | Multiplicity of infection |
References
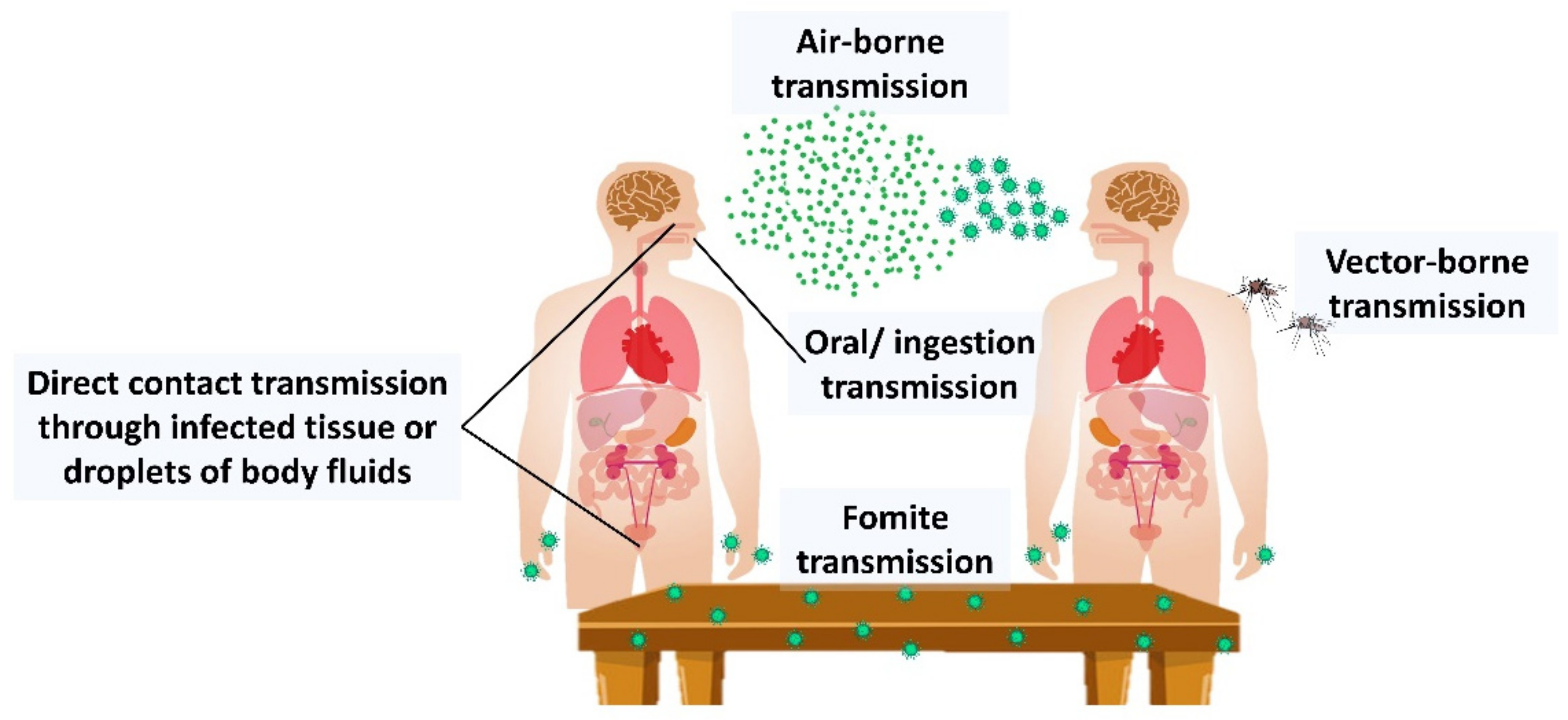
- La Rosa, G.; Fratini, M.; Libera, S.D.; Iaconelli, M.; Muscillo, M. Viral infections acquired indoors through airborne, droplet or contact transmission. Ann. Dell’istituto Super. Sanita 2013, 49, 124–132. [Google Scholar]
- Nadaf, M.; Joharchi, M.; Amiri, M.S. Ethnomedicinal uses of plants for the treatment of nervous disorders at the herbal markets of Bojnord, North Khorasan Province, Iran. Avicenna J. Phytomedicine 2019, 9, 153. [Google Scholar]
- Sousa, S.G.; Oliveira, L.A.; Magalhães, D.A.; de Brito, T.A.; Batista, J.A.; Pereira, C.M.C.; Costa, M.S.; Mazulo, J.C.R.; Filgueiras, M.C.; Vasconselos, D.F.P.; et al. Chemical structure and anti-inflammatory effect of polysaccharide extracted from Morinda citrifolia Linn (Noni). Carbohydr. Polym. 2018, 197, 515–523. [Google Scholar] [CrossRef]
- Petrovska, B.B. Historical review of medicinal plants’ usage. Pharmacogn. Rev. 2012, 6, 1. [Google Scholar] [CrossRef]
- Mazid, M.; Khan, T.A.; Mohammad, F. Medicinal plants of rural India: A review of use by Indian folks. Indo Glob. J. Pharm. Sci. 2012, 2, 286–304. [Google Scholar]
- Jassim, S.A.; Naji, M.A. Novel antiviral agents: A medicinal plant perspective. J. Appl. Microbiol. 2003, 95, 412–427. [Google Scholar] [CrossRef]
- Azwanida, N. A review on the extraction methods use in medicinal plants, principle, strength and limitation. Med. Aromat Plants 2015, 4, 1000196. [Google Scholar] [CrossRef]
- Divya, M.; Vijayakumar, S.; Chen, J.; Vaseeharan, B.; Esteban, F.; Durán-Larac, D. South Indian medicinal plants can combat deadly viruses along with COVID-19?—A review. Microb. Pathog. 2020, 148, 104277. [Google Scholar] [CrossRef]
- Musidlak, O.; Nawrot, R.; Goździcka-Józefiak, A. Which plant proteins are involved in antiviral defense? Review on in vivo and in vitro activities of selected plant proteins against viruses. Int. J. Mol. Sci. 2017, 18, 2300. [Google Scholar] [CrossRef]
- Stollenwerk, N.; Harper, R.W.; Sandrock, C.E. Bench-to-bedside review: Rare and common viral infections in the intensive care unit–linking pathophysiology to clinical presentation. Crit. Care 2008, 12, 219. [Google Scholar] [CrossRef][Green Version]
- Burrell, C.J.; Howard, C.R.; Murphy, F.A. Fenner and White’s Medical Virology; Academic Press: Cambridge, MA, USA, 2016. [Google Scholar]
- Heagy, W.; Crumpacker, C.; Lopez, P.A.; Finberg, R.W. Inhibition of immune functions by antiviral drugs. J. Clin. Investig. 1991, 87, 1916–1924. [Google Scholar] [CrossRef]
- Richman, D.D. Antiviral drug resistance. Antivir. Res. 2006, 71, 117–121. [Google Scholar] [CrossRef] [PubMed]
- Warren, T.; Harris, J.; Brennan, C.A. Efficacy and safety of valacyclovir for the suppression and episodic treatment of herpes simplex virus in patients with HIV. Clin. Infect. Dis. 2004, 39 (Suppl. 5), S258–S266. [Google Scholar] [CrossRef] [PubMed]
- Razonable, R.R. Antiviral drugs for Viruses Other than Human Immunodeficiency Virus. In Mayo Clinic Proceedings; Elsevier: Amsterdam, The Netherlands, 2011. [Google Scholar]
- Paintsil, E.; Cheng, Y.-C. Antiviral agents. Encycl. Microbiol. 2019, 176–225. [Google Scholar] [CrossRef]
- Mondal, D. Valganciclovir. In xPharm: The Comprehensive Pharmacology Reference; Enna, S.J., Bylund, D.B., Eds.; Elsevier: New York, NY, USA, 2007; pp. 1–4. [Google Scholar]
- Pinto, L.H. Understanding the mechanism of action of the anti-influenza virus drug amantadine. Thends Microbiol. 1995, 3, 271. [Google Scholar] [CrossRef]
- Papich, M.G.; Davis, J.L. Antimicrobial therapy. In Equine Infectious Diseases; Elsevier: St. Louis, MO, USA, 2007; pp. 578–591. [Google Scholar]
- Anderson, P.L.; Rower, J.E. Zidovudine and Lamivudine for HIV Infection. Clin. Med. Rev. Ther. 2010, 2, a2004. [Google Scholar]
- Berk, L.; Schalm, S.W.; Heijtink, R.A. Zidovudine inhibits hepatitis B virus replication. Antivir. Res. 1992, 19, 111–118. [Google Scholar] [CrossRef]
- Malin, J.J.; Suárez, I.; Priesner, V.; Fätkenheuer, G.; Rybniker, J. Remdesivir against COVID-19 and Other Viral Diseases. Clin. Microbiol. Rev. 2020, 34, e00162-20. [Google Scholar] [CrossRef]
- Ahmad, B.; Batool, M.; Ain, Q.; Moon Suk Kim, M.S.; Choi, S. Exploring the binding mechanism of PF-07321332 SARS-CoV-2 protease inhibitor through molecular dynamics and binding free energy simulations. Int. J. Mol. Sci. 2021, 22, 9124. [Google Scholar] [CrossRef]
- Wat, D. The common cold: A review of the literature. Eur. J. Intern. Med. 2004, 15, 79–88. [Google Scholar] [CrossRef]
- Delfan, B.; Kazemeini, H.; Bahmani, M. Identifying Effective Medicinal Plants for Cold in Lorestan Province, West of Iran. J. Evidence-Based Integr. Med. 2015, 20, 173–179. [Google Scholar] [CrossRef]
- Greenberg, S.B. Update on rhinovirus and coronavirus infections. In Seminars in Respiratory and Critical Care Medicine; Thieme Medical Publishers: New York, NY, USA, 2011. [Google Scholar]
- Su, S.; Wong, G.; Shi, W.; Liu, J.; Lai, A.C.K.; Zhou, J.; Liu, W.; Bi, Y.; Gao, G.F. Epidemiology, Genetic Recombination, and Pathogenesis of Coronaviruses. Trends Microbiol. 2016, 24, 490–502. [Google Scholar] [CrossRef]
- Branche, A.R.; Falsey, A.R. Parainfluenza virus infection. In Seminars in Respiratory and Critical Care Medicine; Thieme Medical Publishers: New York, NY, USA, 2016. [Google Scholar]
- Lugo, D.; Krogstad, P. Enteroviruses in the early 21st century: New manifestations and challenges. Curr. Opin. Pediatrics 2016, 28, 107. [Google Scholar] [CrossRef] [PubMed]
- Chiru, T.; Fursenco, C.; Ciobanu, N.; Dinu, M.; Popescu, E.; Ancuceanu, R.; Volmer, D.; Raal, A. Use of medicinal plants in complementary treatment of the common cold and influenza–perception of pharmacy customers in Moldova and Romania. J. Herb. Med. 2020, 21, 100346. [Google Scholar] [CrossRef]
- Hudson, J.; Vimalanathan, S. Echinacea—A Source of Potent Antivirals for Respiratory Virus Infections. Pharmaceuticals 2011, 4, 1019–1031. [Google Scholar] [CrossRef]
- Signer, J.; Jonsdottir, H.R.; Albrich, W.C.; Strasser, M.; Züst, R.; Ryter, S.; Ackermann-Gäumann, R.; Lenz, N.; Denise Siegrist, D.; Suter, A.; et al. Engler In vitro virucidal activity of Echinaforce®, an Echinacea purpurea preparation, against coronaviruses, including common cold coronavirus 229E and SARS-CoV-2. Virol. J. 2020, 17, 136. [Google Scholar] [CrossRef] [PubMed]
- Weng, J.-R.; Lin, C.-S.; Lai, H.-C.; Lin, Y.-P.; Wang, C.-Y.; Tsai, Y.-C.; Wu, K.-C.; Huang, S.-H.; Lin, C.-W. Antiviral activity of Sambucus FormosanaNakai ethanol extract and related phenolic acid constituents against human coronavirus NL63. Virus Res. 2019, 273, 197767. [Google Scholar] [CrossRef]
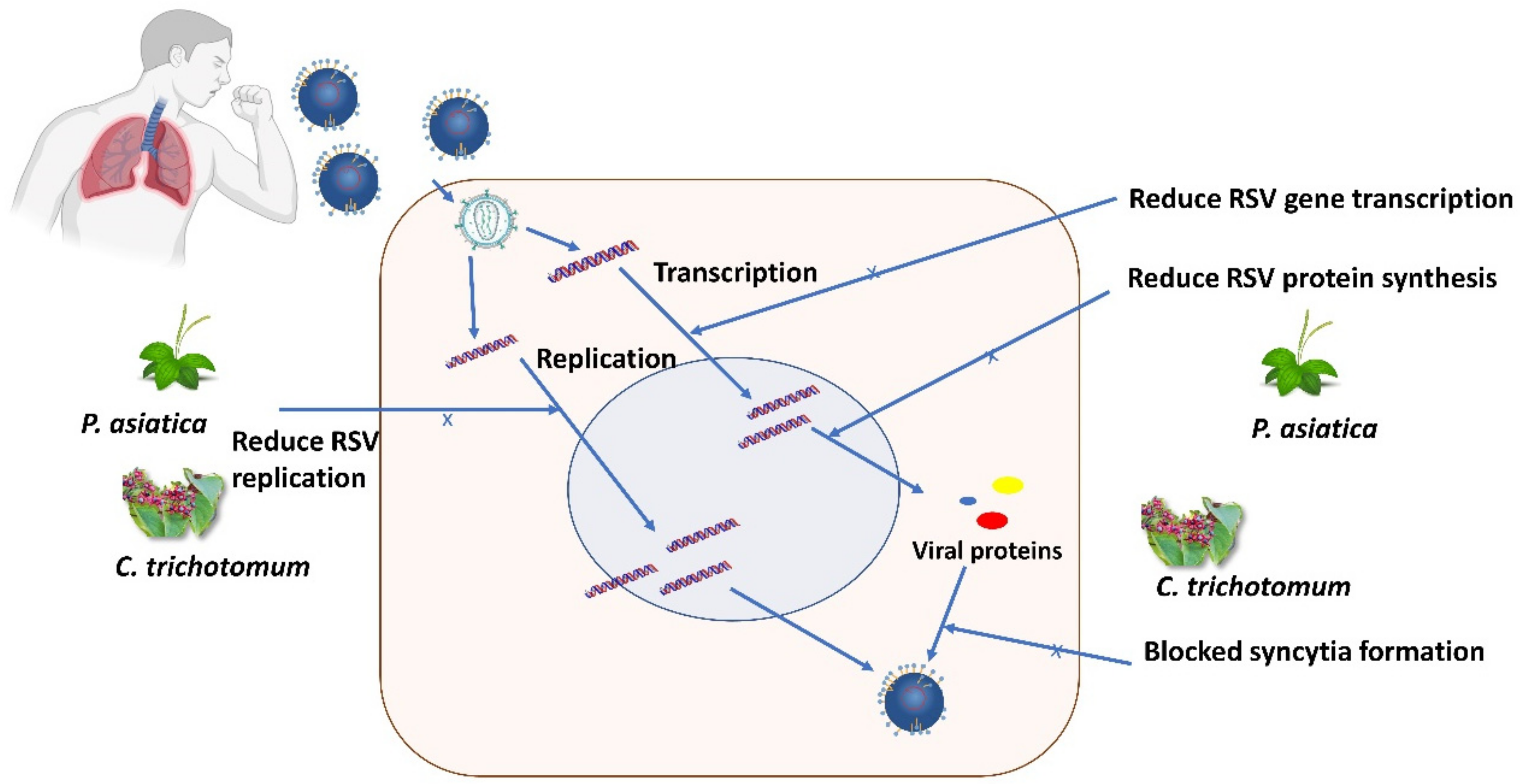
- Chathuranga, K.; Kim, M.S.; Lee, H.-C.; Kim, T.-H.; Kim, J.-H.; Gayan Chathuranga, W.A.; Ekanayaka, P.; Wijerathne, H.M.S.M.; Cho, W.-K.; Kim, H.; et al. Anti-Respiratory Syncytial Virus Activity of Plantago asiatica and Clerodendrum trichotomum Extracts In Vitro and In Vivo. Viruses 2019, 11, 604. [Google Scholar] [CrossRef]
- Eldin, S.M.S.; Shawky, E.; Sallam, S.M.; El-Nikhely, N.; El Sohafy, S.M. Metabolomics approach provides new insights into the immunomodulatory discriminatory biomarkers of the herbs and roots of Echinacea species. Ind. Crop. Prod. 2021, 168, 113611. [Google Scholar] [CrossRef]
- Sperber, S.J.; Shah, L.P.; Gilbert, R.D.; Ritchey, T.W.; Monto, A.S. Echinacea purpurea for Prevention of Experimental Rhinovirus Colds. Clin. Infect. Dis. 2004, 38, 1367–1371. [Google Scholar] [CrossRef]
- Hudson, J.; Vimalanathan, S.; Kang, L.; Amiguet, V.T.; Livesey, J.; Arnason, J.T. Characterization of antiviral activities in Echinacea. Root preparations. Pharm. Biol. 2005, 43, 790–796. [Google Scholar] [CrossRef]
- Młynarczyk, K.; Walkowiak-Tomczak, D.; Łysiak, G. Bioactive properties of Sambucus nigra L. as a functional ingredient for food and pharmaceutical industry. J. Funct. Foods 2018, 40, 377–390. [Google Scholar] [CrossRef] [PubMed]
- Mahboubi, M. Sambucus nigra (black elder) as alternative treatment for cold and flu. Adv. Tradit. Med. 2021, 21, 405–414. [Google Scholar] [CrossRef]
- Chen, C.; Zuckerman, D.; Brantley, S.; Sharpe, M.; Hoiczyk, E.; Pendleton, A. Sambucus nigra Extracts Inhibit Infectious Bronchitis Virus at an Early Point during Replication. Planta Medica 2013, 79, PN83. [Google Scholar] [CrossRef]
- Haddadian, K.; Haddadian, K.; Zahmatkash, M. A Review of Plantago Plant; NISCAIR-CSIR: New Delhi, India, 2014. [Google Scholar]
- Parsaei, P.; Bahmani, M.; Naghdi, N.; Asadi-Samani, M.; Rafieian-Kopaei, M.; Tajeddini, P.; Sepehri-Boroujeni, M. Identification of medicinal plants effective on common cold: An ethnobotanical study of Shiraz, South Iran. Der Pharm. Lett. 2016, 8, 90–97. [Google Scholar]
- Chiang, L.; Chiang, W.; Chang, M.; Ng, L.; Lin, C. Antiviral activity of Plantago major extracts and related compounds in vitro. Antivir. Res. 2002, 55, 53–62. [Google Scholar] [CrossRef]
- Eaton, E.; Caudullo, G.; de Rigo, D. Tilia Cordata, Tilia Platyphyllos and Other Limes in Europe: Distribution, Habitat, Usage and Threats. In European Atlas of Forest Tree Species; Publications Office of the European Union: Luxemburg, 2016; p. e010ec5. [Google Scholar]
- Fitsiou, L.; Tzakou, O.; Hancianu, M.; Poiata, A. Volatile Constituents and Antimicrobial Activity of Tilia tomentosa Moench and Tilia cordata Miller Oils. J. Essent. Oil Res. 2007, 19, 183–185. [Google Scholar] [CrossRef]
- Vogl, S.; Picker, P.; Mihaly-Bison, J.; Fakhrudin, N.; Atanasov, A.G.; Heiss, E.H.; Wawrosch, C.; Reznicek, G.; Dirsch, V.M.; Saukel, J.; et al. Ethnopharmacological in vitro studies on Austria’s folk medicine—An unexplored lore in vitro anti-inflammatory activities of 71 Austrian traditional herbal drugs. J. Ethnopharmacol. 2013, 149, 750–771. [Google Scholar] [CrossRef]
- Czerwińska, M.E.; Dudek, M.K.; Pawłowska, K.A.; Pruś, A.; Ziaja, M.; Granica, S. The influence of procyanidins isolated from small-leaved lime flowers (Tilia cordata Mill.) on human neutrophils. Fitoterapia 2018, 127, 115–122. [Google Scholar] [CrossRef]
- Xu, R.; Wang, R.; Ha, W.; Shi, Y.-P. New cyclohexylethanoids from the leaves of Clerodendrum trichotomum. Phytochem. Lett. 2014, 7, 111–113. [Google Scholar] [CrossRef]
- Long, J.S.; Mistry, B.; Haslam, S.M.; Barclay, W.S. Host and viral determinants of influenza A virus species specificity. Nat. Rev. Genet. 2019, 17, 67–81. [Google Scholar] [CrossRef]
- Moghadami, M. A Narrative Review of Influenza: A Seasonal and Pandemic Disease. Iran. J. Med. Sci. 2017, 42, 2. [Google Scholar]
- Yahaya, R.; Dash, G.K.; Abdullah, M.S.; Mathews, A. Clinacanthus nutans (burm. F.) Lindau: An useful medicinal plant of south-east Asia. Int. J. Pharmacogn. Phytochem. Res. 2015, 7, 1244–1250. [Google Scholar]
- Tran, T.T.; Kim, M.; Jang, Y.; Lee, H.W.; Nguyen, H.T.; Nguyen, T.N.; Park, H.W.; Le Dang, Q.; Kim, J.-C. Characterization and mechanisms of anti-influenza virus metabolites isolated from the Vietnamese medicinal plant Polygonum chinense. BMC Complement. Altern. Med. 2017, 17, 162. [Google Scholar] [CrossRef] [PubMed]
- Rajasekaran, D.; Palombo, E.A.; Yeo, T.C.; Ley, D.L.S.; Tu, C.L.; Malherbe, F.; Grollo, L. Identification of Traditional Medicinal Plant Extracts with Novel Anti-Influenza Activity. PLoS ONE 2013, 8, e79293. [Google Scholar] [CrossRef]
- Moradi, M.-T.; Karimi, A.; Shahrani, M.; Hashemi, L.; Ghaffari-Goosheh, M.-S. Anti-Influenza Virus Activity and Phenolic Content of Pomegranate (Punica granatum L.) Peel Extract and Fractions. Avicenna J. Med. Biotechnol. 2019, 11, 285. [Google Scholar] [PubMed]
- Haidari, M.; Ali, M.; Casscells, S.W.; Madjid, M. Pomegranate (Punica granatum) purified polyphenol extract inhibits influenza virus and has a synergistic effect with oseltamivir. Phytomedicine 2009, 16, 1127–1136. [Google Scholar] [CrossRef] [PubMed]
- Sundararajan, A.; Ganapathy, R.; Huan, L.; Dunlap, J.R.; Webby, R.J.; Kotwal, G.J.; Sangster, M.Y. Influenza virus variation in susceptibility to inactivation by pomegranate polyphenols is determined by envelope glycoproteins. Antivir. Res. 2010, 88, 1–9. [Google Scholar] [CrossRef]
- Pantev, A.; Ivancheva, S.; Staneva, L.; Serkedjieva, J. Biologically Active Constituents of a Polyphenol Extract from Geranium sanguineum L. with Anti-Influenza Activity. Z. Für Nat. C 2006, 61, 508–516. [Google Scholar] [CrossRef]
- Serkedjieva, J.; Hay, A.J. In vitro anti-influenza virus activity of a plant preparation from Geranium sanguineum L. Antivir. Res. 1998, 37, 121–130. [Google Scholar] [CrossRef]
- Droebner, K.; Ehrhardt, C.; Poetter, A.; Ludwig, S.; Planz, O. CYSTUS052, a polyphenol-rich plant extract, exerts anti-influenza virus activity in mice. Antivir. Res. 2007, 76, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Ehrhardt, C.; Hrincius, E.R.; Korte, V.; Mazur, I.; Droebner, K.; Poetter, A.; Dreschers, S.; Schmolke, M.; Planz, O.; Ludwig, S. A polyphenol rich plant extract, CYSTUS052, exerts anti influenza virus activity in cell culture without toxic side effects or the tendency to induce viral resistance. Antivir. Res. 2007, 76, 38–47. [Google Scholar] [CrossRef]
- Michaelis, M.; Geiler, J.; Naczk, P.; Sithisarn, P.; Ogbomo, H.; Altenbrandt, B.; Leutz, A.; Doerr, H.W.; Cinatl, J. Glycyrrhizin inhibits highly pathogenic H5N1 influenza A virus-induced pro-inflammatory cytokine and chemokine expression in human macrophages. Med. Microbiol. Immunol. 2010, 199, 291–297. [Google Scholar] [CrossRef] [PubMed]
- Sawai, R.; Kuroda, K.; Shibata, T.; Gomyou, R.; Osawa, K.; Shimizu, K. Anti-influenza virus activity of Chaenomeles sinensis. J. Ethnopharmacol. 2008, 118, 108–112. [Google Scholar] [CrossRef] [PubMed]
- Sawai-Kuroda, R.; Kikuchi, S.; Shimizu, Y.K.; Sasaki, Y.; Kuroda, K.; Tanaka, T.; Yamamoto, T.; Sakurai, K.; Shimizu, K. A polyphenol-rich extract from Chaenomeles sinensis (Chinese quince) inhibits influenza A virus infection by preventing primary transcription in vitro. J. Ethnopharmacol. 2013, 146, 866–872. [Google Scholar] [CrossRef] [PubMed]
- Torabian, G.; Valtchev, P.; Adil, Q.; Dehghani, F. Anti-influenza activity of elderberry (Sambucus nigra). J. Funct. Foods 2019, 54, 353–360. [Google Scholar] [CrossRef]
- Liu, G.; Xiong, S.; Xiang, Y.-F.; Guo, C.-W.; Ge, F.; Yang, C.-R.; Zhang, Y.-J.; Wang, Y.-F.; Kitazato, K. Antiviral activity and possible mechanisms of action of pentagalloylglucose (PGG) against influenza A virus. Arch. Virol. 2011, 156, 1359–1369. [Google Scholar] [CrossRef]
- Lv, J.J.; Yu, S.; Xin, Y.; Cheng, R.R.; Zhu, H.T.; Wang, D.; Yang, C.R.; Xu, M.; Zhang, Y.J. Anti-viral and cytotoxic norbisabolane sesquiterpenoid glycosides from Phyllanthus emblica and their absolute configurations. Phytochemistry 2015, 117, 123–134. [Google Scholar] [CrossRef]
- Song, J.-M.; Lee, K.-H.; Seong, B.-L. Antiviral effect of catechins in green tea on influenza virus. Antivir. Res. 2005, 68, 66–74. [Google Scholar] [CrossRef]
- Vimalanathan, S.; Schoop, R.; Hudson, J. High-potency anti-influenza therapy by a combination of Echinacea purpurea fresh herb and root tinctures. J. Appl. Pharm. Sci. 2013, 3, 001–005. [Google Scholar]
- Viapiana, A.; Konopacka, A.; Waleron, K.; Wesolowski, M. Cistus incanus L. commercial products as a good source of polyphenols in human diet. Ind. Crop. Prod. 2017, 107, 297–304. [Google Scholar] [CrossRef]
- Rebensburg, S.; Helfer, M.; Schneider, M.; Koppensteiner, H.; Eberle, J.; Schindler, M.; Gürtler, L.; Brack-Werner, R. Potent in vitro antiviral activity of Cistus incanus extract against HIV and Filoviruses targets viral envelope proteins. Sci. Rep. 2016, 6, 20394. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Liang, J.; Zhang, J.; Wang, Y.; Chai, X. Natural Chalcones in Chinese Materia Medica: Licorice. Evid. Based Complement. Altern. Med. 2020, 2020, 821248. [Google Scholar] [CrossRef]
- Gomaa, A.A.; Abdel-Wadood, Y.A. The potential of glycyrrhizin and licorice extract in combating COVID-19 and associated conditions. Phytomed. Plus 2021, 1, 100043. [Google Scholar] [CrossRef] [PubMed]
- Cinatl, J.; Morgenstern, B.; Bauer, G.; Chandra, P.; Rabenau, H.; Doerr, H. Glycyrrhizin, an active component of liquorice roots, and replication of SARS-associated coronavirus. Lancet 2003, 361, 2045–2046. [Google Scholar] [CrossRef]
- Chen, F. In vitro susceptibility of 10 clinical isolates of SARS coronavirus to selected antiviral compounds. J. Clin. Virol. 2004, 31, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Moisy, D.; Avilov, S.V.; Jacob, Y.; Laoide, B.M.; Ge, X.; Baudin, F.; Naffakh, N.; Jestin, J.-L. HMGB1 Protein Binds to Influenza Virus Nucleoprotein and Promotes Viral Replication. J. Virol. 2012, 86, 9122–9133. [Google Scholar] [CrossRef] [PubMed]
- Fal, A.M.; Conrad, F.; Schönknecht, K.; Sievers, H.; Pawińska, A. Antiviral activity of the Virus Blocking Factor (VBF) derived i.a. from Pelargonium extract and Sambucus juice against different human-pathogenic cold viruses in vitro. Wiad. Lek. 2016, 69 Pt 2, 499–511. [Google Scholar]
- Lee, Y.H.; Jang, Y.H.; Kim, Y.-S.; Kim, J.; Seong, B.L. Evaluation of green tea extract as a safe personal hygiene against viral infections. J. Biol. Eng. 2018, 12, 1. [Google Scholar] [CrossRef]
- Low, N.; Broutet, N.; Turner, R. A Collection on the Prevention, Diagnosis, and Treatment of Sexually Transmitted Infections: Call for Research Papers; Public Library of Science: San Francisco, CA, USA, 2017; p. e1002333. [Google Scholar]
- Denny, L.A.; Franceschi, S.; de Sanjosé, S.; Heard, I.; Moscicki, A.B.; Palefsky, J. Human Papillomavirus, Human Immunodeficiency Virus and Immunosuppression. Vaccine 2012, 30, F168–F174. [Google Scholar] [CrossRef]
- Gheit, T. Mucosal and Cutaneous Human Papillomavirus Infections and Cancer Biology. Front. Oncol. 2019, 9, 355. [Google Scholar] [CrossRef] [PubMed]
- Somia, I.K.A.; Teeratakulpisarn, N.; Jeo, W.S.; Yee, I.A.; Pankam, T.; Nonenoy, S.; Trachuntong, D.; Mingkwanrungrueng, P.; Sukmawati, M.D.D.; Ramautarsing, R.; et al. Prevalence of and risk factors for anal high-risk HPV among HIV-negative and HIV-positive MSM and transgender women in three countries at South-East Asia. Medicine 2018, 97, e9898. [Google Scholar] [CrossRef] [PubMed]
- Sharp, P.M.; Hahn, B.H. Origins of HIV and the AIDS pandemic. Cold Spring Harb. Perspect. Med. 2011, 1, a006841. [Google Scholar] [CrossRef] [PubMed]
- Bazazi, A.R.; Crawford, F.; Zelenev, A.; Heimer, R.; Kamarulzaman, A.; Altice, F.L. HIV prevalence among people who inject drugs in greater Kuala Lumpur recruited using respondent-driven sampling. AIDS Behav. 2015, 19, 2347–2357. [Google Scholar] [CrossRef] [PubMed]
- Vergara-Ortega, D.N.; Sevilla-Reyes, E.E.; Herrera-Ortiz, A.; Torres-Ibarra, L.; Salmerón, J.; Lazcano-Ponce, E.; Sánchez-Alemán, M.A. Real time PCR to evaluate HSV-2 shedding from anal and genital samples among men who have sex with men, living with HIV. J. Med Virol. 2018, 90, 745–752. [Google Scholar] [CrossRef]
- Benjamin, M.M.; Gummelt, K.L.; Zaki, R.; Afzal, A.; Sloan, L.; Shamim, S. Herpes simplex virus meningitis complicated by ascending paralysis. In Baylor University Medical Center Proceedings; Taylor & Franci: Oxfordshire, UK, 2013. [Google Scholar]
- Olayinka, A.T.; Nasidi, A.; Gidado, S.; Sha’Aibu, S.; Balogun, M.S.; Oyemakinde, A.; Nguku, P.; Ajudua, A.; Egwuenu-Oladejo, A.; Musa, B.O.P.; et al. Seroprevalence of Hepatitis B Infection in Nigeria: A National Survey. Am. J. Trop. Med. Hyg. 2016, 95, 902. [Google Scholar] [CrossRef]
- Kunsorn, P.; Ruangrungsi, N.; Lipipun, V.; Khanboon, A.; Rungsihirunrat, K. The identities and anti-herpes simplex virus activity of Clinacanthus nutans and Clinacanthus siamensis. Asian Pac. J. Trop. Biomed. 2013, 3, 284–290. [Google Scholar] [CrossRef]
- Wahab, N.Z.A.; Bunawan, H.; Ibrahim, N. Cytotoxicity and antiviral activity of methanol extract from Polygonum minus. In Proceedings of the AIP Conference Proceedings, Selangor, Malaysia, 15–16 April 2015; AIP Publishing LLC: Melville, NY, USA, 2015. [Google Scholar]
- Brianti, P.; De Flammineis, E.; Mercuri, S.R. Review of HPV-related diseases and cancers. New Microbiol. 2017, 40, 85. [Google Scholar]
- Gershon, A.A.; Breuer, J.; Cohen, J.I.; Cohrs, R.J.; Gershon, M.D.; Gilden, D.; Grose, C.; Hambleton, S.; Kennedy, P.G.; Oxman, M.N.; et al. Varicella zoster virus infection. Nat. Rev. Dis. Primers 2015, 1, 15016. [Google Scholar] [CrossRef]
- Mueller, N.H.; Gilden, D.H.; Cohrs, R.J.; Mahalingam, R.; Nagel, M.A. Varicella Zoster Virus Infection: Clinical Features, Molecular Pathogenesis of Disease, and Latency. Neurol. Clin. 2008, 26, 675–697. [Google Scholar] [CrossRef]
- Phyu, W.K.; Ong, K.C.; Wong, K.T. A consistent orally-infected hamster model for enterovirus A71 encephalomyelitis demonstrates squamous lesions in the paws, skin and oral cavity reminiscent of hand-foot-and-mouth disease. PLoS ONE 2016, 11, e0147463. [Google Scholar] [CrossRef] [PubMed]
- Lerch, M.; Mainetti, C.; Beretta-Piccoli, B.T.; Harr, T. Current Perspectives on Erythema Multiforme. Clin. Rev. Allergy Immunol. 2018, 54, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Sokumbi, O.; Wetter, D.A. Clinical features, diagnosis, and treatment of erythema multiforme: A review for the practicing dermatologist. Int. J. Dermatol. 2012, 51, 889–902. [Google Scholar] [CrossRef] [PubMed]
- De Wet, H.; Nciki, S.; van Vuuren, S.F. Medicinal plants used for the treatment of various skin disorders by a rural community in northern Maputaland, South Africa. J. Ethnobiol. Ethnomed. 2013, 9, 51. [Google Scholar] [CrossRef]
- Arndt, W.; Mitnik, C.; Denzler, K.L.; White, S.; Waters, R.; Jacobs, B.L.; Rochon, Y.; Olson, V.A.; Damon, I.K.; Langland, J.O. In Vitro Characterization of a Nineteenth-Century Therapy for Smallpox. PLoS ONE 2012, 7, e32610. [Google Scholar] [CrossRef]
- Khoo, L.W.; Kow, S.A.; Lee, M.T.; Tan, C.P.; Shaari, K.; Tham, C.L.; Abas, F. A Comprehensive Review on Phytochemistry and Pharmacological Activities of Clinacanthus nutans (Burm.f.) Lindau. Evid.-Based Complement. Altern. Med. 2018, 2018, 9276260. [Google Scholar] [CrossRef]
- Charuwichitratana, S.; Wongrattanapasson, N.; Timpatanapong, P.; Bunjob, M. Herpes Zoster: Treatment with Clinacanthus Nutans Cream. Int. J. Dermatol. 1996, 35, 665–666. [Google Scholar] [CrossRef]
- Koch, C.; Reichling, J.; Schneele, J.; Schnitzler, P. Inhibitory effect of essential oils against herpes simplex virus type 2. Phytomedicine 2008, 15, 71–78. [Google Scholar] [CrossRef]
- Rezazadeh, F.; Moshaverinia, M.; Motamedifar, M.; Alyaseri, M. Assessment of Anti HSV-1 Activity of Aloe Vera Gel Extract: An In Vitro Study. J. Dent. 2016, 17, 49. [Google Scholar]
- Zandi, K.; Zadeh, M.A.; Sartavi, K.; Rastian, Z. Antiviral activity of Aloe vera against herpes simplex virus type 2: An in vitro study. Afr. J. Biotechnol. 2007, 6, 15. [Google Scholar]
- Song, J.-H.; Park, K.; Shim, A.; Kwon, B.-E.; Ahn, J.-H.; Choi, Y.J.; Kim, J.K.; Yeo, S.-G.; Yoon, K.; Ko, H.-J. Complete Sequence Analysis and Antiviral Screening of Medicinal Plants for Human Coxsackievirus A16 Isolated in Korea. Osong Public Heal. Res. Perspect. 2015, 6, 52–58. [Google Scholar] [CrossRef] [PubMed]
- Bae, S.; Song, Y.-J. Inhibition of varicella-zoster virus replication by an ethanol extract of Lysimachia mauritiana. Mol. Med. Rep. 2017, 15, 3847–3851. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Wang, C.; Xu, L.; Chen, X.; Wang, W.; Yang, G.; Tan, R.X.; Li, E.; Jin, Y. A laboratory evaluation of medicinal herbs used in china for the treatment of hand, foot, and mouth disease. Evid. Based Complement. Altern. Med. 2013, 2013, 504563. [Google Scholar] [CrossRef] [PubMed]
- Zou, C.; Liu, H.; Feugang, J.M.; Hao, Z.; Chow, H.-H.S.; Garcia, F. Green Tea Compound in Chemoprevention of Cervical Cancer. Int. J. Gynecol. Cancer 2010, 20, 617–624. [Google Scholar] [CrossRef] [PubMed]
- Omilabu, A.S.; Oyefolu, O.A.B. Antiviral effect of Hibiscus sabdariffa and Celosia argentea on measles virus. Afr. J. Microbiol. Res. 2010, 4, 293–296. [Google Scholar]
- Wang, J.; Chen, X.; Wang, W.; Zhang, Y.; Yang, Z.; Jin, Y.; Ge, H.M.; Li, E.; Yang, G. Glycyrrhizic acid as the antiviral component of Glycyrrhiza uralensis Fisch. against coxsackievirus A16 and enterovirus 71 of hand foot and mouth disease. J. Ethnopharmacol. 2013, 147, 114–121. [Google Scholar] [CrossRef]
- Kuo, K.-K.; Chang, J.-S.; Wang, K.-C.; Chiang, L.-C. Water Extract of Glycyrrhiza uralensis Inhibited Enterovirus 71 in a Human Foreskin Fibroblast Cell Line. Am. J. Chin. Med. 2009, 37, 383–394. [Google Scholar] [CrossRef]
- Dah, M.V.; Beckstead, A.L.; Rheins, L.A. Treatment of herpetic cold sores with an extract of Sarracenia Purpurea. Glob. Dermatol. 2017, 4. [Google Scholar] [CrossRef]
- Kannan, L.; Kumar, A.; Kumar, A.; Jacobs, B.; Langland, J. Anti-herpes virus activity of the carnivorous botanical, Sarracenia purpurea. Sci. Rep. 2020, 10, 18953. [Google Scholar] [CrossRef]
- Aslam, M.S.; Ahmad, M.S.; Mamat, A.S.; Ahmad, M.Z.; Salam, F. Antioxidant and wound healing activity of polyherbal fractions of Clinacanthus nutans and Elephantopus scaber. Evid. Based Complement. Altern. Med. 2016, 2016, 4685246. [Google Scholar] [CrossRef]
- Lalla, R.V.; Sonis, S.T.; Peterson, D.E. Management of Oral Mucositis in Patients Who Have Cancer. Dent. Clin. North Am. 2008, 52, 61–77. [Google Scholar] [CrossRef] [PubMed]
- Dawid-Pać, R. Medicinal plants used in treatment of inflammatory skin diseases. Adv. Dermatol. Allergol./Postȩpy Dermatol. I Alergol. 2013, 30, 170. [Google Scholar] [CrossRef] [PubMed]
- Moghaddasi Mohammad, S. Study on Cammomile (Matricaria chamomilla L.) usage and Farming. Adv. Environ. Biol. 2011, A263659352, 1446–1454. [Google Scholar]
- Hekmatpou, D.; Mehrabi, F.; Rahzani, K.; Aminiyan, A. The Effect of Aloe Vera Clinical Trials on Prevention and Healing of Skin Wound: A Systematic Review. Iran. J. Med. Sci. 2019, 44, 1. [Google Scholar] [PubMed]
- Rajeswari, R.; Umadevi, M.; Rahale, C.S.; Pushpa, R.; Selvavenkadesh, S.; Kumar, K.S.; Bhowmik, D. Aloe vera: The miracle plant its medicinal and traditional uses in India. J. Pharmacogn. Phytochem. 2012, 1, 118–124. [Google Scholar]
- Lavoie, S.; Côté, I.; Pichette, A.; Gauthier, C.; Ouellet, M.; Nagau-Lavoie, F.; Mshvildadze, V.; Legault, J. Chemical composition and anti-herpes simplex virus type 1 (HSV-1) activity of extracts from Cornus canadensis. BMC Complement. Altern. Med. 2017, 17, 123. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, H.; Wei, B. Immune response of T cells during herpes simplex virus type 1 (HSV-1) infection. J. Zhejiang Univ. Sci. B 2017, 18, 277–288. [Google Scholar] [CrossRef]
- Brahmi, F.; Khodir, M.; Mohamed, C.; Pierre, D. Chemical composition and biological activities of Mentha species. Aromat. Med. Plants-Back Nat. 2017, 10, 47–79. [Google Scholar]
- Miyoshi, N.; Tanabe, H.; Suzuki, T.; Saeki, K.; Hara, Y. Applications of a Standardized Green Tea Catechin Preparation for Viral Warts and Human Papilloma Virus-Related and Unrelated Cancers. Molecules 2020, 25, 2588. [Google Scholar] [CrossRef]
- Ajoku, G.; Okwute, S.; Okogun, J. Preliminary phytochemical and antimicrobial screening of the calyx of green Hibiscus sabdariffa (Linn)(Malvaceae). J. Phytomed. Ther. 2015, 15, 41–52. [Google Scholar]
- Bhalla, A.; Thirumalaikolundusubramanian, P.; Fung, J.; Cordero-Schmidt, G.; Soghoian, S.; Sikka, V.K.; Dhindsa, H.S.; Singh, S. Native medicines and cardiovascular toxicity. In Heart and Toxins; Elsevier: Amsterdam, The Netherlands, 2015; pp. 175–202. [Google Scholar]
- Tanemoto, R.; Okuyama, T.; Matsuo, H.; Okumura, T.; Ikeya, Y.; Nishizawa, M. The constituents of licorice (Glycyrrhiza uralensis) differentially suppress nitric oxide production in interleukin-1β-treated hepatocytes. Biochem. Biophys. Rep. 2015, 2, 153–159. [Google Scholar] [CrossRef]
- Shebl, R.I.; Amin, M.A.; Emad-Eldin, A.; Bin Dajem, S.M.; Mostafa, A.S.; Ibrahim, E.H.; Mohamed, A.F. Antiviral activity of liquorice powder extract against varicella zoster virus isolated from Egyptian patients. Chang. Gung. Med. J. 2012, 35, 231–239. [Google Scholar] [PubMed]
- Sarisetyaningtyas, P.V.; Hadinegoro, S.R.; Munasir, Z. Randomized controlled trial of Phyllanthus niruri Linn extract. Paediatr. Indones. 2006, 46, 77–81. [Google Scholar] [CrossRef]
- Sarwar, M. Mosquito-Borne Viral Infections and Diseases among Persons and Interfering with the Vector Activities. Int. J. Vaccines Vaccin. 2016, 3, 63. [Google Scholar] [CrossRef]
- Jiang, W.-L.; Luo, X.-L.; Kuang, S.-J. Effects of Alternanthera philoxeroides Griseb against dengue virus in vitro. Di 1 Jun Yi Da Xue Xue Bao Acad. J. First Med. Coll. PLA 2005, 25, 454–456. [Google Scholar]
- Hidari, K.I.P.J.; Takahashi, N.; Arihara, M.; Nagaoka, M.; Morita, K.; Suzuki, T. Structure and anti-dengue virus activity of sulfated polysaccharide from a marine alga. Biochem. Biophys. Res. Commun. 2008, 376, 91–95. [Google Scholar] [CrossRef] [PubMed]
- Tu, S.-F.; Liu, R.H.; Cheng, Y.-B.; Hsu, Y.-M.; Du, Y.-C.; El-Shazly, M.; Wu, Y.-C.; Chang, F.-R. Chemical Constituents and Bioactivities of Clinacanthus nutans Aerial Parts. Molecules 2014, 19, 20382–20390. [Google Scholar] [CrossRef]
- Sakdarat, S.; Sittiso, S.; Ekalaksananan, T.; Pientong, C.; Charoensri, N.; Kongyingyoes, B. Study on Effects of Compounds from Clinacanthus Nutans on Dengue Virus Type 2 Infection. 2017. Available online: https://ssrn.com/abstract=2961652 (accessed on 29 July 2022).
- Direkbusarakom, S.; Ruangpan, L.; Ezura, Y.; Yoshimizu, M. Protective Efficacy of Clinacanthus nutans on Yellow-head Disease in Black Tiger Shrimp (Penaeus monodon). Fish Pathol. 1998, 33, 401–404. [Google Scholar] [CrossRef]
- Castillo-Maldonado, I.; Moreno-Altamirano, M.M.B.; Serrano-Gallardo, L.B. Anti-dengue serotype-2 activity effect of Sambucus nigra leaves-and flowers-derived compounds. Virol. Res. Rev. 2017, 1, 1–5. [Google Scholar] [CrossRef]
- Joseph, B.; Sankarganesh, P.; Ichiyama, K.; Yamamoto, N. In vitro study on cytotoxic effect and anti-DENV2 activity of Carica papaya L. leaf. Front. Life Sci. 2015, 8, 18–22. [Google Scholar] [CrossRef]
- Salim, F.; Abu, N.A.; Yaakob, H.; Kadir, L.; Zainol, N.; Taher, Z. Interaction of Carica papaya L. leaves optimum extract on virus dengue infected cells. Sci. Int. 2018, 30, 437–441. [Google Scholar]
- Raghavendhar, S.; Tripati, P.K.; Ray, P.; Patel, A.K. Evaluation of medicinal herbs for Anti-CHIKV activity. Virology 2019, 533, 45–49. [Google Scholar] [CrossRef] [PubMed]
- Chan, Y.S.; Khoo, K.S. Investigation of twenty selected medicinal plants from Malaysia for anti-Chikungunya virus activity. Int. Microbiol. 2016, 19, 175–182. [Google Scholar] [PubMed]
- Hurtado, J.; Acharya, D.; Lai, H.; Sun, H.; Kallolimath, S.; Steinkellner, H.; Bai, F.; Chen, Q. In vitro and in vivo efficacy of anti-chikungunya virus monoclonal antibodies produced in wild-type and glycoengineered Nicotiana benthamiana plants. Plant Biotechnol. J. 2020, 18, 266–273. [Google Scholar] [CrossRef] [PubMed]
- Clain, E.; Sinigaglia, L.; Koishi, A.C.; Gorgette, O.; Gadea, G.; Viranaicken, W.; Krejbich-Trotot, P.; Mavingui, P.; Desprès, P.; Dos Santos, C.N.D.; et al. Extract from Aphloia theiformis, an edible indigenous plant from Reunion Island, impairs Zika virus attachment to the host cell surface. Sci. Rep. 2018, 8, 10856. [Google Scholar] [CrossRef]
- Clain, E.; Haddad, J.G.; Koishi, A.C.; Sinigaglia, L.; Rachidi, W.; Desprès, P.; dos Santos, C.N.D.; Guiraud, P.; Jouvenet, N.; El Kalamouni, C. The Polyphenol-Rich Extract from Psiloxylon mauritianum, an Endemic Medicinal Plant from Reunion Island, Inhibits the Early Stages of Dengue and Zika Virus Infection. Int. J. Mol. Sci. 2019, 20, 1860. [Google Scholar] [CrossRef]
- Khan, E.; Hasan, R. Dengue infection in Asia; a regional concern. J. Postgrad. Med. Inst. 2012, 26, 1. [Google Scholar]
- Ooi, E.-E.; Gubler, D.J. Dengue in Southeast Asia: Epidemiological characteristics and strategic challenges in disease prevention. Cad. De Saude Publica 2009, 25, S115–S124. [Google Scholar] [CrossRef]
- Kadir, S.L.A.; Yaakob, H.; Zulkifli, R. Potential anti-dengue medicinal plants: A review. J. Nat. Med. 2013, 67, 677–689. [Google Scholar] [CrossRef]
- Klawikkan, N.; Nukoolkarn, V.; Jirakanjanakit, N.; Yoksan, S.; Wiwat, C. Effect of Thai medicinal plant extracts against Dengue virus in vitro. Mahidol Univ. J. Pharm. Sci. 2010, 38, 13–18. [Google Scholar]
- Vij, T.; Prashar, Y. A review on medicinal properties of Carica papaya Linn. Asian Pac. J. Trop. Dis. 2015, 5, 1–6. [Google Scholar] [CrossRef]
- Subenthiran, S.; Choon, T.C.; Cheong, K.C.; Thayan, R.; Teck, M.B.; Muniandy, P.K.; Afzan, A.; Abdullah, N.R.; Ismail, Z. Carica papaya Leaves Juice Significantly Accelerates the Rate of Increase in Platelet Count among Patients with Dengue Fever and Dengue Haemorrhagic Fever. Evid. Based Complement. Altern. Med. 2013, 2013, 616737. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, N.; Fazal, H.; Ayaz, M.; Abbasi, B.H.; Mohammad, I.; Fazal, L. Dengue fever treatment with Carica papaya leaves extracts. Asian Pac. J. Trop. Biomed. 2011, 1, 330–333. [Google Scholar] [CrossRef]
- Dubrulle, M.; Mousson, L.; Moutailler, S.; Vazeille, M.; Failloux, A.-B. Chikungunya Virus and Aedes Mosquitoes: Saliva Is Infectious as soon as Two Days after Oral Infection. PLoS ONE 2009, 4, e5895. [Google Scholar] [CrossRef] [PubMed]
- McFee, R.B. Selected mosquito-borne illnesses—Chikungunya. Dis.-A-Mon. 2018, 64, 222. [Google Scholar] [CrossRef]
- Wiwanitkit, S.; Wiwanitkit, V. Zika virus infection: Existence of hemoconcentration. Ann. Trop. Med. Public Health 2017, 10, 486. [Google Scholar] [CrossRef]
- Yang, M.; Sun, H.; Lai, H.; Hurtado, J.; Chen, Q. Plant-produced Zika virus envelope protein elicits neutralizing immune responses that correlate with protective immunity against Zika virus in mice. Plant Biotechnol. J. 2018, 16, 572–580. [Google Scholar] [CrossRef]
- Aboelsoud, N.H. Herbal medicine in ancient Egypt. J. Med. Plants Res. 2010, 4, 82–86. [Google Scholar]
- Bartak, M.; Lange, A.; Słonska, A.; Cymerys, J. Antiviral and healing potential of Sambucus nigra extracts. Bionatura 2020, 5, 1264–1270. [Google Scholar] [CrossRef]
- Alam, A.; Ferdosh, S.; Ghafoor, K.; Hakim, A.; Juraimi, A.S.; Khatib, A.; Sarker, Z.I. Clinacanthus nutans: A review of the medicinal uses, pharmacology and phytochemistry. Asian Pac. J. Trop. Med. 2016, 9, 402–409. [Google Scholar] [CrossRef]
- Thongchai, S.; Ekalaksananan, T.; Pientong, C.; Aromdee, C.; Seubsasana, S.; Sukpol, C.; Kongyingyoes, B. Anti-herpes simplex virus type 1 activity of crude ethyl acetate extract derived from leaves of Clinacanthus nutans Lindau. Khon Kaen Univ. J. 2008, 8, 65–73. [Google Scholar]
- Pongmuangmul, S.; Phumiamorn, S.; Sanguansermsri, P.; Wongkattiya, N.; Fraser, I.H.; Sanguansermsri, D. Anti-herpes simplex virus activities of monogalactosyl diglyceride and digalactosyl diglyceride from Clinacanthus nutans, a traditional Thai herbal medicine. Asian Pac. J. Trop. Biomed. 2016, 6, 192–197. [Google Scholar] [CrossRef]
- Yarnell, E. Herbs against human papillomavirus. Altern. Complementary Ther. 2015, 21, 71–76. [Google Scholar] [CrossRef]
- Fink, R.C.; Roschek, J.B.; Alberte, R.S. HIV Type-1 Entry Inhibitors with a New Mode of Action. Antivir. Chem. Chemother. 2009, 19, 243–255. [Google Scholar] [CrossRef] [PubMed]
- Barnes, J.; Anderson, L.A.; Gibbons, S.; Phillipson, J.D. Echinacea species (Echinacea angustifolia (DC.) Hell., Echinacea pallida (Nutt.) Nutt., Echinacea purpurea (L.) Moench): A review of their chemistry, pharmacology and clinical properties. J. Pharm. Pharmacol. 2005, 57, 929–954. [Google Scholar] [CrossRef] [PubMed]
- Fusco, D.; Liu, X.; Savage, C.; Taur, Y.; Xiao, W.; Kennelly, E.; Yuan, J.; Cassileth, B.; Salvatore, M.; Papanicolaou, G.A. Echinacea purpurea aerial extract alters course of influenza infection in mice. Vaccine 2010, 28, 3956–3962. [Google Scholar] [CrossRef]
- Cech, N.B.; Kandhi, V.; Davis, J.M.; Hamilton, A.; Eads, D.; Laster, S.M. Echinacea and its alkylamides: Effects on the influenza A-induced secretion of cytokines, chemokines, and PGE2 from RAW 264.7 macrophage-like cells. Int. Immunopharmacol. 2010, 10, 1268–1278. [Google Scholar] [CrossRef]
- Kim, M.; Kim, S.-Y.; Lee, H.W.; Shin, J.S.; Kim, P.; Jung, Y.-S.; Jeong, H.-S.; Hyun, J.-K.; Lee, C.-K. Inhibition of influenza virus internalization by (−)-epigallocatechin-3-gallate. Antivir. Res. 2013, 100, 460–472. [Google Scholar] [CrossRef]
- Ide, K.; Kawasaki, Y.; Kawakami, K.; Yamada, H. Anti-influenza Virus Effects of Catechins: A Molecular and Clinical Review. Curr. Med. Chem. 2016, 23, 4773–4783. [Google Scholar] [CrossRef]
- Li, R.; Tee, C.S.; Jiang, Y.L.; Jiang, X.Y.; Venkatesh, P.N.; Sarojam, R.; Ye, J. A terpenoid phytoalexin plays a role in basal defense of Nicotiana benthamiana against Potato virus X. Sci. Rep. 2015, 5, 9682. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antiviral Drugs | Mechanism of Actions | Viruses |
|---|---|---|
| Acyclovir, Valacyclovir | Valacyclovir and valganciclovir are hydrolyzed with the presence of esterases secreted from the intestine and liver into nucleoside analogs, acyclovir, and ganciclovir, respectively [16,17]. Acyclovir and ganciclovir go through phosphorylation, catalyzed by intracellular and viral kinase into acyclovir-triphosphate and ganciclovir, respectively. Both triphosphate molecules act as substrates for viral DNA polymerization, compete with deoxyguanosine triphosphate (dGTP) and inhibit the elongation of the viral DNA chain [15,16]. | HSV-1, HSV-2, and VZV |
| Ganciclovir, Valganciclovir | Cytomegalovirus (CMV) and HSV-1, HSV-2 | |
| Amantadine, Rimantadine | Interrupts the ion-channel activity of the M2 protein of influenza A by deforming the subunits of the channel [18]. The interruption inhibits the uncoating of viral RNA, thus preventing viral replication [19]. | Influenza A |
| Zidovudine (ZDV), Lamivudine (3TC) | Both antiretroviral drugs work synergically through phosphorylation into zidovudine and lamivudine triphosphate, respectively, in host cells. Both triphosphate anabolites inhibit the enzyme for HIV reverse transcription and polymerization of hepatitis B virus, thus preventing the viral DNA elongation and viral multiplication [20,21]. | HIV and hepatitis B virus |
| Remdesivir | This nucleoside analog prodrug undergoes a tri-phosphorylation process into remdesivir triphosphate. The nucleoside triphosphate embodies the new strand as the substrate for SARS-CoV-2 RNA-dependent RNA polymerase [22]. | SARS-CoV-2 |
| Nirmatrelvir | Also known as PF-07321332, a modified version of PF-00835231, hydroxymethyl ketone derivative. Nirmatrelvir inhibits the 3CL protease activity, an active drug target of SARV-CoV-2 that stimulates the cleave of the viral protein polypeptide bond [23]. | SARS-CoV-2 |
| Plant Name | Parts Used | Plant Extract | IC50/ED50/EC50/CC50/MIC100 | Assay | Cell Line Used | Mechanism of Action | Outcome |
|---|---|---|---|---|---|---|---|
| Echinacea pallida var. angustifolia [31] | Root | 70% ethanol | MIC₁00 = 62 μg/mL) | Viral plaque | Vero, H-1 and BEAS-2B | Inhibition of rhinovirus replication | Plant showed anti-rhinovirus activity |
| Ethyl acetate | MIC₁00 = 85 μg/mL | ||||||
| Hexane | MIC₁00 = 69 μg/mL | ||||||
| Echinacea purpurea [32] | Aerial parts and root | v/v 65% ethanol | IC50 = 3.2 μg/mL | MTT | Huh-7, Vero, and Vero E6 | Inhibition of HCoV-229E penetration into the cell and progeny shedding | HCoV-229E virucidal activity |
| Sambucus Formosana Nakai [33] | Stem | Ethanol | IC50 = 1.17 ± 0.75 μg/mL | Virus yield reduction | LLC-MK2 | Inhibition of binding of HCoV-NL63 with Sai cell-surface receptor | HCoV-NL63 virucidal activity |
| IC50 = 4.67 ± 1.21 μg/mL | Viral plaque | ||||||
| IC50 = 15.75 ± 6.65 μg/mL | Virus attachment | ||||||
| CC50 = 180.62 ± 63.04 μg/mL | MTT | ||||||
| Plantago asiatica [34] | Whole plant | Aqueous | CC50 = 938.43 μg/mL EC50 = 39.82 μg/mL SI value = 23.5 | Cell cytotoxicity, standard viral plaque, RSV-GFP virus replication inhibition | Human epithelial type 2: HEp2 with HeLa contaminant and A549 | Reduced RSV replication, RSV-induced cell death, RSV gene transcription, RSV protein synthesis, blocked syncytia formation | Antiviral therapy for respiratory syncytial virus (RSV) infection |
| Clerodendrum trichotomum [34] | Whole plant | Aqueous | EC50 = 27.95 μg/mL CC50 = 764.17 μg/mL. SI value = 27.3 | Cell cytotoxicity, standard viral plaque, RSV-GFP virus replication inhibition | Human epithelial type 2: HEp2with HeLa contaminant and A549 | Reduced RSV replication, RSV-induced cell death, RSV gene transcription, RSV protein synthesis, blocked syncytia formation | Antiviral therapy for respiratory syncytial virus (RSV) infection |
| Plant Name | Parts Used | Plant Extract | IC50/ED50/EC50/CC50 | Assay | Cell Line | Mechanism of Action | Outcome |
|---|---|---|---|---|---|---|---|
| Punica granatum | Peel [54] | 80% ethanol | IC50 = 6.45 μg/mL CC50 = 55.66 μg/mL SI value = 8.63 | Cytotoxicity, Cytopathic effect reduction | MDCK cells | Inhibition of influenza virus replication | Plant has high inhibitory effect on Influenza A virus replication in vitro |
| Fruit [55,56] | Polyphenol extract | Real-time PCR, viral plaque, TCID50, MTT | Changed viral surface glycoproteins and promoted damage to virion integrity | Plant exhibit virucidal effect on Influenza A virus | |||
| Geranium sanguineum [57,58] | AerialRoots | Polyphenol-rich methanol and ethanol extract | EC50 = 3.6 μg/mL IC50 = 64 μg/mL | Cytopathogenic effect reduction | MDCK cells | Inhibited the CPE of Influenza A/Rostock virus in CEF cells | Plant extract showed virucidal effect on Influenza A/Rostock virus |
| IC50 = 72 μg/mL | CEF cells | ||||||
| Echinacea purpurea [37] | Roots | Aqueous fraction | MIC100 = 2.2–2.5 μg/mL | Cytopathogenic effect reduction, viral plaque | Vero, H-1, and BEAS-2B cell line | Eliminated viral cytopathic effect | Plant root contains a potent water-soluble antiviral ingredient |
| Echinacea pallida var. angustifolia [37] | Roots | Ethanol 55% extract | MIC100 = 33.5 μg/mL | Cytopathogenic effect reduction, viral plaque | Vero, H-1, and BEAS-2B cell line | Eliminated viral cytopathic effect | Plant root contains a potent water-soluble antiviral ingredient |
| Ethyl acetate extract | MIC₁00 = 348 μg/mL | ||||||
| Cistus incanus [59,60] | Dinstinct variety of Cistus incanus | CYSTUS052 extract | Effective dose = 50 μg/mL | Aerosol formulation treatment, plaque reduction, hemagglutination | MDCK cells | Prevented adsorption of the Influenza A virus to host cells | Plant extract reduced 90% Influenza A viral plaque formation |
| Inbred female Balb/c and C57Bl/6 mice (age of 6–8 weeks) | Protected the animals against clinical disease symptoms caused by Influenza A virus infection | CYSTUS052-treated mice did not develop disease after being infected with the Influenza A | |||||
| Glycyrrhiza uralensis, Glycyrrhiza inflate and Glycyrrhiza glabra (Licorice) [61] | Root and rhizomes | Glycyrrhizin | IC50 = 0.0081 μg/mL | Cell viability, CPE reduction, Virus yield reduction | Natural killer (NK) cells, Human peripheral blood mononuclear cells (PBMC) | Reduced H5N1-induced cytokine expression, H5N1-induced caspase activation, and H5N1-induced apoptosis | Glycyrrhizin may develop into potential anti-influenza drug |
| Chaenomeles sinensis [62,63] | Fruit | Polyphenol-rich extract, CSD 3 | IC50 = 0.3 μg/mL | Plaque, hemagglutination, hemolysis | MDCK cells | Reduced cell-viral binding, hemagglutination, and hemolytic activities and infectivity of Influenza A/Udorn/72(H3N2) | Reduced primary transcription of Influenza A/Udorn/72(H3N2) virus-infected MDCK cells. |
| Sambucus nigra [64] | Fruit | Fruit juice at pH 4.4 ± 0.1 | IC50 = 6000 ± 800 μg/mL CC50 = 770 ± 60 μg/mL | Cytotoxicity, plaque reduction | MDCK cells | Suppressed viral entry, restrained viral transmission from cell to cell, and modulated the release of cytokines | Plant’s fruit exhibited multiple modes of therapeutic action against influenza infection |
| CC50 = 810 ± 30 μg/mL | A549 cells | ||||||
| Phyllanthus emblica | Branches and leaves [65] | 1,2,3,4,6-penta-O-galloyl-β-D-glucose (PGG) dissolved in DMSO | CC50 = 29.59 ± 4.32 μg/mL EC50 = 2.36 ± 0.29 μg/mL SI value = 12.54 | Water-soluble tetrazolium-1, Plaque-forming unit (PFU) | MDCK cells, A549 cells | Inhibited hemagglutination of chicken red blood cells induced by Influenza A virus, reduced virus budding and progeny virus release from infected cells | PGG as a promising antiviral agent against influenza A virus |
| Roots [66] | Fraction containing phyllaemblicin B | CC50 = 6.9 ± 0.9 μg/mL IC50 = 2.6 ± 0.7 μg/mL | Cytopathic end-point, Cytotoxicity | MDCK cells, Vero cells | Exerted inhibitory effect on Influenza A H3N2 strain | Both plant fractions showed potential anti-viral activities against Influenza A H3N2 strain | |
| Fraction containing glochicoccinoside D | CC50 =13.4 ± 2.1 μg/mL IC50 = 4.5 ± 0.6 μg/mL | ||||||
| Camelia sinensis [67] | Leaves | Cathecin extract of (−)-epigallocatechin gallate (EGCG) | EC50 = 22–28 μg/mL CC50 = 275.4 ± 22.8 μg/mL | Plaque inhibition, Virus growth inhibition, Hemagglutination inhibition, Neuraminidase inhibition, MTT | MDCK cells | Inhibited hemagglutination inhibition activity, suppressed viral RNA synthesis, inhibited neuraminidase activity, inhibited viral attachment, altered viral and cell membrane | Inhibitory potential on Influenza A virus was shown in order of ECGC > ECG > EGC |
| Cathecin extract of (−)-epicatechin gallate (ECG) | EC50 = 22–40 μg/mL CC50 = 525.9 ± 30.7 μg/mL | ||||||
| Cathecin extract of (−)-epigallocatechin (EGC)) | EC50 = 309–318 μg/mL CC50 = 1233.1 ± 44.9 μg/mL |
| Plant Name | Parts Used | Plant Extract | IC50/ED50/EC50/CC50 | Assay | Cell Line | Mechanism of Action | Outcome |
|---|---|---|---|---|---|---|---|
| Clinacanthus nutans [87] | Leaves | n-hexane | IC50 = 32.05 µg/mL, 72.62 μg/mL CC50 = 1600 μg/mL | Plaque reduction, MTT | Vero | Inhibited HSV-1 and HSV-2 viral plaque formation | n-hexane and dichloromethane extracts exhibited better antiviral activity against HSV-1 than methanol extract |
| Dichloro-methane | IC50 = 44.50 ± 2.66 µg/mL, 65.19 µg/mL CC50 = 869 μg/mL | ||||||
| Methanol | IC50 = 64.93 µg/mL, 65.13 μg/mL CC50 = 1600 μg/mL | ||||||
| Clinacanthus siamensis [87] | Leaves | n-hexane | IC50 = 60.00 µg/mL, 46.52 µg/mL CC50 = 1600 μg/mL | Plaque reduction, MTT | Vero | Inhibited HSV-1 and HSV-2 viral plaque formation | Methanolic extract possessed the greatest anti-HSV-1. n-hexane and dichloromethane extracts exhibited the best anti-HSV-2 in vitro |
| Dichloromethane | IC50 = 55.69 ± 4.41 µg/mL, 49.63 µg/mL CC50 = 194 μg/mL | ||||||
| Methanol | IC50 = 37.39 µg/mL, 72.64 µg/mL CC50 = 1600 μg/mL | ||||||
| Cistus incanus [59,70] | Dinstinct variety of Cistus incanus (CYSTUS052 extract) | Aqueous (boiled) | EC50 = 8.06 μg/mL CC50 = 250 μg/mL | Time-of-addition (TOA), Virus attachment, Virus capture, MTT | HEK293T, H9, and LC5 | Blocked primary virus attachment to cells by selective targeting of the viral envelope glycoproteins | Exhibited broad antiviral activity with low risk of virus resistance |
| Polyphenol- enriched fraction | CC50 = 1200 μg/mL | Plant fraction possessed antiviral activity on HIV-1LAI | |||||
| Plantago major [43] | Whole plant | Pure compound (caffeic acid) | EC50 = 15.3 µg/mL, 87.3 µg/mL SI value = 671, 118 CC50 = 10,293 µg/mL | XTT | BCC-1/KMC | Inhibited HSV virus replication | Caffeic acid possessed the best anti-HSV viral activity than chlorogenic acid |
| Pure compound (chlorogenic acid) | EC50 = 47.6 µg/mL, 86.5 µg/mL SI value = 83.9, 46.2 CC50 = 3995 µg/mL | ||||||
| Polygonum minus [88] | Leaves, stem | Methanol | LC50 leaves = 875 µg/mL LC50 stem = 95 µg/mL | Virus attachment | Vero | Inhibited HSV-1 attachment | 0.1 LC50 gave higher cell survival |
| Plant Name | Parts Used | Plant Extract | IC50/ED50/EC50/CC50 | Assay | Cell Line | Mechanism of Action | Outcome |
|---|---|---|---|---|---|---|---|
| Sarracenia purpurea [96] | Whole plant | Juice | CC50 = 70–75 µg/mL SI value = 5–7 | VACV plaque | RK-13 | Prevented replication of monkeypox virus and variola virus | Sarracenia purpurea acted as effective inhibitor of poxvirus replication |
| Clinacanthus nutans [97,98] | Leaves | 5% C. nutans cream | IC50 = 76 µg/mL | Double-blinded clinical trial | 125 patients infected with VCV | Lowered the chance of developing clinical disease | Exhibited a positive curing effect against VZV infection |
| Matricaria recutita [99] | Whole plant | Essential oil | Not identified SI value = 20 | Plaque reduction | RC-37 | Interacted with the viral envelope and glycoproteins | Reduced the infectivity of the HSV-2 virus |
| Aloe vera [100,101] | Leaves | Hot glycerin extract | CC50 = 3238 µg/mL IC50 = 428 µg/mL SI value = 7.56 | Cytotoxicity, plaque reduction | Vero | Inhibited HSV replication pre-attachment of virus on the cell | Showed significant inhibitory effect on HSV |
| IC50 = 536 µg/mL SI value = 6.04 | Inhibited HSV replication post attachment of virus replication | ||||||
| Cornus canadensis [102] | Leaves | Ethanol | EC50 = 9 μg/mL | Plaque reduction, cytotoxicity | Vero | Inhibited the lysis plaque, inhibited virus absorption | Exhibited as the potent virus absorption inhibitors |
| Lysimachia mauritania [103] | Whole plant | Ethanol | IC50 = 26.09 µg/mL | Plaque reduction, cell viability | MRC-5 | Inhibited the replication of varicella-zoster virus | Showed potent inhibitory effects on VZV gene expression and replication |
| Mentha haplocalyx [104] | Whole plant | Aqueous | IC50 = 70.3 μg/mL | Cytotoxicity, MTT, immunoblotting | Vero | Blocked viral infection proinflammatory response | Showed antiviral and anti-inflammatory activities |
| Camellia sinensis [105] | Leaves | Polyphenon E (poly E) | Concentration 1 and 5 µg/mL Concentration 10, 25, and 50 µg/mL | Immunofluorescence | TCL-1 | Inhibited growth of HPV-immortalized cervical epithelial (TCL-1) cells | All compounds showed inhibitory response on growth of HPV and poly E |
| Epigallocatechin gallate (EGCG) | |||||||
| Hibiscus sabdariffa [106] | Leaves (red and green leaves) | Ethanol | Concentration of 10 and 15 mg/mL | Cytotoxicity | Hep-2 | Inhibited measles virus replication | Showed antiviral activities on pre and post-inoculative treatment |
| Glycyrrhiza uralensis [107,108] | Root and stolon | Aqueous | IC50 = 0.056 μg/mL | XTT | Human foreskin | Suppressed EV71 replication | Showed antiviral activity against EV71 and CVA16 infection |
| Glycyrrhizic acid | 200 μg/mL | MTT, plaque forming | Vero | Blocked viral replication of EV71 and CVA16 | Inhibited EV71 and CVA16 replication | ||
| Phyllanthus emblica [66] | Roots | Fraction containing glochicoccinoside D | IC50 = 2.6 ± 0.7 μg/mL | Cytopathic end-point, Cytotoxicity | MDCK, Vero | Inhibited EV71 | Showed potential anti-viral activities against EV71 |
| Fraction containing phyllaemblicin C | IC50 = 2.6 ± 0.8 μg/mL |
| Plant Name | Part Used | Plant Extract | IC50/ED50/EC50/CC50 | Assay | Cell Line Used | Mechanism of Action | Outcome |
|---|---|---|---|---|---|---|---|
| Alternanthera philoxeroids [127] | Leaves | Coumarin based | TD50 = 535.91 | MTT | C6/36 | Inhibited C6/36 cell lines and dengue virus | Petroleum ether extract had the strongest inhibitory effect on dengue virus |
| Petroleum ether | ED50 = 47.43 | ||||||
| Cladosiphon okamuranus [128] | Whole plant | Fucoidan | Concentration of 10 μg/mL | Focus-forming | BHK-21 | Inhibited virus infection | Fucoidan reduced DENV-2 infectivity by 20% at 10 μg/mL |
| Clinacanthus nutans | Aerial part [129] | 80% ethanol | IC50 31.04 μg/mL | Anti-inflammatory, anti-dengue, immune-modulating activity | Naïve Huh-7 | Anti-inflammatory, anti-dengue virus and immune-modulating activity | Possessed moderate anti-dengue virus activity |
| Leaves [130,131] | Chloroform | CC50 of 25 μg/mL | MTT, immunofluorescence | C6/36, A549 | Inhibited dengue viral 2 in pre-entry replication step and suppressed PGE2 production | Showed virucidal activity against dengue virus 2 | |
| Sambucus nigra [132] | Leaves and flowers | Methanolic | 400 µg/mL | Viral plaque, indirect immunofluorescence | BHK-21 and Vero | Protected cell monolayers pre-treated cells before dengue virus-2 infection | Exhibited anti-DENV-2 activity on pre-incubated cells before dengue virus-2 infection |
| Carica papaya | Leaves [133,134] | Chloroform | CC50 = 1000 μg/mL EC50 = 1000 μg/mL SI value = 1 | Plaque forming, cytotoxicity, anti-DENV2 | LLC-MK2 | Inhibited DENV2 growth | Possessed promising anti-dengue properties |
| Aqueous | CC50 = 10437 μg/mL IC50 = 137.6 µg/mL SI value = 75.85 | MTT, foci forming unit reduction (FFURA) | Vero | Inhibited the virus replication, decreased number of dengue viral foci | |||
| Picrorhiza kurroa | Whole plant [135] | Aqueous | Min concentration to inhibit the plaque = 10 μg/mL | Viral plaque, helicase, protease | The monkey kidney cells, Vero | Inhibited viral attachment, inhibited helicase and protease activities | Showed virucidal activity on Chikungunya virus |
| Ocimum tenuiflorum | |||||||
| Terminalia chebula | |||||||
| Rhapis excelsa [136] | Leaves | Chloroform | EC50 = 29.9 ± 0.9 μg/mL CC50 = 161.5 ± 19.2 μg/mL | Cytopathic effect inhibition and cytotoxicity | African monkey kidney epithelial (Vero) | Showed cytopathic effect, inhibitory activity on Vero cells and reduction in the Chikungunya viral load | Showed virucidal activity on Chikungunya virus |
| Vernonia amygdalina [136] | Leaves | Ethyl acetate | EC50 = 32.4 ± 1.3 μg/mL CC50 = 165.5 ± 9.2 μg/mL | African monkey kidney epithelial (Vero) | |||
| Nicotiana benthamiana (Wild type) | Leaves | CHKVmab extract | EC50 = 390.8 μg/mL | Plaque reduction | Vero (ATCC, CCL-81) | Neutralization activity against CHIKV | Plant monoclonal antibodies have the potential to be used as effective treatment to prevent CHIKV infection |
| Nicotiana benthamiana (glycoengineered) [137] | EC50 = 130.5 μg/mL | ||||||
| Aphloia theiformis [138] | Aerial parts | Aqueous | CC50 = 3000 µg/mL IC50 = 100 µg/mL SI value = 30 | Plaque-forming, Immunofluorescence | Vero and human-derived Huh7.5 hepatoma | Prevented the viral entry into host cells | Promising sources of naturally derived antiviral compounds to prevent ZIKV |
| Psiloxylon mauritianum [139] | Fresh aerial parts | Aqueous | CC50 = 1044 ± 106.2 μg/mL | MTT, genotoxicity, viral inactivation, Time-of-drug-addiction | Vero | Inhibited early steps of the viral replication | Showed antiviral activity against historical and contemporary strains of ZIKV |
| CC50 = 657 ± 15.7 μg/mL | A549 | ||||||
| CC50 = 353 ± 84.4 μg/mL | Human primary keratinocytes | ||||||
| CC50 = 820 ± 26.5 μg/mL | Fibroblast (FMa) | ||||||
| IC50 = 19.5 ± 4.8 μg/mL SI value = 53.5 |
| Plant | Active Compound (s) | Mechanism of Action |
|---|---|---|
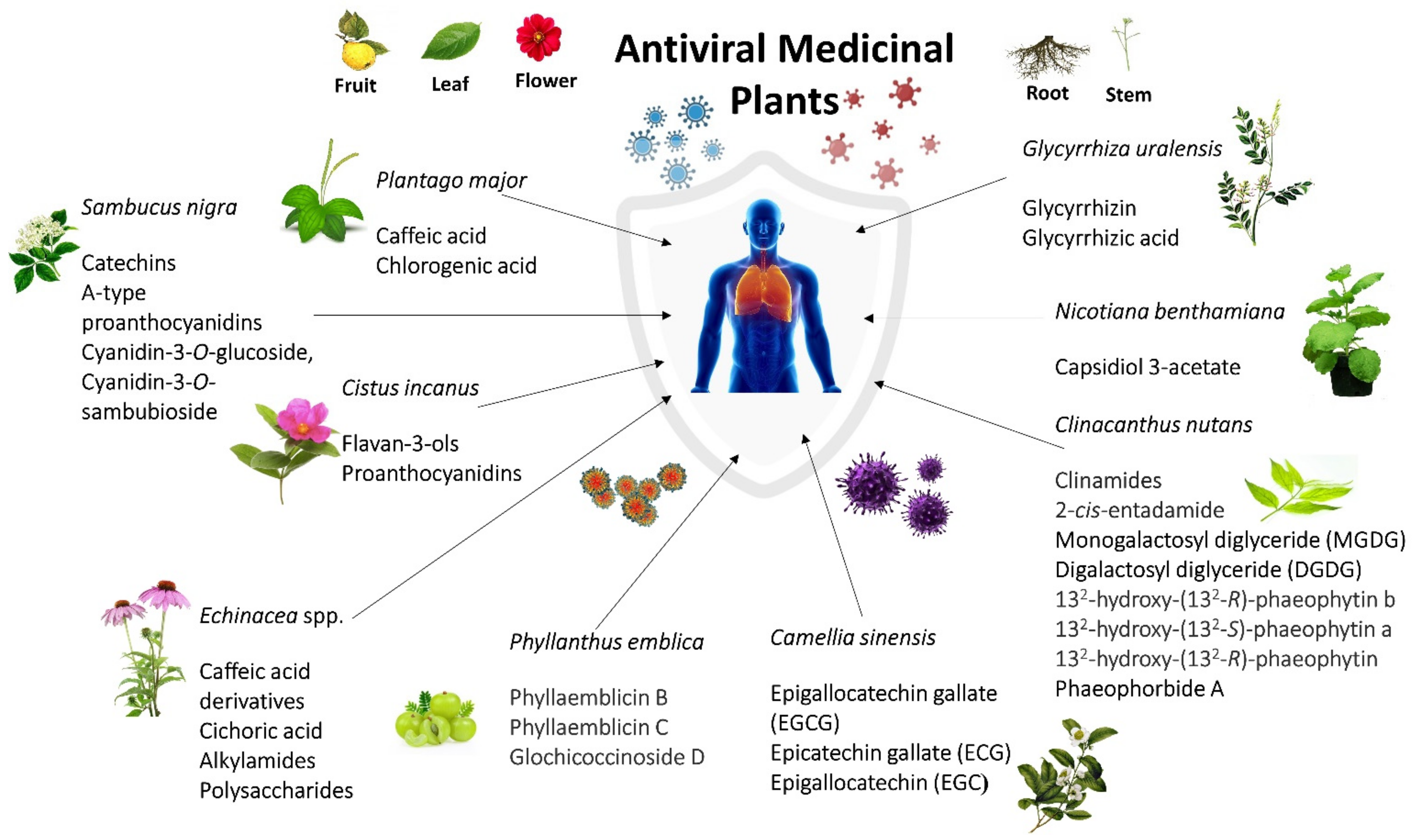
| Sambucus nigra | Catechins and A-type pro-anthocyanidins, cyanidin-3-O-glucoside, cyanidin-3-O-sambubioside | Bind to surface of influenza virus and prevent the influenza virus from entering and replicating in the host cell [152]. |
| Plant lectins bind to host cell membranes and prevent the influenza virus hemagglutinin’s attachment to host cells [39]. | ||
| Block the ability of HIV virions to infect host cells [157]. | ||
| Clinacanthus nutans | Clinamides and 2-cis-entadamide | Promote down-regulation of IFN-γ and exhibit immune-modulating activities [129]. |
| monogalactosyl diglyceride (MGDG) and digalactosyl diglyceride (DGDG). | Anti-HSV activities at post-infection stage [155]. | |
| 132-hydroxy-(132-R)-phaeophytin b, 132-hydroxy-(132-S)-phaeophytin a, and 132-hydroxy-(132-R)-phaeophytin | Affected the viral adsorption and penetration of HSV into host cells [130]. | |
| Phaeophorbide A | Inhibit the making of dengue virus RNA and protein in infected cells [130]. | |
| Echinacea spp. | Caffeic acid derivatives and cichoric acid | Enhanced innate immunity through activation of the neutrophils, macrophages, polymorphonuclear leukocytes (PMN), and natural killer (NK) cells [37,158]. |
| Caffeic acid derivatives, alkylamides, polysaccharides | Suppress cytokine and chemokine production from human blood monocytes stimulated by influenza viruses [159,160]. | |
| Plantago major | Caffeic acid, chlorogenic acid | Inhibit replication of HSV-1, HSV-2, ADV-3 and ADV-11 [43]. |
| Glycyrrhiza uralensis | Glycyrrhizin | Inhibition of influenza virus replication by inhibiting virus polymerase activity [75]. |
| Glycyrrhizic acid | Targeting early infection of coxsackievirus A16 on Vero cells to deactivate or inhibit coxsackievirus A16 infection [107]. | |
| Phyllanthus emblica | Phyllaemblicin B, phyllaemblicin C, and glochicoccinoside D | Displayed anti-viral activities and inhibitory activities against influenza A virus (H3N2), Enterovirus (EV71), coxsackievirus B3 and HSV-1[66]. |
| Camellia sinensis | Epigallocatechin gallate (EGCG) | Inhibits hemifusion events between virus particles and the cellular membrane by reducing the viral membrane integrity, thereby resulting in the loss of the cell penetration capacity of the influenza virus [161]. |
| Epicatechin gallate (ECG) and EGCG | Inhibits neuraminidase activity and blocks the function of viral neuraminidases of the influenza virus [162]. | |
| Exhibits hemagglutination inhibition activity [67]. | ||
| Suppresses viral RNA synthesis in MDCK cells [67]. | ||
| Epigallocatechin (EGC) | Inhibits neuraminidase activity and blocks the function of viral neuraminidases of the influenza virus [67]. | |
| Cistus incanus | flavan-3-ols and proanthocyanidins | Bind to the virus surface and inhibit the binding process of hemagglutinin to cellular receptors [60] |
| Prevent primary attachment of the HIV-1 and HIV-2 onto the cell surface [70]. | ||
| Blocking the viral attachment to cells and selective targeting the viral envelope glycoprotein [69]. | ||
| Nicotiana benthamiana | capsidiol 3-acetate | Inducing self-defense mechanism in Nicotiana benthamiana against Potato virus X infection [163]. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saifulazmi, N.F.; Rohani, E.R.; Harun, S.; Bunawan, H.; Hamezah, H.S.; Nor Muhammad, N.A.; Azizan, K.A.; Ahmed, Q.U.; Fakurazi, S.; Mediani, A.; et al. A Review with Updated Perspectives on the Antiviral Potentials of Traditional Medicinal Plants and Their Prospects in Antiviral Therapy. Life 2022, 12, 1287. https://doi.org/10.3390/life12081287
Saifulazmi NF, Rohani ER, Harun S, Bunawan H, Hamezah HS, Nor Muhammad NA, Azizan KA, Ahmed QU, Fakurazi S, Mediani A, et al. A Review with Updated Perspectives on the Antiviral Potentials of Traditional Medicinal Plants and Their Prospects in Antiviral Therapy. Life. 2022; 12(8):1287. https://doi.org/10.3390/life12081287
Chicago/Turabian StyleSaifulazmi, Nur Fadlin, Emelda Rosseleena Rohani, Sarahani Harun, Hamidun Bunawan, Hamizah Shahirah Hamezah, Nor Azlan Nor Muhammad, Kamalrul Azlan Azizan, Qamar Uddin Ahmed, Sharida Fakurazi, Ahmed Mediani, and et al. 2022. "A Review with Updated Perspectives on the Antiviral Potentials of Traditional Medicinal Plants and Their Prospects in Antiviral Therapy" Life 12, no. 8: 1287. https://doi.org/10.3390/life12081287
APA StyleSaifulazmi, N. F., Rohani, E. R., Harun, S., Bunawan, H., Hamezah, H. S., Nor Muhammad, N. A., Azizan, K. A., Ahmed, Q. U., Fakurazi, S., Mediani, A., & Sarian, M. N. (2022). A Review with Updated Perspectives on the Antiviral Potentials of Traditional Medicinal Plants and Their Prospects in Antiviral Therapy. Life, 12(8), 1287. https://doi.org/10.3390/life12081287