
Entomopathogenic Fungi Infecting Lepidopteran Larvae: A Case from Central Argentina

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hesketh, H.; Roy, H.E.; Eilenberg, J.; Pell, J.K.; Hails, R.S. Challenges in Modelling Complexity of Fungal Entomopathogens in Semi-Natural Populations of Insects. BioControl 2010, 55, 55–73. [Google Scholar] [CrossRef]
- Samson, R.A.; Evans, H.C.; Latge, J.-P. Atlas of Entomopathogenic Fungi; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2013; ISBN 978-3-662-05890-9. [Google Scholar]
- Kepler, R.M.; Luangsa-Ard, J.J.; Hywel-Jones, N.L.; Quandt, C.A.; Sung, G.-H.; Rehner, S.A.; Aime, M.C.; Henkel, T.W.; Sanjuan, T.; Zare, R.; et al. A Phylogenetically-Based Nomenclature for Cordycipitaceae (Hypocreales). IMA Fungus 2017, 8, 335–353. [Google Scholar] [CrossRef] [Green Version]
- Vinit, K. Phylogenetic Placement of Akanthomyces Muscarius, a New Endophyte Record from Nypa Fruticans in Thailand. CREAM 2018, 8, 404–417. [Google Scholar] [CrossRef]
- Santos, A.C.D.S.; Diniz, A.G.; Tiago, P.V.; de Oliveira, N.T. Entomopathogenic Fusarium Species: A Review of Their Potential for the Biological Control of Insects, Implications and Prospects. Fungal Biol. Rev. 2020, 34, 41–57. [Google Scholar] [CrossRef]
- Hajek, A.; Goettel, M. Guidelines for Evaluating Effects of Entomopathogens on Non-Target Organisms. In Field Manual of Techniques in Invertebrate Pathology; Lacey, L.A., Kaya, H.K., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 816–833. ISBN 978-1-4020-5931-5. [Google Scholar]
- Sharma, L.; Marques, G. Fusarium, an Entomopathogen—A Myth or Reality? Pathogens 2018, 7, 93. [Google Scholar] [CrossRef] [Green Version]
- Murphy, S.M.; Loewy, K.J. Trade-Offs in Host Choice of an Herbivorous Insect Based on Parasitism and Larval Performance. Oecologia 2015, 179, 741–751. [Google Scholar] [CrossRef] [PubMed]
- Gielen, R.; Põldmaa, K.; Tammaru, T. In Search of Ecological Determinants of Fungal Infections: A Semi-Field Experiment with Folivorous Moths. Ecol. Evol. 2022, in press. [CrossRef] [PubMed]
- Sánchez-Bayo, F.; Wyckhuys, K.A.G. Worldwide Decline of the Entomofauna: A Review of Its Drivers. Biol. Conserv. 2019, 232, 8–27. [Google Scholar] [CrossRef]
- Elliot; Sabelis; Janssen; Geest, V.D.; Beerling; Fransen. Can Plants Use Entomopathogens as Bodyguards? Ecol. Lett. 2000, 3, 228–235. [Google Scholar] [CrossRef] [Green Version]
- Vega, F.E.; Kaya, H.K. Insect Pathology, 2nd ed.; Academic Press: Cambridge, MA, USA, 2012; ISBN 978-0-12-384984-7. [Google Scholar]
- Amatuzzi, R.F.; Cardoso, N.; Poltronieri, A.S.; Poitevin, C.G.; Dalzoto, P.; Zawadeneak, M.A.; Pimentel, I.C. Potential of Endophytic Fungi as Biocontrol Agents of Duponchelia Fovealis (Zeller) (Lepidoptera:Crambidae). Braz. J. Biol. 2017, 78, 429–435. [Google Scholar] [CrossRef] [Green Version]
- McKinnon, A.C.; Saari, S.; Moran-Diez, M.E.; Meyling, N.V.; Raad, M.; Glare, T.R. Beauveria Bassiana as an Endophyte: A Critical Review on Associated Methodology and Biocontrol Potential. BioControl 2017, 62, 1–17. [Google Scholar] [CrossRef]
- Nicoletti, R.; Becchimanzi, A. Endophytism of Lecanicillium and Akanthomyces. Agriculture 2020, 10, 205. [Google Scholar] [CrossRef]
- Meyling, N.V.; Hajek, A.E. Principles from Community and Metapopulation Ecology: Application to Fungal Entomopathogens. BioControl 2010, 55, 39–54. [Google Scholar] [CrossRef]
- Poitevin, C.G.; Porsani, M.V.; Poltronieri, A.S.; Zawadneak, M.A.C.; Pimentel, I.C. Fungi Isolated from Insects in Strawberry Crops Act as Potential Biological Control Agents of Duponchelia Fovealis (Lepidoptera: Crambidae). Appl. Entomol. Zool. 2018, 53, 323–331. [Google Scholar] [CrossRef]
- Braga, M.P.; Janz, N. Host Repertoires and Changing Insect–Plant Interactions. Ecol. Entomol. 2021, 46, 1241–1253. [Google Scholar] [CrossRef]
- Singer, M.C.; Wee, B. Spatial Pattern in Checkerspot Butterfly—Host Plant Association at Local, Metapopulation and Regional Scales. Ann. Zool. Fenn. 2005, 42, 347–361. [Google Scholar]
- Põldmaa, K.; Bills, G.; Lewis, D.P.; Tamm, H. Taxonomy of the Sphaerostilbella Broomeana-Group (Hypocreales, Ascomycota). Mycol. Prog. 2019, 18, 77–89. [Google Scholar] [CrossRef]
- Abarenkov, K.; Tedersoo, L.; Nilsson, R.H.; Vellak, K.; Saar, I.; Veldre, V.; Parmasto, E.; Prous, M.; Aan, A.; Ots, M.; et al. PlutoF—A Web Based Workbench for Ecological and Taxonomic Research, with an Online Implementation for Fungal ITS Sequences. Evol. Bioinform. 2010, 6, EBO.S6271. [Google Scholar] [CrossRef]
- Kõljalg, U.; Nilsson, R.H.; Abarenkov, K.; Tedersoo, L.; Taylor, A.F.S.; Bahram, M.; Bates, S.T.; Bruns, T.D.; Bengtsson-Palme, J.; Callaghan, T.M.; et al. Towards a Unified Paradigm for Sequence-Based Identification of Fungi. Mol. Ecol. 2013, 22, 5271–5277. [Google Scholar] [CrossRef] [Green Version]
- Kõljalg, U.; Nilsson, H.R.; Schigel, D.; Tedersoo, L.; Larsson, K.-H.; May, T.W.; Taylor, A.F.S.; Jeppesen, T.S.; Frøslev, T.G.; Lindahl, B.D.; et al. The Taxon Hypothesis Paradigm—On the Unambiguous Detection and Communication of Taxa. Microorganisms 2020, 8, 1910. [Google Scholar] [CrossRef]
- Fox, J.; Weisberg, S. An R Companion to Applied Regression; SAGE Publications: Thousand Oaks, CA, USA, 2018; ISBN 978-1-5443-3648-0. [Google Scholar]
- Gielen, R.; Meister, H.; Tammaru, T.; Põldmaa, K. Fungi Recorded on Folivorous Lepidoptera: High Diversity Despite Moderate Prevalence. J. Fungi 2021, 7, 25. [Google Scholar] [CrossRef] [PubMed]
- Batta, Y.A. Efficacy of Endophytic and Applied Metarhizium Anisopliae (Metch.) Sorokin (Ascomycota: Hypocreales) against Larvae of Plutella Xylostella L. (Yponomeutidae: Lepidoptera) Infesting Brassica Napus Plants. Crop Prot. 2013, 44, 128–134. [Google Scholar] [CrossRef]
- St. Leger, R.J.; Wang, J.B. Metarhizium: Jack of All Trades, Master of Many. Open Biol. 2020, 10, 200307. [Google Scholar] [CrossRef] [PubMed]
- Tarasco, E.; Bievre, C.D.; Papierok, B.; Poliseno, M.; Triggiani, O. Occurrence of Entomopathogenic Fungi in Soils in Southern Italy. Entomologica 1997, 31, 157–166. [Google Scholar] [CrossRef]
- Vänninen, I. Distribution and Occurrence of Four Entomopathogenic Fungi in Finland: Effect of Geographical Location, Habitat Type and Soil Type. Mycol. Res. 1996, 100, 93–101. [Google Scholar] [CrossRef]
- Sun, B.-D.; Liu, X.-Z. Occurrence and Diversity of Insect-Associated Fungi in Natural Soils in China. Appl. Soil Ecol. 2008, 39, 100–108. [Google Scholar] [CrossRef]
- Chen, W.; Xie, W.; Cai, W.; Thaochan, N.; Hu, Q. Entomopathogenic Fungi Biodiversity in the Soil of Three Provinces Located in Southwest China and First Approach to Evaluate Their Biocontrol Potential. J. Fungi 2021, 7, 984. [Google Scholar] [CrossRef]
- Ali-Shtayeh, M.S.; Mara’i, A.-B.B.M.; Jamous, R.M. Distribution, Occurrence and Characterization of Entomopathogenic Fungi in Agricultural Soil in the Palestinian Area. Mycopathologia 2002, 156, 235–244. [Google Scholar] [CrossRef]
- O’Donnell, K.; Humber, R.A.; Geiser, D.M.; Kang, S.; Park, B.; Robert, V.A.R.G.; Crous, P.W.; Johnston, P.R.; Aoki, T.; Rooney, A.P.; et al. Phylogenetic Diversity of Insecticolous Fusaria Inferred from Multilocus DNA Sequence Data and Their Molecular Identification via FUSARIUM-ID and Fusarium MLST. Mycologia 2012, 104, 427–445. [Google Scholar] [CrossRef] [Green Version]
- Tedersoo, L.; Bahram, M.; Põlme, S.; Kõljalg, U.; Yorou, N.S.; Wijesundera, R.; Ruiz, L.V.; Vasco-Palacios, A.M.; Thu, P.Q.; Suija, A.; et al. Global Diversity and Geography of Soil Fungi. Science 2014, 346, 1256688. [Google Scholar] [CrossRef] [Green Version]
- Põldmaa, K. Records of Hypomyces, Including Two New Species, from Chanchamayo, Peru. Mycotaxon 2007, 102, 183–197. [Google Scholar]
- Põldmaa, K. The Genus Hypomyces and Allied Fungicolous Fungi in Estonia. I. Species Growing on Aphyllophoralean Basidiomycetes. Folia Cryptogam. Est. 1999, 34, 15–31. [Google Scholar]
- U’Ren, J.M.; Lutzoni, F.; Miadlikowska, J.; Zimmerman, N.B.; Carbone, I.; May, G.; Arnold, A.E. Host Availability Drives Distributions of Fungal Endophytes in the Imperilled Boreal Realm. Nat. Ecol. Evol. 2019, 3, 1430–1437. [Google Scholar] [CrossRef] [PubMed]
- Al-Hatmi, A.M.; Hagen, F.; Menken, S.B.; Meis, J.F.; de Hoog, G.S. Global Molecular Epidemiology and Genetic Diversity of Fusarium, a Significant Emerging Group of Human Opportunists from 1958 to 2015. Emerg. Microbes Infect. 2016, 5, e124. [Google Scholar] [CrossRef] [Green Version]
- Seye, F.; Bawin, T.; Boukraa, S.; Zimmer, J.-Y.; Ndiaye, M.; Delvigne, F.; Francis, F. Effect of Entomopathogenic Aspergillus Strains against the Pea Aphid, Acyrthosiphon Pisum (Hemiptera: Aphididae). Appl. Entomol. Zool. 2014, 49, 453–458. [Google Scholar] [CrossRef]
- Kamala Jayanthi, P.D.; Arthikirubha, A.; Vivek, K.; Ravindra, M.A.; Selvakumar, G.; Abraham, V. Aspergillus Flavus Impairs Antioxidative Enzymes of Sternochetus Mangiferae during Mycosis. J. Invertebr. Pathol. 2015, 124, 73–77. [Google Scholar] [CrossRef]
- Karthi, S.; Vaideki, K.; Shivakumar, M.S.; Ponsankar, A.; Thanigaivel, A.; Chellappandian, M.; Vasantha-Srinivasan, P.; Muthu-Pandian, C.K.; Hunter, W.B.; Senthil-Nathan, S. Effect of Aspergillus Flavus on the Mortality and Activity of Antioxidant Enzymes of Spodoptera Litura Fab. (Lepidoptera: Noctuidae) Larvae. Pestic. Biochem. Physiol. 2018, 149, 54–60. [Google Scholar] [CrossRef]


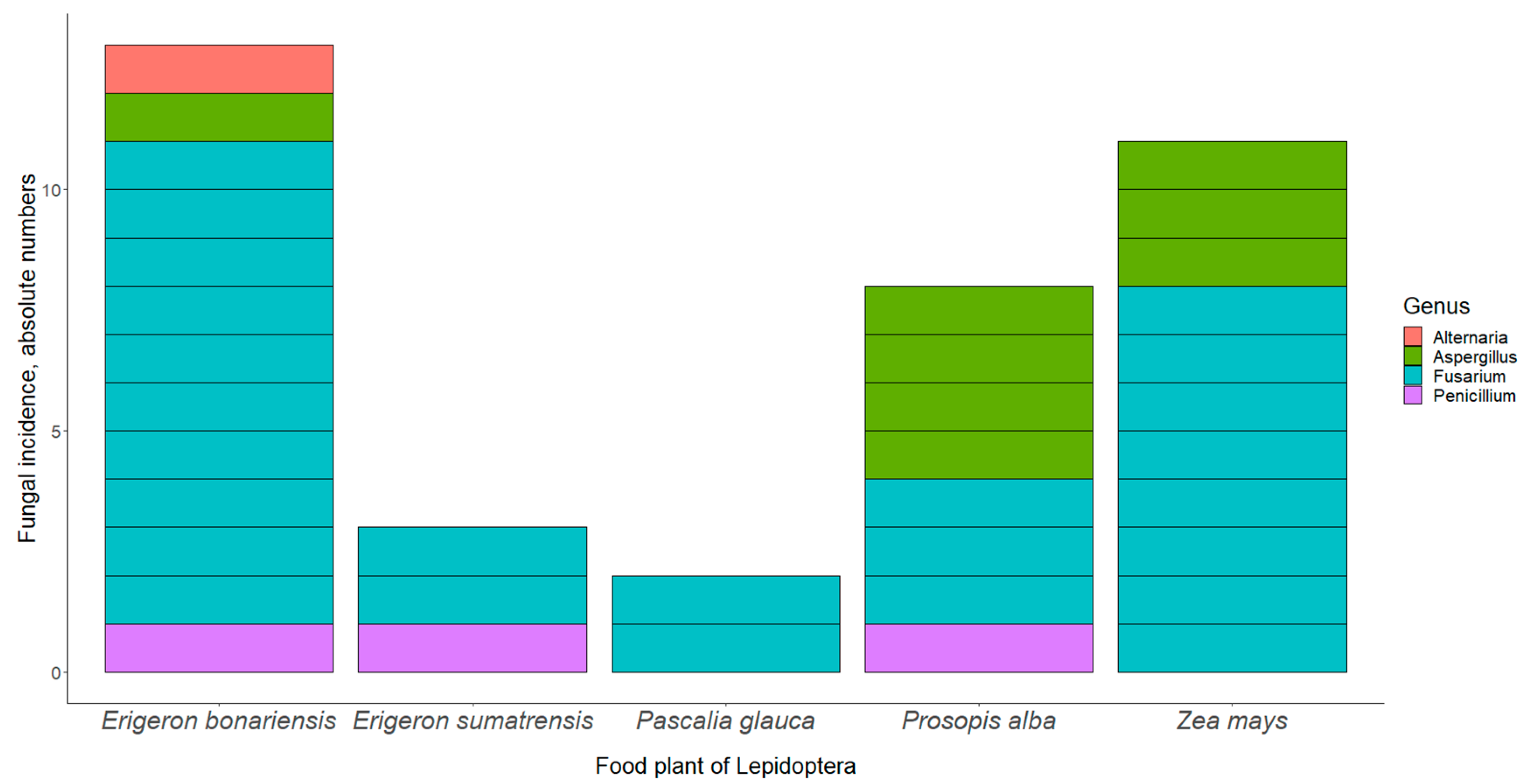{kind=link}
| Lepidopteran Species | Mother/Brood No | Collected From | Date | No of Larvae | Larvae Died of Fungi |
|---|---|---|---|---|---|
| Anicla infecta | AI4 | 31°18′22″ S 64°20′43″ W | 4 January 2020 | 73 | 0 |
| AI6 | 31°08′21″ S 64°21′48″ W | 9 January 2020 | 86 | 0 | |
| AI7 | 31°08′21″ S 64°21′48″ W | 9 January 2020 | 42 | 3 | |
| AI8 | 31°08′21″ S 64°21′48″ W | 9 January 2020 | 6 | 0 | |
| AI9 | 31°08′21″ S 64°21′48″ W | 9 January 2020 | 12 | 1 | |
| AI16 | 31°18′22″ S 64°20′43″ W | 28 January 2020 | 20 | 3 | |
| Dargida albilinea | FA17 | 31°51′26″ S 63°44′16″ W | 17 December 2019 | 116 | 4 |
| FA20 | 31°51′26″ S 63°44′16″ W | 17 December 2019 | 93 | 2 | |
| Hypercompe indecisa | HI1 | 31°18′22″ S 64°20′43″ W | 23 December 2019 | 221 | 12 |
| HI3 | 31°18′22″ S 64°20′43″ W | 4 January 2020 | 309 | 12 |
| Order/Family | Species | SH DOI | Lepidopteran Species | Food Plant |
|---|---|---|---|---|
| Eurotiales/Aspergillaceae | Aspergillus flavus Link | SH1532328.08FU | A. infecta 1 D. albilinea 2 H. indecisa 5 | E. bonariensis 1 P. alba 4 Z. mays 3 |
| Penicillium sp. 1 Link | A. infecta 1 H. indecisa 2 | P. alba 1 E. bonariensis 1 E. sumatrensis 1 | ||
| Hypocreales/Nectriaceae | Fusarium chlamydosporum sc* | SH1610186.08FU | D. albilinea 1 | Z. mays 1 |
| Fusarium fujikuroi sc* | SH1610157.08FU | A. infecta 3 D. albilinea 1 H. indecisa 8 | E. bonariensis 10 Z. mays 2 | |
| Fusarium incarnatum-equiseti sc* | SH1458596.08FU | A. infecta 1 D. albilinea 1 H. indecisa 7 | E. sumatrensis 2 P. alba 3 Z. mays 4 | |
| Fusarium oxysporum sc* 2 | SH1656686.08FU | D. albilinea 1 | P. glauca 1 | |
| Fusarium solani sc* | SH1623679.08FU | A. infecta 1 H. indecisa 1 | P. glauca 1 Z. mays 1 | |
| Pleosporales/Pleosporaceae | Alternaria sp. 3 Nees. | SH1526648.08FU | H. indecisa 1 | E. bonariensis 1 |
| df | χ2 | p | |
|---|---|---|---|
| Total data | |||
| Lepidoptera sp. | 2 | 1.8 | 0.41 |
| Food plant (Lep. sp.) | 12 | 29.9 | 0.003 |
| Genus Fusarium | |||
| Lepidoptera sp. | 2 | 0.05 | 0.97 |
| Food plant (Lep. sp.) | 9 | 8.9 | 0.45 |
| Fusarium fujikuroi | |||
| Lepidoptera sp. | 2 | 0.85 | 0.65 |
| Food plant (Lep. sp.) | 9 | 26.1 | 0.002 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gielen, R.; Robledo, G.; Zapata, A.I.; Tammaru, T.; Põldmaa, K. Entomopathogenic Fungi Infecting Lepidopteran Larvae: A Case from Central Argentina. Life 2022, 12, 974. https://doi.org/10.3390/life12070974
Gielen R, Robledo G, Zapata AI, Tammaru T, Põldmaa K. Entomopathogenic Fungi Infecting Lepidopteran Larvae: A Case from Central Argentina. Life. 2022; 12(7):974. https://doi.org/10.3390/life12070974
Chicago/Turabian StyleGielen, Robin, Gerardo Robledo, Adriana Inés Zapata, Toomas Tammaru, and Kadri Põldmaa. 2022. "Entomopathogenic Fungi Infecting Lepidopteran Larvae: A Case from Central Argentina" Life 12, no. 7: 974. https://doi.org/10.3390/life12070974
APA StyleGielen, R., Robledo, G., Zapata, A. I., Tammaru, T., & Põldmaa, K. (2022). Entomopathogenic Fungi Infecting Lepidopteran Larvae: A Case from Central Argentina. Life, 12(7), 974. https://doi.org/10.3390/life12070974