
Phloretin in Benign Prostate Hyperplasia and Prostate Cancer: A Contemporary Systematic Review

 ,
,  ,
,
Abstract
1. Introduction
2. Phloretin, BPH, and PCa
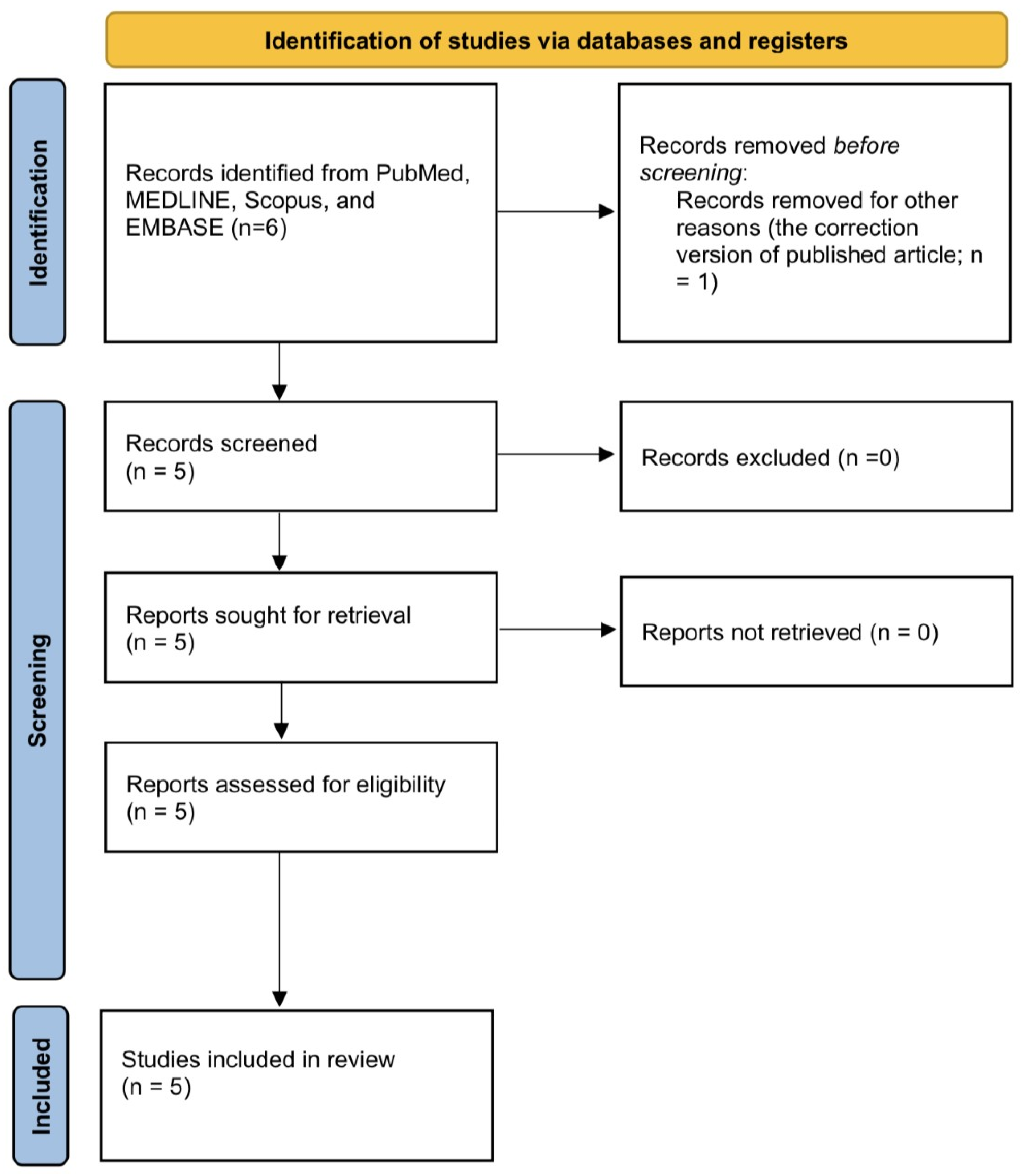
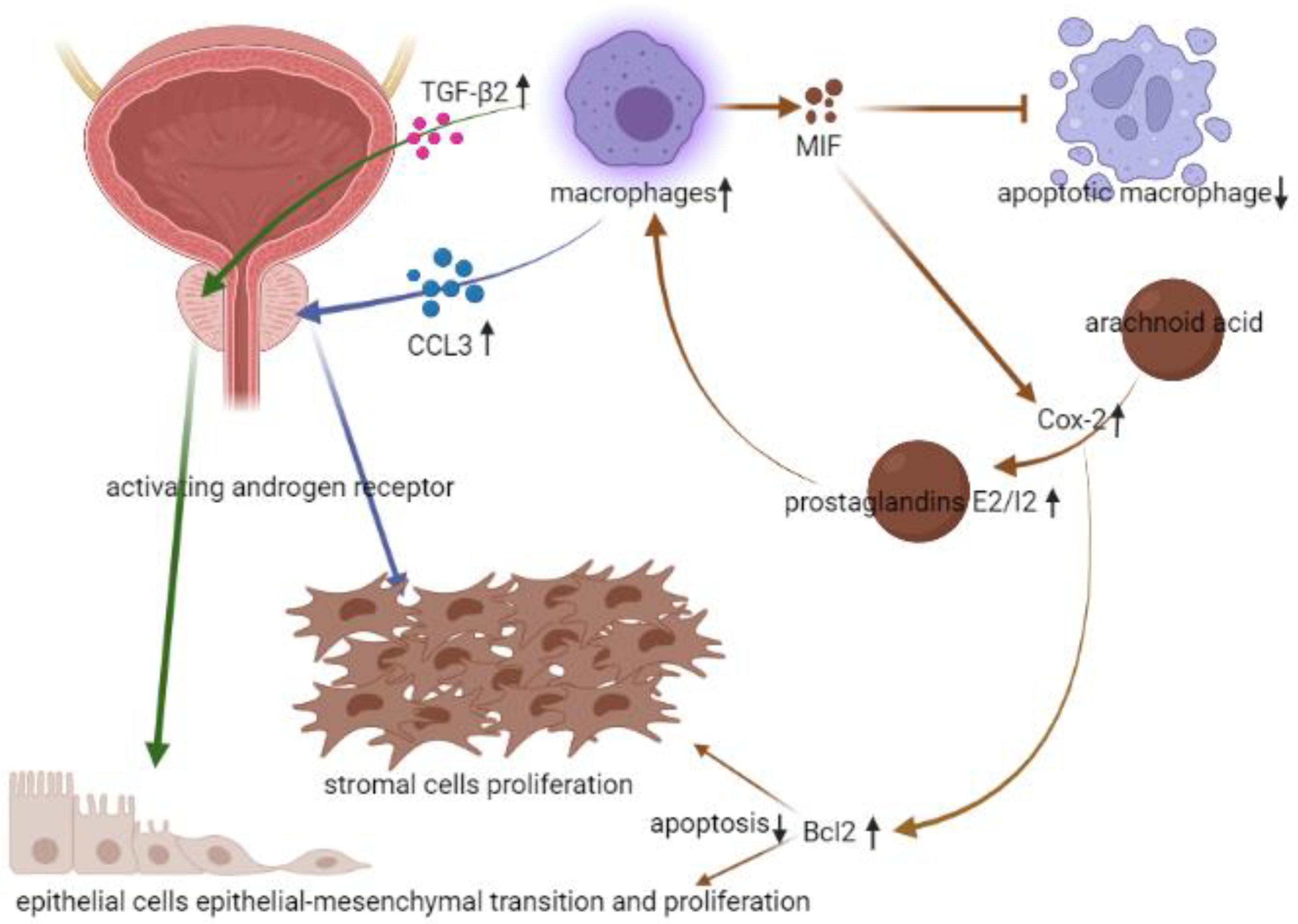
2.1. Immune Microenvironment in BPH
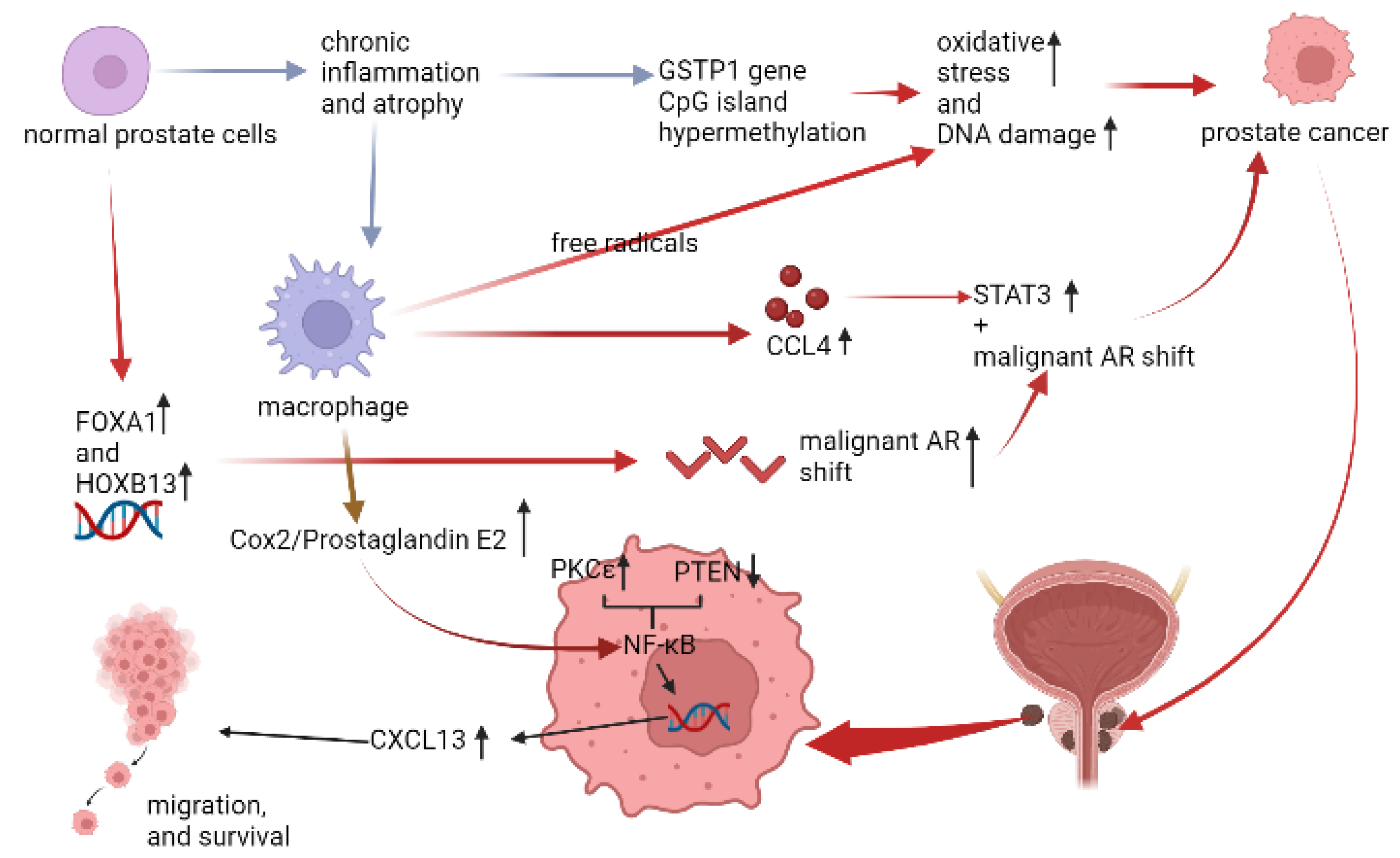
2.2. Immune Microenvironment in PCa
2.3. Phloretin and the Current Medical Options for BPH
2.4. The Current Medical Options for PCa
3. Perspective of Future Studies of Phloretin in Treating BPH and PCa
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Commisso, M.; Bianconi, M.; Poletti, S.; Negri, S.; Munari, F.; Ceoldo, S.; Guzzo, F. Metabolomic Profiling and Antioxidant Activity of Fruits Representing Diverse Apple and Pear Cultivars. Biology 2021, 10, 380. [Google Scholar] [CrossRef]
- Choi, B.Y. Biochemical Basis of Anti-Cancer-Effects of Phloretin—A Natural Dihydrochalcone. Molecules 2019, 24, 278. [Google Scholar] [CrossRef] [PubMed]
- Mendes, R.A.; e Silva, B.L.S.; Takeara, R.; Freitas, R.G.; Brown, A.; de Souza, G.L.C. Probing the antioxidant potential of phloretin and phlorizin through a computational investigation. J. Mol. Model. 2018, 24, 101. [Google Scholar] [CrossRef] [PubMed]
- Schaefer, S.; Baum, M.; Eisenbrand, G.; Dietrich, H.; Will, F.; Janzowski, C. Polyphenolic apple juice extracts and their major constituents reduce oxidative damage in human colon cell lines. Mol. Nutr. Food Res. 2006, 50, 24–33. [Google Scholar] [CrossRef]
- Park, S.Y.; Kim, E.J.; Shin, H.K.; Kwon, D.Y.; Kim, M.S.; Surh, Y.J.; Park, J.H. Induction of apoptosis in HT-29 colon cancer cells by phloretin. J. Med. Food 2007, 10, 581–586. [Google Scholar] [CrossRef]
- Ma, L.; Wang, R.; Nan, Y.; Li, W.; Wang, Q.; Jin, F. Phloretin exhibits an anticancer effect and enhances the anticancer ability of cisplatin on non-small cell lung cancer cell lines by regulating expression of apoptotic pathways and matrix metalloproteinases. Int. J. Oncol. 2016, 48, 843–853. [Google Scholar] [CrossRef] [PubMed]
- Diakos, C.I.; Charles, K.A.; McMillan, D.C.; Clarke, S.J. Cancer-related inflammation and treatment effectiveness. Lancet Oncol. 2014, 15, e493–e503. [Google Scholar] [CrossRef]
- DiDonato, J.A.; Mercurio, F.; Karin, M. NF-κB and the link between inflammation and cancer. Immunol. Rev. 2012, 246, 379–400. [Google Scholar] [CrossRef]
- Huang, W.C.; Lai, C.L.; Liang, Y.T.; Hung, H.C.; Liu, H.C.; Liou, C.J. Phloretin attenuates LPS-induced acute lung injury in mice via modulation of the NF-κB and MAPK pathways. Int. Immunopharmacol. 2016, 40, 98–105. [Google Scholar] [CrossRef]
- Jung, M.; Triebel, S.; Anke, T.; Richling, E.; Erkel, G. Influence of apple polyphenols on inflammatory gene expression. Mol. Nutr. Food Res. 2009, 53, 1263–1280. [Google Scholar] [CrossRef]
- Hsu, C.Y.; Lin, Y.S.; Weng, W.C.; Panny, L.; Chen, H.L.; Tung, M.C.; Ou, Y.C.; Lin, C.C.; Yang, C.H. Phloretin Ameliorates Testosterone-Induced Benign Prostatic Hyperplasia in Rats by Regulating the Inflammatory Response, Oxidative Stress and Apoptosis. Life 2021, 11, 743. [Google Scholar] [CrossRef] [PubMed]
- Kim, U.; Kim, C.Y.; Lee, J.M.; Oh, H.; Ryu, B.; Kim, J.; Park, J.H. Phloretin Inhibits the Human Prostate Cancer Cells Through the Generation of Reactive Oxygen Species. Pathol. Oncol. Res. 2020, 26, 977–984. [Google Scholar] [CrossRef] [PubMed]
- Kang, D.; Zuo, W.; Wu, Q.; Zhu, Q.; Liu, P. Inhibition of Specificity Protein 1 Is Involved in Phloretin-Induced Suppression of Prostate Cancer. Biomed. Res. Int. 2020, 2020, 1358674. [Google Scholar] [CrossRef] [PubMed]
- Szliszka, E.; Czuba, Z.P.; Mazur, B.; Paradysz, A.; Krol, W. Chalcones and Dihydrochalcones Augment TRAIL-Mediated Apoptosis in Prostate Cancer Cells. Molecules 2010, 15, 5336–5353. [Google Scholar] [CrossRef]
- Gonzalez-Menendez, P.; Hevia, D.; Rodriguez-Garcia, A.; Mayo, J.C.; Sainz, R.M. Regulation of GLUT transporters by flavonoids in androgen-sensitive and -insensitive prostate cancer cells. Endocrinology 2014, 155, 3238–3250. [Google Scholar] [CrossRef]
- Hayward, S.W.; Baskin, L.S.; Haughney, P.C.; Foster, B.A.; Cunha, A.R.; Dahiya, R.; Prins, G.S.; Cunha, G.R. Stromal development in the ventral prostate, anterior prostate and seminal vesicle of the rat. Acta Anat. 1996, 155, 94–103. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.L.; Yuan, Y.; Geng, H.; Xia, S.J. Influence of immune inflammation on androgen receptor expression in benign prostatic hyperplasia tissue. Asian J. Androl. 2012, 14, 316–319. [Google Scholar] [CrossRef]
- Debes, J.D.; Tindall, D.J. The role of androgens and the androgen receptor in prostate cancer. Cancer Lett. 2002, 187, 1–7. [Google Scholar] [CrossRef]
- Mainwaring, W.I.; Mangan, F.R.; Wilce, P.A.; Milroy, E.G. Androgens. I. A review of current research on the binding and mechanism of action of androgenic steroids, notably 5 alpha-dihydrotestosterone. Adv. Exp. Med. Biol. 1973, 36, 197–231. [Google Scholar]
- Wang, G.C.; Huang, T.R.; Wang, K.Y.; Wu, Z.L.; Xie, J.B.; Zhang, H.L.; Yin, L.; Tang, W.L.; Peng, B. Inflammation induced by lipopolysaccharide advanced androgen receptor expression and epithelial-mesenchymal transition progress in prostatitis and prostate cancer. Transl. Androl. Urol. 2021, 10, 4275–4287. [Google Scholar] [CrossRef]
- Alonso-Magdalena, P.; Brössner, C.; Reiner, A.; Cheng, G.; Sugiyama, N.; Warner, M.; Gustafsson, J.A. A role for epithelial-mesenchymal transition in the etiology of benign prostatic hyperplasia. Proc. Natl. Acad. Sci. USA 2009, 106, 2859–2863. [Google Scholar] [CrossRef] [PubMed]
- Kong, D.; Sethi, S.; Li, Y.; Chen, W.; Sakr, W.A.; Heath, E.; Sarkar, F.H. Androgen receptor splice variants contribute to prostate cancer aggressiveness through induction of EMT and expression of stem cell marker genes. Prostate 2015, 75, 161–174. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Lin, W.J.; Izumi, K.; Jiang, Q.; Lai, K.P.; Xu, D.; Fang, L.Y.; Lu, T.; Li, L.; Xia, S.; et al. Increased infiltrated macrophages in benign prostatic hyperplasia (BPH): Role of stromal androgen receptor in macrophage-induced prostate stromal cell proliferation. J. Biol. Chem. 2012, 287, 18376–18385. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Wang, X.; Jiang, C.; Ruan, Y.; Xia, S.; Wang, X. The androgen receptor plays different roles in macrophage-induced proliferation in prostate stromal cells between transitional and peripheral zones of benign prostatic hypertrophy. EXCLI J. 2017, 16, 939–948. [Google Scholar] [PubMed]
- Lu, T.; Lin, W.J.; Izumi, K.; Wang, X.; Xu, D.; Fang, L.Y.; Li, L.; Jiang, Q.; Jin, J.; Chang, C. Targeting androgen receptor to suppress macrophage-induced EMT and benign prostatic hyperplasia (BPH) development. Mol. Endocrinol. 2012, 26, 1707–1715. [Google Scholar] [CrossRef]
- Mitchell, R.A.; Liao, H.; Chesney, J.; Fingerle-Rowson, G.; Baugh, J.; David, J.; Bucala, R. Macrophage migration inhibitory factor (MIF) sustains macrophage proinflammatory function by inhibiting p53: Regulatory role in the innate immune response. Proc. Natl. Acad. Sci. USA 2002, 99, 345–350. [Google Scholar] [CrossRef]
- Wang, W.; Bergh, A.; Damber, J.E. Chronic inflammation in benign prostate hyperplasia is associated with focal upregulation of cyclooxygenase-2, Bcl-2, and cell proliferation in the glandular epithelium. Prostate 2004, 61, 60–72. [Google Scholar] [CrossRef]
- Song, H.; Shen, Q.; Hu, S.; Jin, J. The role of macrophage migration inhibitory factor in promoting benign prostatic hyperplasia epithelial cell growth by modulating COX-2 and P53 signaling. Biol. Open 2020, 9, bio053447. [Google Scholar] [CrossRef]
- Ford, O.H., 3rd; Gregory, C.W.; Kim, D.; Smitherman, A.B.; Mohler, J.L. Androgen receptor gene amplification and protein expression in recurrent prostate cancer. J. Urol. 2003, 170, 1817–1821. [Google Scholar] [CrossRef]
- Palmberg, C.; Koivisto, P.; Kakkola, L.; Tammela, T.L.; Kallioniemi, O.P.; Visakorpi, T. Androgen receptor gene amplification at primary progression predicts response to combined androgen blockade as second line therapy for advanced prostate cancer. J. Urol. 2000, 164, 1992–1995. [Google Scholar] [CrossRef]
- Kumar, A.; Coleman, I.; Morrissey, C.; Zhang, X.; True, L.D.; Gulati, R.; Etzioni, R.; Bolouri, H.; Montgomery, B.; White, T.; et al. Substantial interindividual and limited intraindividual genomic diversity among tumors from men with metastatic prostate cancer. Nat. Med. 2016, 22, 369–378. [Google Scholar] [CrossRef] [PubMed]
- Dahiya, U.R.; Heemers, H.V. Analyzing the Androgen Receptor Interactome in Prostate Cancer: Implications for Therapeutic Intervention. Cells 2022, 11, 936. [Google Scholar] [CrossRef] [PubMed]
- Pomerantz, M.M.; Li, F.; Takeda, D.Y.; Lenci, R.; Chonkar, A.; Chabot, M.; Cejas, P.; Vazquez, F.; Cook, J.; Shivdasani, R.A.; et al. The androgen receptor cistrome is extensively reprogrammed in human prostate tumorigenesis. Nat. Genet. 2015, 47, 1346–1351. [Google Scholar] [CrossRef] [PubMed]
- McAllister, M.J.; Underwood, M.A.; Leung, H.Y.; Edwards, J. A review on the interactions between the tumor microenvironment and androgen receptor signaling in prostate cancer. Transl. Res. 2019, 206, 91–106. [Google Scholar] [CrossRef] [PubMed]
- Woenckhaus, J.; Fenic, I. Proliferative inflammatory atrophy: A background lesion of prostate cancer? Andrologia 2008, 40, 134–137. [Google Scholar] [CrossRef] [PubMed]
- Moreira, D.M.; de O Freitas, D.M.; Nickel, J.C.; Andriole, G.L.; Castro-Santamaria, R.; Freedland, S.J. The combination of histological prostate atrophy and inflammation is associated with lower risk of prostate cancer in biopsy specimens. Prostate Cancer Prostatic Dis. 2017, 20, 413–417. [Google Scholar] [CrossRef]
- Billis, A. Prostatic atrophy. Clinicopathological significance. Int. Braz. J. Urol. 2010, 36, 401–409. [Google Scholar] [CrossRef]
- Bastian, P.J.; Yegnasubramanian, S.; Palapattu, G.S.; Rogers, C.G.; Lin, X.; De Marzo, A.M.; Nelson, W.G. Molecular biomarker in prostate cancer: The role of CpG island hypermethylation. Eur. Urol. 2004, 46, 698–708. [Google Scholar] [CrossRef]
- Nakayama, M.; Bennett, C.J.; Hicks, J.L.; Epstein, J.I.; Platz, E.A.; Nelson, W.G.; De Marzo, A.M. Hypermethylation of the human glutathione S-transferase-pi gene (GSTP1) CpG island is present in a subset of proliferative inflammatory atrophy lesions but not in normal or hyperplastic epithelium of the prostate: A detailed study using laser-capture microdissection. Am. J. Pathol. 2003, 163, 923–933. [Google Scholar]
- Wagenlehner, F.M.; Elkahwaji, J.E.; Algaba, F.; Bjerklund-Johansen, T.; Naber, K.G.; Hartung, R.; Weidner, W. The role of inflammation and infection in the pathogenesis of prostate carcinoma. BJU Int. 2007, 100, 733–737. [Google Scholar] [CrossRef]
- Fang, L.Y.; Izumi, K.; Lai, K.P.; Liang, L.; Li, L.; Miyamoto, H.; Lin, W.J.; Chang, C. Infiltrating macrophages promote prostate tumorigenesis via modulating androgen receptor-mediated CCL4-STAT3 signaling. Cancer Res. 2013, 73, 5633–5646. [Google Scholar] [CrossRef] [PubMed]
- Garg, R.; Blando, J.M.; Perez, C.J.; Lal, P.; Feldman, M.D.; Smyth, E.M.; Ricciotti, E.; Grosser, T.; Benavides, F.; Kazanietz, M.G. COX-2 mediates pro-tumorigenic effects of PKCε in prostate cancer. Oncogene 2018, 37, 4735–4749. [Google Scholar] [CrossRef] [PubMed]
- Yao, C.; Li, G.; Cai, M.; Qian, Y.; Wang, L.; Xiao, L.; Thaiss, F.; Shi, B. Prostate cancer downregulated SIRP-α modulates apoptosis and proliferation through p38-MAPK/NF-κB/COX-2 signaling. Oncol. Lett. 2017, 13, 4995–5001. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Wang, J. Toll-Like Receptor 4 (TLR4)/Cyclooxygenase-2 (COX-2) Regulates Prostate Cancer Cell Proliferation, Migration, and Invasion by NF-κB Activation. Med. Sci. Monit. 2018, 24, 5588–5597. [Google Scholar] [CrossRef] [PubMed]
- Madrigal-Martínez, A.; Constâncio, V.; Lucio-Cazaña, F.J.; Fernández-Martínez, A.B. PROSTAGLANDIN E2 stimulates cancer-related phenotypes in prostate cancer PC3 cells through cyclooxygenase-2. J. Cell Physiol. 2019, 234, 7548–7559. [Google Scholar] [CrossRef]
- Pereira, V.S.; Alves, B.D.C.A.; Waisberg, J.; Fonseca, F.; Gehrke, F. Detection of COX-2 in liquid biopsy of patients with prostate cancer. J. Clin. Pathol. 2021. [Google Scholar] [CrossRef]
- Xu, X.-F.; Liu, G.-X.; Guo, Y.-S.; Zhu, H.-Y.; He, D.-L.; Qiao, X.-M.; Li, X.-H. Global, Regional, and National Incidence and Year Lived with Disability for Benign Prostatic Hyperplasia from 1990 to 2019. Am. J. Mens Health 2021, 15, 15579883211036786. [Google Scholar] [CrossRef]
- Madersbacher, S.; Sampson, N.; Culig, Z. Pathophysiology of Benign Prostatic Hyperplasia and Benign Prostatic Enlargement: A Mini-Review. Gerontology 2019, 65, 458–464. [Google Scholar] [CrossRef]
- Lepor, H. Pathophysiology of benign prostatic hyperplasia in the aging male population. Rev. Urol. 2005, 7 (Suppl. 4), S3–S12. [Google Scholar]
- La Vignera, S.; Condorelli, R.A.; Russo, G.I.; Morgia, G.; Calogero, A.E. Endocrine control of benign prostatic hyperplasia. Andrology 2016, 4, 404–411. [Google Scholar] [CrossRef]
- Miñana, B.; Molero, J.M.; Agra Rolán, A.; Martínez-Fornes, M.T.; Cuervo Pinto, R.; Lorite Mingot, D.; Carreño, Á.; Palacios-Moreno, J.M. Real-world therapeutic management and evolution of patients with benign prostatic hyperplasia in primary care and urology in Spain. Int. J. Clin. Pract. 2021, 75, e14250. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Cui, Y.; Wu, J.; Ding, R.; Cai, T.; Gao, Z. Meta-analysis of the efficacy and safety of combination of tamsulosin plus dutasteride compared with tamsulosin monotherapy in treating benign prostatic hyperplasia. BMC Urol. 2019, 19, 17. [Google Scholar] [CrossRef] [PubMed]
- Trost, L.; Saitz, T.R.; Hellstrom, W.J. Side Effects of 5-Alpha Reductase Inhibitors: A Comprehensive Review. Sex Med. Rev. 2013, 1, 24–41. [Google Scholar] [CrossRef] [PubMed]
- Sebastianelli, A.; Spatafora, P.; Morselli, S.; Vignozzi, L.; Serni, S.; McVary, K.T.; Kaplan, S.; Gravas, S.; Chapple, C.; Gacci, M. Tadalafil Alone or in Combination with Tamsulosin for the Management for LUTS/BPH and E.D. Curr. Urol. Rep. 2020, 21, 56. [Google Scholar] [CrossRef]
- Wang, Y.; Bao, Y.; Liu, J.; Duan, L.; Cui, Y. Tadalafil 5 mg Once Daily Improves Lower Urinary Tract Symptoms and Erectile Dysfunction: A Systematic Review and Meta-analysis. Low. Urin. Tract Symptoms 2018, 10, 84–92. [Google Scholar] [CrossRef]
- Rassweiler, J.; Teber, D.; Kuntz, R.; Hofmann, R. Complications of transurethral resection of the prostate (TURP)--incidence, management, and prevention. Eur. Urol. 2006, 50, 969–979. [Google Scholar] [CrossRef]
- Kramer, G.; Mitteregger, D.; Maj-Hes, A.; Sevchenco, S.; Brozek, W. Chronic inflammation as promotor and treatment target in benign prostate hyperplasia (BPH) and in prostate cancer. Urol. A 2007, 46, 1095–1096. [Google Scholar] [CrossRef]
- Paulis, G. Inflammatory mechanisms and oxidative stress in prostatitis: The possible role of antioxidant therapy. Res. Rep. Urol. 2018, 10, 75–87. [Google Scholar] [CrossRef]
- Cicero, A.F.G.; Allkanjari, O.; Busetto, G.M.; Cai, T.; Larganà, G.; Magri, V.; Perletti, G.; Robustelli Della Cuna, F.S.; Russo, G.I.; Stamatiou, K.; et al. Nutraceutical treatment and prevention of benign prostatic hyperplasia and prostate cancer. Arch. Ital. Urol. Androl. 2019, 91. [Google Scholar] [CrossRef]
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer Statistics, 2021. CA Cancer J. Clin. 2021, 71, 7–33. [Google Scholar] [CrossRef]
- Pienta, K.J.; Bradley, D. Mechanisms Underlying the Development of Androgen-Independent Prostate Cancer. Clin. Cancer Res. 2006, 12, 1665–1671. [Google Scholar] [CrossRef] [PubMed]
- Sharifi, N.; Gulley, J.L.; Dahut, W.L. Androgen Deprivation Therapy for Prostate Cancer. JAMA 2005, 294, 238–244. [Google Scholar] [CrossRef] [PubMed]
- Ortmann, O.; Weiss, J.M.; Diedrich, K. Gonadotrophin-releasing hormone (GnRH) and GnRH agonists: Mechanisms of action. Reprod. Biomed. Online 2002, 5 (Suppl. 1), 1–7. [Google Scholar] [CrossRef]
- Nguyen, P.L.; Alibhai, S.M.H.; Basaria, S.; D’Amico, A.V.; Kantoff, P.W.; Keating, N.L.; Penson, D.F.; Rosario, D.J.; Tombal, B.; Smith, M.R. Adverse Effects of Androgen Deprivation Therapy and Strategies to Mitigate Them. Eur. Urol. 2015, 67, 825–836. [Google Scholar] [CrossRef]
- Kumar, R.J.; Barqawi, A.; Crawford, E.D. Adverse events associated with hormonal therapy for prostate cancer. Rev. Urol. 2005, 7, S37–S43. [Google Scholar]
- Fizazi, K.; Tran, N.; Fein, L.; Matsubara, N.; Rodriguez-Antolin, A.; Alekseev, B.Y.; Özgüroğlu, M.; Ye, D.; Feyerabend, S.; Protheroe, A.; et al. Abiraterone plus Prednisone in Metastatic, Castration-Sensitive Prostate Cancer. N. Engl. J. Med. 2017, 377, 352–360. [Google Scholar] [CrossRef] [PubMed]
- Sternberg, C.N.; Fizazi, K.; Saad, F.; Shore, N.D.; De Giorgi, U.; Penson, D.F.; Ferreira, U.; Efstathiou, E.; Madziarska, K.; Kolinsky, M.P.; et al. Enzalutamide and Survival in Nonmetastatic, Castration-Resistant Prostate Cancer. N. Engl. J. Med. 2020, 382, 2197–2206. [Google Scholar] [CrossRef] [PubMed]
- Nakabayashi, M.; Sartor, O.; Jacobus, S.; Regan, M.M.; McKearn, D.; Ross, R.W.; Kantoff, P.W.; Taplin, M.E.; Oh, W.K. Response to docetaxel/carboplatin-based chemotherapy as first- and second-line therapy in patients with metastatic hormone-refractory prostate cancer. BJU Int. 2008, 101, 308–312. [Google Scholar] [CrossRef]
- Gravis, G.; Boher, J.-M.; Joly, F.; Soulié, M.; Albiges, L.; Priou, F.; Latorzeff, I.; Delva, R.; Krakowski, I.; Laguerre, B.; et al. Androgen Deprivation Therapy (ADT) Plus Docetaxel Versus ADT Alone in Metastatic Non castrate Prostate Cancer: Impact of Metastatic Burden and Long-term Survival Analysis of the Randomized Phase 3 GETUG-AFU15 Trial. Eur. Urol. 2016, 70, 256–262. [Google Scholar] [CrossRef]
- Baker, J.; Ajani, J.; Scotté, F.; Winther, D.; Martin, M.; Aapro, M.S.; von Minckwitz, G. Docetaxel-related side effects and their management. Eur. J. Oncol. Nurs. 2009, 13, 49–59. [Google Scholar] [CrossRef]
- Porter, C.M.; Shrestha, E.; Peiffer, L.B.; Sfanos, K.S. The microbiome in prostate inflammation and prostate cancer. Prostate Cancer Prostatic Dis. 2018, 21, 345–354. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Kim, J.; Bae, J.S. ROS homeostasis and metabolism: A critical liaison for cancer therapy. Exp. Mol. Med. 2016, 48, e269. [Google Scholar] [CrossRef] [PubMed]
- Panieri, E.; Santoro, M.M. ROS homeostasis and metabolism: A dangerous liason in cancer cells. Cell Death Dis. 2016, 7, e2253. [Google Scholar] [CrossRef]
- Jia, P.; Dai, C.; Cao, P.; Sun, D.; Ouyang, R.; Miao, Y. The role of reactive oxygen species in tumor treatment. RSC Adv. 2020, 10, 7740–7750. [Google Scholar] [CrossRef]
- Malvy, D.J.; Favier, A.; Faure, H.; Preziosi, P.; Galan, P.; Arnaud, J.; Roussel, A.M.; Briançon, S.; Hercberg, S. Effect of two years’ supplementation with natural antioxidants on vitamin and trace element status biomarkers: Preliminary data of the S.U.VI.MAX study. Cancer Detect. Prev. 2001, 25, 479–485. [Google Scholar] [PubMed]
- Meyer, F.; Galan, P.; Douville, P.; Bairati, I.; Kegle, P.; Bertrais, S.; Estaquio, C.; Hercberg, S. Antioxidant vitamin and mineral supplementation and prostate cancer prevention in the, S.U.VI.MAX trial. Int. J. Cancer 2005, 116, 182–186. [Google Scholar] [CrossRef]
- Zahra, K.F.; Lefter, R.; Ali, A.; Abdellah, E.C.; Trus, C.; Ciobica, A.; Timofte, D. The Involvement of the Oxidative Stress Status in Cancer Pathology: A Double View on the Role of the Antioxidants. Oxid. Med. Cell Longev. 2021, 2021, 9965916. [Google Scholar] [CrossRef]
- Norström, M.M.; Rådestad, E.; Sundberg, B.; Mattsson, J.; Henningsohn, L.; Levitsky, V.; Uhlin, M. Progression of benign prostatic hyperplasia is associated with pro-inflammatory mediators and chronic activation of prostate-infiltrating lymphocytes. Oncotarget 2016, 7, 23581–23593. [Google Scholar] [CrossRef]
- Kim, B.H.; Kim, C.I.; Chang, H.S.; Choe, M.S.; Jung, H.R.; Kim, D.Y.; Park, C.H. Cyclooxygenase-2 overexpression in chronic inflammation associated with benign prostatic hyperplasia: Is it related to apoptosis and angiogenesis of prostate cancer? Korean J. Urol. 2011, 52, 253–259. [Google Scholar] [CrossRef]
- Fiard, G.; Stavrinides, V.; Chambers, E.S.; Heavey, S.; Freeman, A.; Ball, R.; Akbar, A.N.; Emberton, M. Cellular senescence as a possible link between prostate diseases of the ageing male. Nat. Rev. Urol. 2021, 18, 597–610. [Google Scholar] [CrossRef]
- Srinivasan, D.; Senbanjo, L.; Majumdar, S.; Franklin, R.B.; Chellaiah, M.A. Androgen receptor expression reduces stemness characteristics of prostate cancer cells (PC3) by repression of CD44 and SOX2. J. Cell Biochem. 2018, 120, 2413–2428. [Google Scholar] [CrossRef] [PubMed]
- Calogero, A.E.; Burgio, G.; Condorelli, R.A.; Cannarella, R.; La Vignera, S. Epidemiology and risk factors of lower urinary tract symptoms/benign prostatic hyperplasia and erectile dysfunction. Aging Male 2019, 22, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Pizzol, D.; Xiao, T.; Smith, L.; Sánchez, G.F.L.; Garolla, A.; Parris, C.; Barnett, Y.; Ilie, P.C.; Soysal, P.; Shin, J.I.; et al. Prevalence of erectile dysfunction in male survivors of cancer: A systematic review and meta-analysis of cross-sectional studies. Br. J. Gen. Pract. 2021, 71, e372–e380. [Google Scholar] [CrossRef] [PubMed]
- Ryan, C.; Wefel, J.S.; Morgans, A.K. A review of prostate cancer treatment impact on the CNS and cognitive function. Prostate Cancer Prostatic Dis. 2020, 23, 207–219. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, B.D.; Jim, H.S.; Booth-Jones, M.; Small, B.J.; Sutton, S.K.; Lin, H.Y.; Park, J.Y.; Spiess, P.E.; Fishman, M.N.; Jacobsen, P.B. Course and Predictors of Cognitive Function in Patients With Prostate Cancer Receiving Androgen-Deprivation Therapy: A Controlled Comparison. J. Clin. Oncol. 2015, 33, 2021–2027. [Google Scholar] [CrossRef] [PubMed]
- Moyad, M.A. An introduction to aspirin, NSAIDs, and COX-2 inhibitors for the primary prevention of cardiovascular events and cancer and their potential preventive role in bladder carcinogenesis: Part, I. Semin. Urol. Oncol. 2001, 19, 294–305. [Google Scholar]
- Moyad, M.A. An introduction to aspirin, NSAids, and COX-2 inhibitors for the primary prevention of cardiovascular events and cancer and their potential preventive role in bladder carcinogenesis: Part, II. Semin. Urol. Oncol. 2001, 19, 306–316. [Google Scholar]
- Juszczak, K.; Drewa, T. The cardiovascular and gastrointestinal adverse effects of cyclooxygenase inhibitors seems to be a major concern that restricts their use in the treatment of urinary bladder dysfunction. Cent. Eur. J. Urol. 2015, 68, 57–59. [Google Scholar] [CrossRef]
- Di Francesco, S.; Castellan, P.; Manco, R.; Tenaglia, R.L. Reciprocal cross-talk between Prostaglandin E2 and bone in prostate cancer: A current review. Cent. Eur. J. Urol. 2011, 64, 201–204. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Origins | Leaves of apple trees |
| Category | Flavonoid; dihydrochalcones |
| Chemical structure | 3-(4-Hydroxyphenyl)-1-(2,4,6-trihydroxyphenyl)-1-propanone |
| Antioxidant activity | BDE (tested with B3LYP/6-311 + G(d,p) basis set in water): 79.36 kcal/mol The least value of IPs (tested with B3LYP/6-311 + G(d,p) basis set in water): 139.48 kcal/mol |
| General biological activities in the published literature | Anti-inflammatory ability; anti-oxidative ability; pro-apoptotic ability; anti-proliferative ability |
| PC3 | DU145 | LNCaP | |
|---|---|---|---|
| Cells’ viability (in MTT assay) | Decreased ↓ [13] | Decreased ↓ [13] | Decreased ↓ [13] |
| Cells’ viability (in CCK8 assay) | Decreased ↓ [13] | Decreased ↓ [13] | Decreased ↓ [13] |
| Cells’ proliferation (in Hoechst assay) | Decreased ↓ [15] | - | Decreased ↓ [15] |
| Biomarkers for cells’ proliferation | VEGF ↓ [13]; CCND1 ↓ [13]; CCNB1 ↓ [13] | - | VEGF ↓ [13]; CCND1 ↓ [13]; CCNB1 ↓ [13]; |
| Reactive oxygen species | Increased [12] | Increased [12] | - |
| Oxidative stress | Increased [12] | Increased [12] | - |
| Biomarkers of antioxidant enzymes | Catalase ↓ [12]; SOD2 ↓ [12]; GPx1 ↓ [12]; GPx3 ↓ [12] | Catalase ↓ [12]; SOD2 ↓ [12]; GPx1 ↓ [12]; GPx3 ↓ [12] | - |
| Biomarkers of wnt/β-catenin signaling pathway | β-catenin ↓ [12]; TCF4 ↓ [12]; FoxA2 ↓ [12]; c-Myc ↓ [12] | β-catenin ↓ [12]; TCF4 ↓ [12]; FoxA2 ↓ [12]; c-Myc ↓ [12] | - |
| Biomarkers of extrinsic apoptosis | Caspase3/8 ↑ [13]; PARP-1 ↑ [13] | - | TRAIL ↑ [14]; Caspase3/8 ↑ [13]; PARP-1 ↑ [13] |
| Biomarkers of intrinsic apoptosis | Caspase3/9 ↑ [13]; Bax ↑ [13]; Bcl2 ↓ [13]; XIAP ↓ [13]; Survivin ↓ [13]; | - | Caspase3/9 ↑ [13]; Bax ↑ [13]; Bcl2 ↓ [13]; XIAP ↓ [13]; Survivin ↓ [13]; |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, C.-H.; Ou, Y.-C.; Lin, C.-C.; Lin, Y.-S.; Tung, M.-C.; Yu, C.-C.; Lin, J.-T.; Wen, C.-Y. Phloretin in Benign Prostate Hyperplasia and Prostate Cancer: A Contemporary Systematic Review. Life 2022, 12, 1029. https://doi.org/10.3390/life12071029
Yang C-H, Ou Y-C, Lin C-C, Lin Y-S, Tung M-C, Yu C-C, Lin J-T, Wen C-Y. Phloretin in Benign Prostate Hyperplasia and Prostate Cancer: A Contemporary Systematic Review. Life. 2022; 12(7):1029. https://doi.org/10.3390/life12071029
Chicago/Turabian StyleYang, Che-Hsueh, Yen-Chuan Ou, Chi-Chien Lin, Yi-Sheng Lin, Min-Che Tung, Chia-Cheng Yu, Jen-Tai Lin, and Chen-Yueh Wen. 2022. "Phloretin in Benign Prostate Hyperplasia and Prostate Cancer: A Contemporary Systematic Review" Life 12, no. 7: 1029. https://doi.org/10.3390/life12071029
APA StyleYang, C.-H., Ou, Y.-C., Lin, C.-C., Lin, Y.-S., Tung, M.-C., Yu, C.-C., Lin, J.-T., & Wen, C.-Y. (2022). Phloretin in Benign Prostate Hyperplasia and Prostate Cancer: A Contemporary Systematic Review. Life, 12(7), 1029. https://doi.org/10.3390/life12071029