
Searching for Life, Mindful of Lyfe’s Possibilities


Abstract
:1. How One Defines or Characterizes Life Is Critical to Logical and Lucid Biosignatures Research
2. A Better Understanding of the Origins of Life Is Important to Biosignatures Research
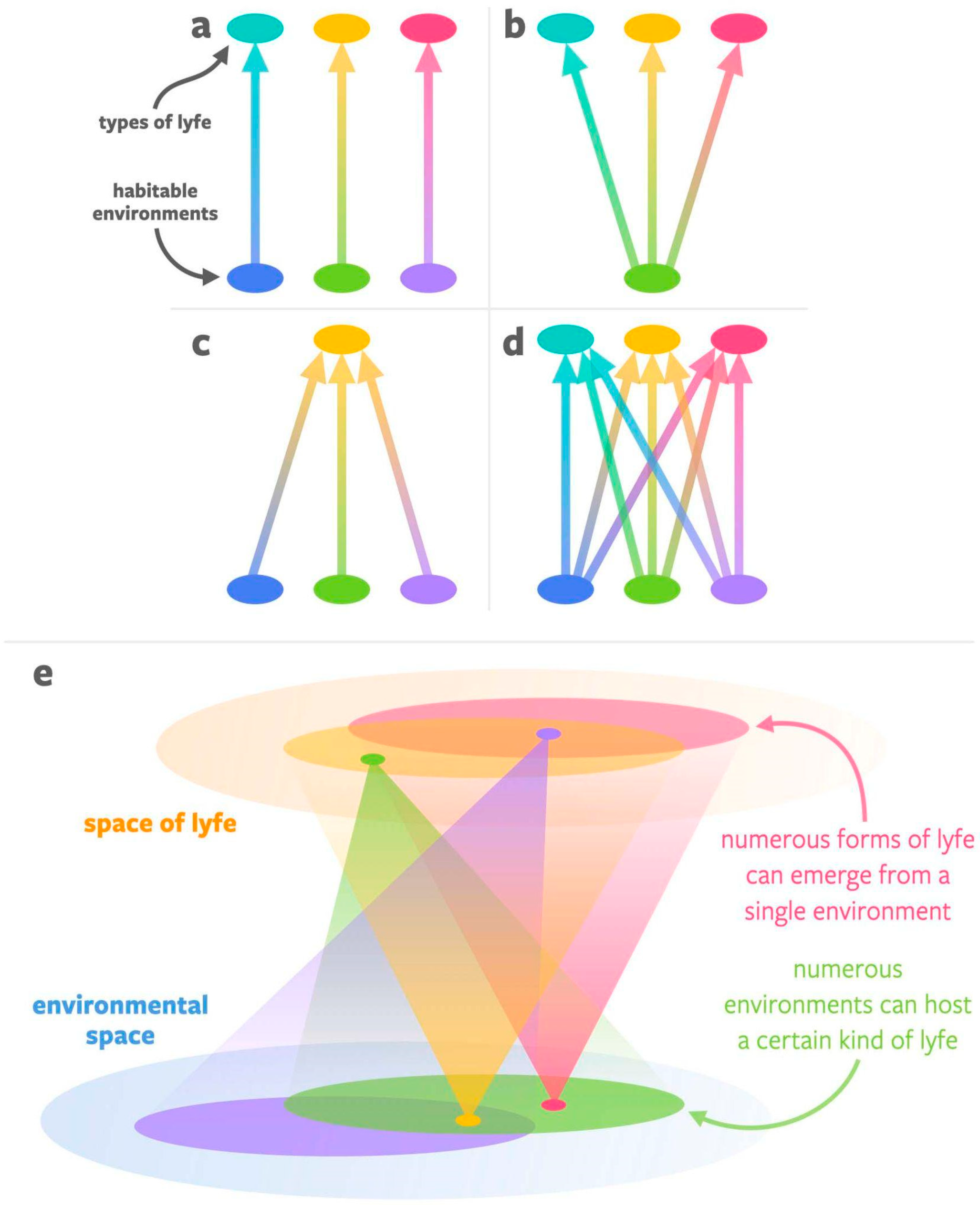
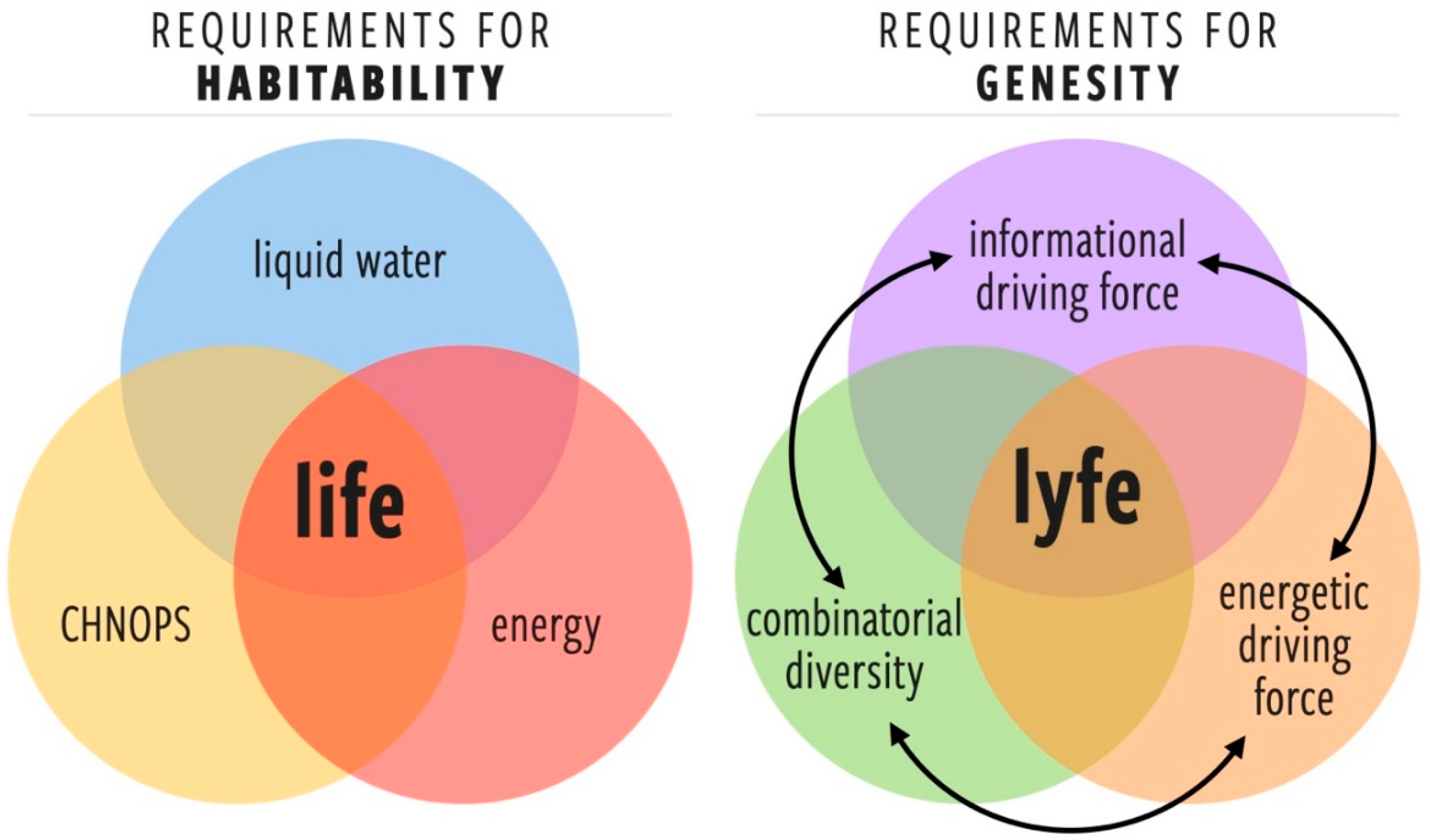
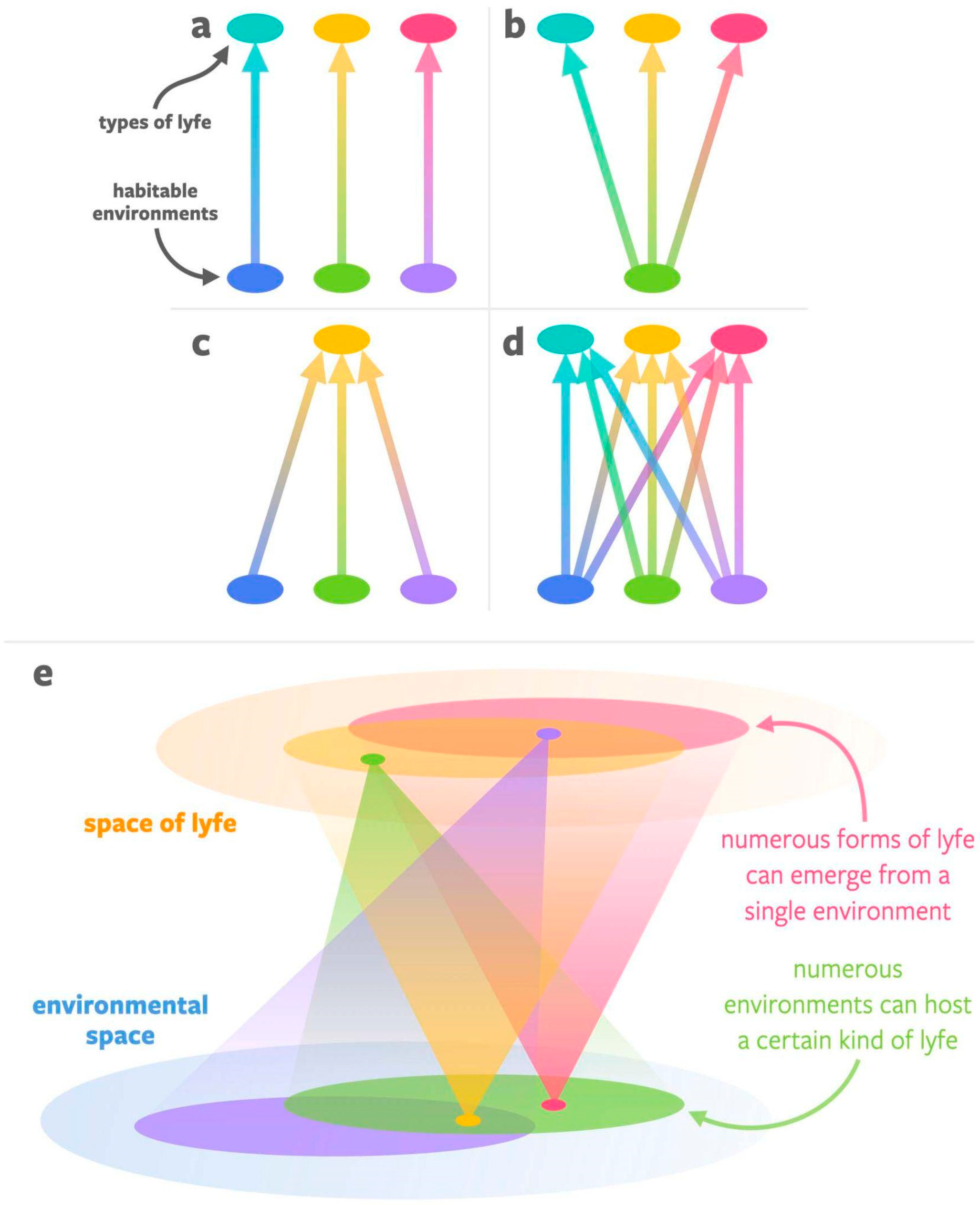
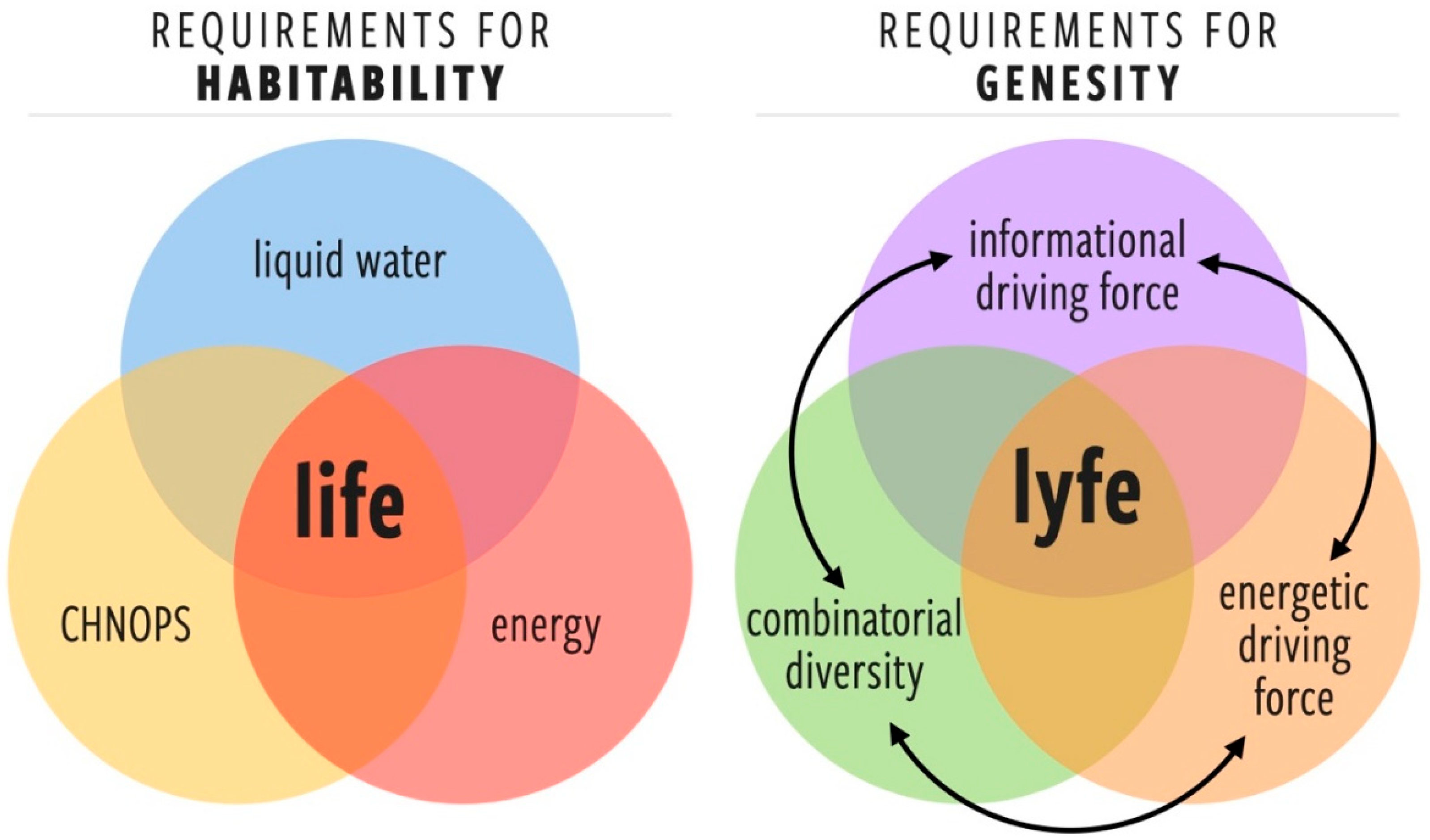
3. Developing a More General Concept of Habitability (Genesity) Is Important to Biosignatures Research
3.1. Proposed Criteria for Genesity, a More General Concept of Habitability
- Energetic driving force (EDF)A requisite set of one or more free energy gradients that provide a sufficient thermodynamic force for the fluxes necessary for starting, maintaining, and complexifying lyfe. Note the additional requirement that such gradients cannot be trivially dissipated by abiotic processes (in such a case, the abiotic channels would become the paths of least resistance).
- 2.
- Informational driving force (IDF)The environment must exhibit a threshold level of complexity such that there is a selection pressure for the information processing and learning behaviors that are at the core of the living state [22,23,24,25,26,27]. There is no physical or computational reason for a lyfeform to emerge or evolve in an environment that is trivially simple. The IDF highlights an important difference between habitability and genesity: a perfectly placid pool of organic broth may be an optimally habitable environment, but it would have a negligible IDF and, therefore, low genesity.
- 3.
- Combinatorial diversity of components (CD)A sufficiently diverse set of components (molecular or otherwise) is required for the emergence of prebiotic systems that can harness the EDF and find dynamical robustness in resolving the information gradients provided by the IDF. The minimal set of such components or their chemical identity is not yet known, but we can surmise that for a given environment, there is a minimal component set required for the emergence of the lowest levels of biological complexity.
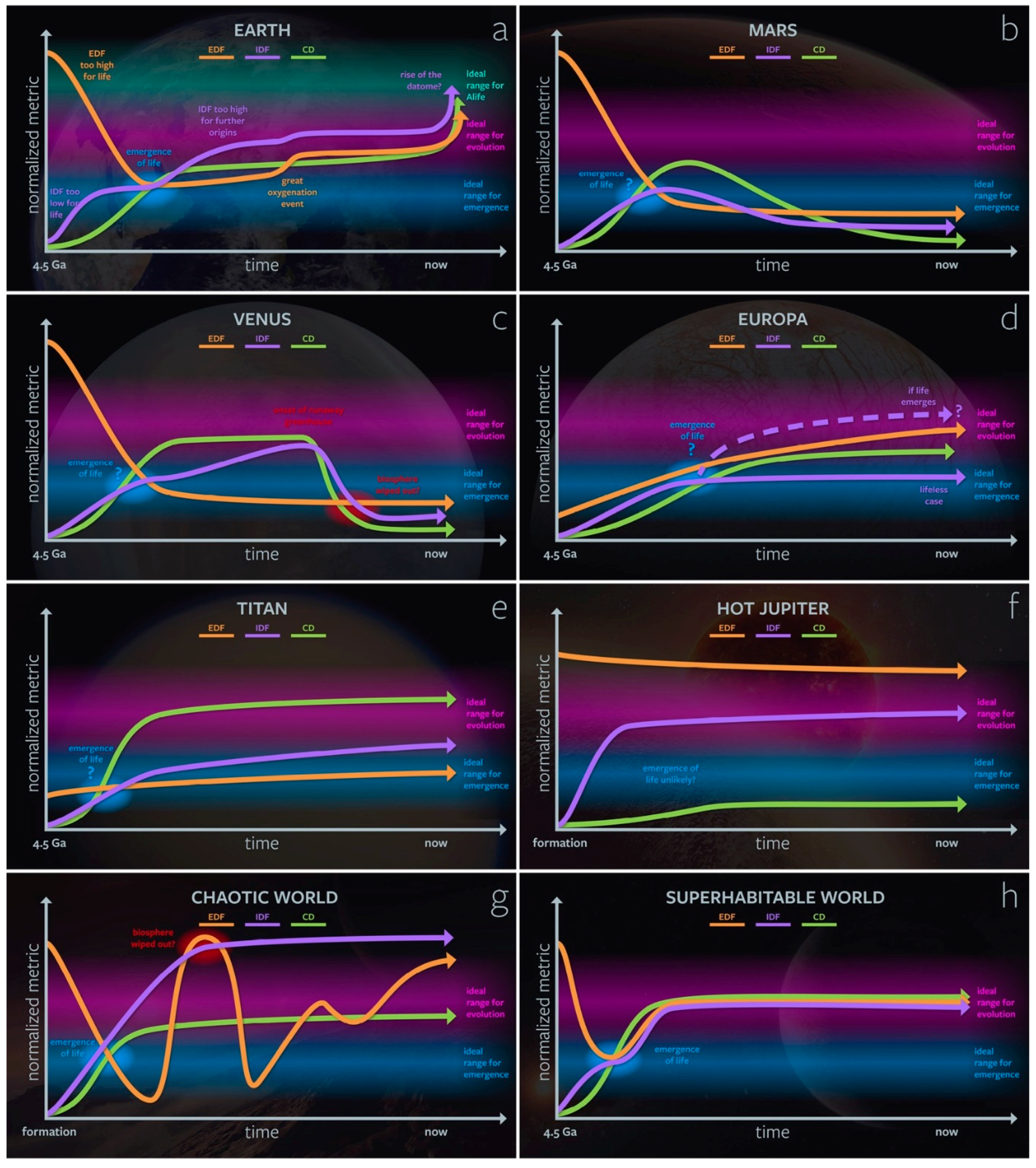
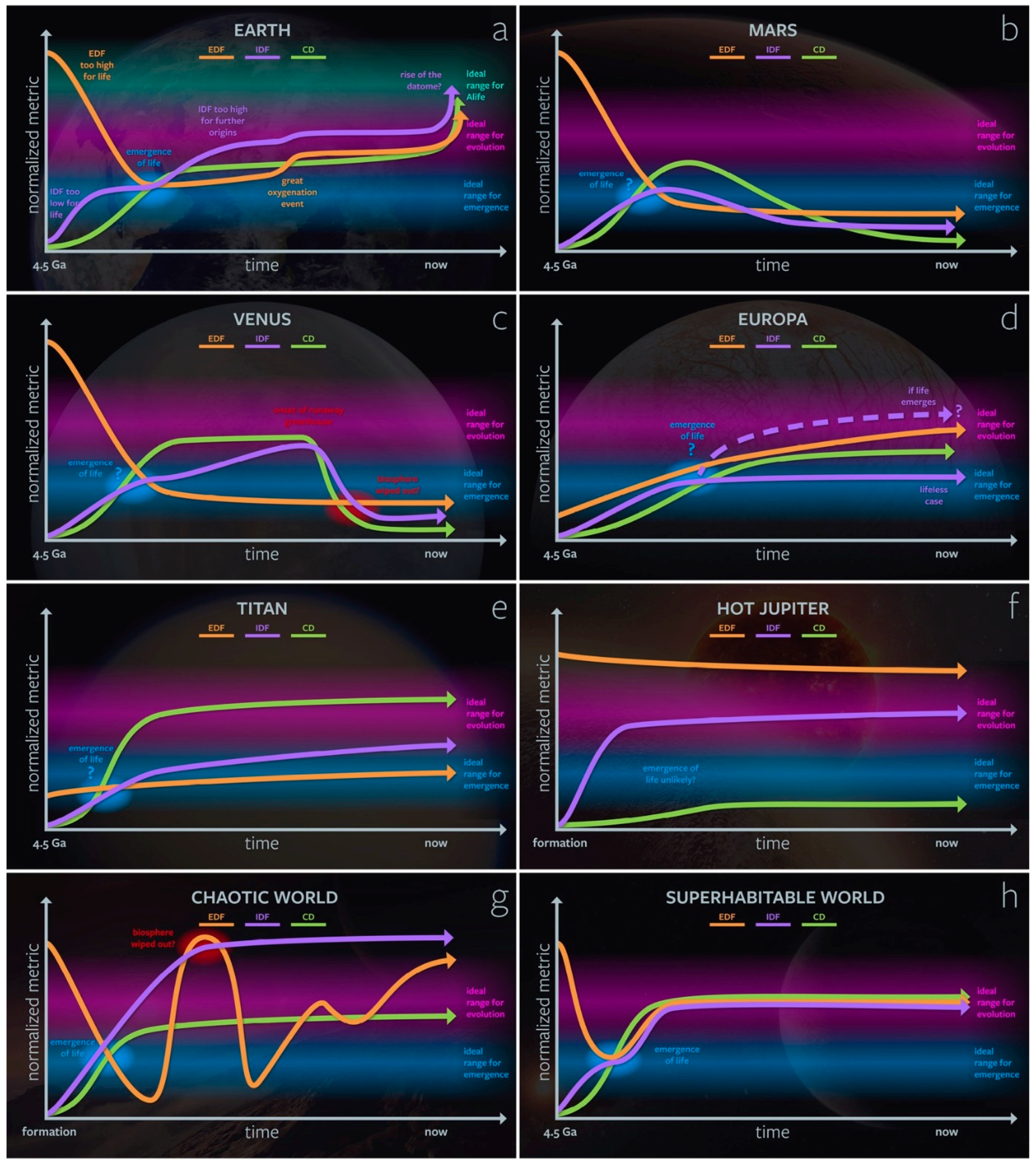
3.2. Genesity Parameter Evolution and Co-Dependence on Planetary Context
3.3. The Genesity of Cyberspace and the Dataome
3.4. Toward the Quantification of Gensity Parameters for Biosignature Science
4. Concluding Remarks
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cleland, C. The Quest for a Universal Theory of Life: Searching for Life as We Don’t Know It; Cambridge University Press: Cambridge, UK, 2019. [Google Scholar]
- Cleland, C.E.; Chyba, C.F. Defining ‘life’. Orig. Life Evol. Biosph. 2002, 32, 387–393. [Google Scholar] [CrossRef] [PubMed]
- Bartlett, S.; Wong, M.L. Defining Lyfe in the Universe: From Three Privileged Functions to Four Pillars. Life 2020, 10, 42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cockell, C.S.; Higgins, P.M.; Johnstone, A.A. Biologically Available Chemical Energy in the Temperate but Uninhabitable Venusian Cloud Layer: What Do We Want to Know? Astrobiology 2021, 21, 1224–1236. [Google Scholar] [CrossRef] [PubMed]
- Krissansen-Totton, J.; Bergsman, D.S.; Catling, D.C. On detecting biospheres from chemical thermodynamic disequilibrium in planetary atmospheres. Astrobiology 2016, 16, 39–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lovelock, J.E. A Physical Basis for Life Detection Experiments. Nature 1965, 207, 568–570. [Google Scholar] [CrossRef] [PubMed]
- Kempes, C.P.; Follows, M.J.; Smith, H.; Graham, H.; House, C.H.; Levin, S.A. Generalized Stoichiometry and Bio-geochemistry for Astrobiological Applications. Bull. Math. Biol. 2021, 83, 73. [Google Scholar] [CrossRef]
- Marshall, S.M.; Mathis, C.; Carrick, E.; Keenan, G.; Cooper, G.J.T.; Graham, H.; Craven, M.; Gromski, P.S.; Moore, D.G.; Walker, S.I.; et al. Identifying molecules as biosignatures with assembly theory and mass spectrometry. Nat. Commun. 2021, 12, 3033. [Google Scholar] [CrossRef]
- Bartlett, S.; Li, J.; Gu, L.; Sinapayen, L.; Fan, S.; Natraj, V.; Jiang, J.H.; Crisp, D.; Yung, Y.L. Assessing planetary complexity and potential agnostic biosignatures using epsilon machines. Nat. Astron. 2022, 6, 387–392. [Google Scholar] [CrossRef]
- Catling David, C.; Kiang Nancy, Y.; Robinson Tyler, D.; Rushby Andrew, J.; Genio Anthony, D. Exoplanet biosignatures: A framework for their assessment. Astrobiology 2018, 18, 709–738. [Google Scholar] [CrossRef]
- Walker, S.I.; Bains, W.; Cronin, L.; DasSarma, S.; Danielache, S.; Domagal-Goldman, S.; Kacar, B.; Kiang, N.Y.; Lenardic, A.; Reinhard, C.T.; et al. Exoplanet Biosignatures: Future Directions. Astrobiology 2018, 18, 779–824. [Google Scholar] [CrossRef]
- Affholder, A.; Guyot, F.; Sauterey, B.; Ferrière, R.; Mazevet, S. Bayesian analysis of Enceladus’s plume data to assess methanogenesis. Nat. Astron. 2021, 5, 805–814. [Google Scholar] [CrossRef]
- Ehlmann, B.L.; Anderson, F.S.; Andrews-Hanna, J.; Catling, D.C.; Christensen, P.R.; Cohen, B.A.; Dressing, C.D.; Edwards, C.S.; Elkins-Tanton, L.T.; Farley, K.A.; et al. The sustainability of habitability on terrestrial planets: Insights, questions, and needed measurements from Mars for understanding the evolution of Earth-like worlds. J. Geophys. Res. Planets 2016, 121, 1927–1961. [Google Scholar] [CrossRef]
- Damer, B. A Field Trip to the Archaean in Search of Darwin’s Warm Little Pond. Life 2016, 6, 21. [Google Scholar] [CrossRef] [Green Version]
- Damer, B.; Deamer, D. The Hot Spring Hypothesis for an Origin of Life. Astrobiology 2020, 20, 429–452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, W.; Russell, M.J. On the origin of biochemistry at an alkaline hydrothermal vent. Philos. Trans. R. Soc. B Biol. Sci. 2007, 362, 1887–1926. [Google Scholar] [CrossRef] [PubMed]
- Pearce, B.K.D.; Pudritz, R.E.; Semenov, D.A.; Henning, T.K. Origin of the RNA world: The fate of nucleobases in warm little ponds. Proc. Natl. Acad. Sci. USA 2017, 114, 11327–11332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barge, L.M.; Rodriguez, L.E.; Weber, J.M.; Theiling, B.P. Determining the “Biosignature Threshold” for Life Detection on Biotic, Abiotic, or Prebiotic Worlds. Astrobiology 2021, 22, 481–493. [Google Scholar] [CrossRef]
- Preiner, M.; Asche, S.; Becker, S.; Betts, H.C.; Boniface, A.; Camprubi, E.; Chandru, K.; Erastova, V.; Garg, S.G.; Khawaja, N.; et al. The Future of Origin of Life Research: Bridging Decades-Old Divisions. Life 2020, 10, 20. [Google Scholar] [CrossRef] [Green Version]
- Kasting, J.F.; Whitmire, D.P.; Reynolds, R.T. Habitable Zones around Main Sequence Stars. Icarus 1993, 101, 108–128. [Google Scholar] [CrossRef]
- Merino, N.; Aronson, H.S.; Bojanova, D.P.; Feyhl-Buska, J.; Wong, M.L.; Zhang, S.; Giovannelli, D. Living at the extremes: Extremophiles and the limits of life in a planetary context. Front. Microbio. 2019, 10, 780. [Google Scholar] [CrossRef] [Green Version]
- Baluška, F.; Levin, M. On Having No Head: Cognition throughout Biological Systems. Front. Psychol. 2016, 7, 902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davies, P.C.W.; Walker, S.I. The hidden simplicity of biology. Rep. Prog. Phys. 2016, 79, 102601. [Google Scholar] [CrossRef] [PubMed]
- Farnsworth, K.D.; Nelson, J.; Gershenson, C. Living is Information Processing: From Molecules to Global Systems. Acta Biotheor. 2013, 61, 203–222. [Google Scholar] [CrossRef] [PubMed]
- Tkačik, G.; Bialek, W. Information processing in living systems. Annu. Rev. Condens. Matter Phys. 2016, 7, 89–117. [Google Scholar] [CrossRef] [Green Version]
- Walker, S.I.; Kim, H.; Davies, P.C. The informational architecture of the cell. Philos. Trans. R. Soc. A Math. Phys. Eng. Sci. 2016, 374, 20150057. [Google Scholar] [CrossRef]
- Witzany, G. What is life? Front. Astron. Space Sci. 2020, 7, 7. [Google Scholar] [CrossRef] [Green Version]
- Crutchfield, J.P. Between order and chaos. Nat. Phys. 2012, 8, 17–24. [Google Scholar] [CrossRef]
- Langton, C.G. Computation at the edge of chaos: Phase transitions and emergent computation. Phys. D Nonlinear Phenom. 1990, 42, 12–37. [Google Scholar] [CrossRef] [Green Version]
- Hazen, R.M. The emergence of patterning in lifes origin and evolution. Int. J. Dev. Biol. 2009, 53, 683–692. [Google Scholar] [CrossRef] [Green Version]
- Hazen, R.M.; Eldredge, N. Themes and Variations in Complex Systems. Elements 2010, 6, 43–46. [Google Scholar] [CrossRef]
- Lyons, T.W.; Reinhard, C.T.; Planavsky, N.J. The rise of oxygen in Earth’s early ocean and atmosphere. Nature 2014, 506, 307–315. [Google Scholar] [CrossRef] [PubMed]
- Schirrmeister, B.E.; de Vos, J.M.; Antonelli, A.; Bagheri, H.C. Evolution of multicellularity coincided with increased diversification of cyanobacteria and the Great Oxidation Event. Proc. Natl. Acad. Sci. USA 2013, 110, 1791–1796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reinhard, C.T.; Planavsky, N.J.; Olson, S.L.; Lyons, T.W.; Erwin, D.H. Earth’s oxygen cycle and the evolution of animal life. Proc. Natl. Acad. Sci. USA 2016, 113, 8933–8938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolbert, E. The Sixth Extinction: An Unnatural History; A&C Black: London, UK, 2014. [Google Scholar]
- Adams, D.; Luo, Y.; Wong, M.L.; Dunn, P.; Christensen, M.; Dong, C.; Hu, R.; Yung, Y. Nitrogen Fixation at Early Mars. Astrobiology 2021, 21, 968–980. [Google Scholar] [CrossRef]
- Clark, B.; Kolb, V.; Steele, A.; House, C.; Lanza, N.; Gasda, P.; VanBommel, S.; Newsom, H.; Martínez-Frías, J. Origin of Life on Mars: Suitability and Opportunities. Life 2021, 11, 539. [Google Scholar] [CrossRef]
- Fornaro, T.; Steele, A.; Brucato, J.R. Catalytic/Protective Properties of Martian Minerals and Implications for Possible Origin of Life on Mars. Life 2018, 8, 56. [Google Scholar] [CrossRef] [Green Version]
- Ingersoll, A.P. The runaway greenhouse: A history of water on Venus. J. Atmos. Sci. 1969, 26, 1191–1198. [Google Scholar] [CrossRef] [Green Version]
- Mogul, R.; Limaye, S.S.; Lee, Y.J.; Pasillas, M. Potential for Phototrophy in Venus’ Clouds. Astrobiology 2021, 21, 1237–1249. [Google Scholar] [CrossRef]
- Chyba, C.F. Energy for microbial life on Europa. Nature 2000, 403, 381–382. [Google Scholar] [CrossRef]
- Gomez-Buckley, A.C.; Showalter, G.M.; Wong, M.L. Modeling Virus and Bacteria Populations in Europa’s Subsurface Ocean. Life 2022, 12, 620. [Google Scholar] [CrossRef]
- Greenberg, R. Transport rates of radiolytic substances into Europa’s ocean: Implications for the potential origin and maintenance of life. Astrobiology 2010, 10, 275–283. [Google Scholar] [CrossRef] [PubMed]
- Hand, K.P.; Carlson, R.W.; Chyba, C.F. Energy, Chemical Disequilibrium, and Geological Constraints on Europa. Astrobiology 2007, 7, 1006–1022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Gudipati, M.S.; Mishra, Y.N.; Liang, M.-C.; Yung, Y.L. Oxidant generation in the ice under electron irradiation: Simulation and application to Europa. Icarus 2021, 373, 114760. [Google Scholar] [CrossRef]
- Vance, S.D.; Hand, K.P.; Pappalardo, R.T. Geophysical controls of chemical disequilibria in Europa. Geophys. Res. Lett. 2016, 43, 4871–4879. [Google Scholar] [CrossRef]
- Willacy, K.; Allen, M.; Yung, Y. A New Astrobiological Model of The Atmosphere of Titan. Astrophys. J. Lett. 2016, 829, 79. [Google Scholar] [CrossRef] [Green Version]
- Frank, A.; Grinspsoon, D.; Walker, S. Intelligence as a planetary scale process. Int. J. Astrobiol. 2022, 21, 47–61. [Google Scholar] [CrossRef]
- Wong, M.L.; Bartlett, S. Asymptotic Burnout & Homeostatic Awakening: A Possible Solution to the Fermi Paradox? J. R. Soc. Interface 2022, 19, 20220029. [Google Scholar] [CrossRef]
- Domagal-Goldman, S.D.; Segura, A.; Claire, M.W.; Robinson, T.D.; Meadows, V.S. Abiotic Ozone and Oxygen in Atmospheres Similar to Prebiotic Earth. Astrophys. J. Lett. 2014, 792, 90. [Google Scholar] [CrossRef] [Green Version]
- Gao, P.; Hu, R.; Robinson, T.D.; Li, C.; Yung, Y.L. Stability of CO2 Atmospheres on Desiccated M Dwarf Exoplanets. Astrophys. J. Lett. 2015, 806, 249. [Google Scholar] [CrossRef] [Green Version]
- Wordsworth, R.; Pierrehumbert, R. Abiotic Oxygen-Dominated Atmospheres on Terrestrial Habitable Zone Planets. Astrophys. J. Lett. 2014, 785, L20. [Google Scholar] [CrossRef] [Green Version]
- Ward, L.M.; Stamenković, V.; Hand, K.; Fischer, W.W. Follow the oxygen: Comparative histories of planetary oxygenation and opportunities for aerobic life. Astrobiology 2019, 19, 811–824. [Google Scholar] [CrossRef] [PubMed]
- Adams, A.M.; Berner, A.; Davies, P.C.W.; Walker, S.I. Physical Universality, State-Dependent Dynamical Laws and Open-Ended Novelty. Entropy 2017, 19, 461. [Google Scholar] [CrossRef] [Green Version]
- Banzhaf, W.; Baumgaertner, B.; Beslon, G.; Doursat, R.; Foster, J.A.; McMullin, B.; De Melo, V.V.; Miconi, T.; Spector, L.; Stepney, S.; et al. Defining and simulating open-ended novelty: Requirements, guidelines, and challenges. Theory Biosci. 2016, 135, 131–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guttenberg, N.; Virgo, N.; Penn, A. On the Potential for Open-Endedness in Neural Networks. Artif. Life 2019, 25, 145–167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Markovitch, O.; Sorek, D.; Lui, L.T.; Lancet, D.; Krasnogor, N. Is There an Optimal Level of Open-Endedness in Prebiotic Evolution? Orig. Life Evol. Biosphere 2012, 42, 469–474. [Google Scholar] [CrossRef] [PubMed]
- Pattee, H.H.; Sayama, H. Evolved Open-Endedness, Not Open-Ended Evolution. Artif. Life 2019, 25, 4–8. [Google Scholar] [CrossRef]
- Ruiz-Mirazo, K.; Pereto, J.; Moreno, A. A Universal Definition of Life: Autonomy and Open-Ended Evolution. Orig. Life Evol. Biosphere 2004, 34, 323–346. [Google Scholar] [CrossRef]
- Stanley, K.O.; Miikkulainen, R. Competitive Coevolution through Evolutionary Complexification. J. Artif. Intell. Res. 2004, 21, 63–100. [Google Scholar] [CrossRef]
- Adams, A.M.; Zenil, H.; Davies, P.C.W.; Walker, S.I. Formal Definitions of Unbounded Evolution and Innovation Reveal Universal Mechanisms for Open-Ended Evolution in Dynamical Systems. Sci. Rep. 2017, 7, 997. [Google Scholar] [CrossRef]
- Taylor, T. Evolutionary Innovations and Where to Find Them: Routes to Open-Ended Evolution in Natural and Artificial Systems. Artif. Life 2019, 25, 207–224. [Google Scholar] [CrossRef] [Green Version]
- Chopra, A.; Lineweaver, C. The Case for a Gaian Bottleneck: The Biology of Habitability. Astrobiology 2016, 16, 7–22. [Google Scholar] [CrossRef] [PubMed]
- Lenton, T.M.; Daines, S.J.; Dyke, J.G.; Nicholson, A.E.; Wilkinson, D.M.; Williams, H.T. Selection for Gaia across Multiple Scales. Trends Ecol. Evol. 2018, 33, 633–645. [Google Scholar] [CrossRef] [PubMed]
- Nicholson, A.E.; Wilkinson, D.M.; Williams, H.T.; Lenton, T.M. Gaian bottlenecks and planetary habitability maintained by evolving model biospheres: The ExoGaia model. Mon. Not. R. Astron. Soc. 2018, 477, 727–740. [Google Scholar] [CrossRef] [Green Version]
- Haglich, P.; Rouff, C.; Pullum, L. Detecting Emergence in Social Networks. In Proceedings of the 2010 IEEE Second International Conference on Social Computing, Minneapolis, MN, USA, 20–22 August 2010; pp. 693–696. [Google Scholar] [CrossRef]
- Kempes, C.P.; Krakauer, D.C. The Multiple Paths to Multiple Life. J. Mol. Evol. 2021, 89, 415–426. [Google Scholar] [CrossRef] [PubMed]
- Scharf, C. The Ascent of Information; Penguin Publishing Group: New York, NY, USA, 2021. [Google Scholar]
- Bartlett, S.J.; Beckett, P. Probing complexity: Thermodynamics and computational mechanics approaches to origins studies. Interface Focus 2019, 9, 20190058. [Google Scholar] [CrossRef]
- Crutchfield, J.P. The calculi of emergence: Computation, dynamics and induction. Phys. D Nonlinear Phenom. 1994, 75, 11–54. [Google Scholar] [CrossRef]
- Ashby, W.R. Requisite Variety and Its Implications for the Control of Complex Systems. In Facets of Systems Science; Springer: Boston, MA, USA, 1991; pp. 405–417. [Google Scholar] [CrossRef]
- Johnson, S.S.; Anslyn, E.V.; Graham, H.V.; Mahaffy, P.R.; Ellington, A.D. Fingerprinting Non-Terran Biosignatures. Astrobiology 2018, 18, 915–922. [Google Scholar] [CrossRef]
- Ray, C.; Glein, C.R.; Waite, J.H.; Teolis, B.; Hoehler, T.; Huber, J.A.; Lunine, J.; Postberg, F. Oxidation processes diversify the metabolic menu on Enceladus. Icarus 2020, 364, 114248. [Google Scholar] [CrossRef]
- Meadows, V.S.; Graham, H.; Abrahamsson, V.; Adam, Z.; Amador-French, E.; Arney, G.; Barge, L.; Barlow, E.; Berea, A.; Bose, M.; et al. Community Report From the Biosignatures Standards of Evidence Workshop. 2022. Available online: https://www.nfold.org/_files/ugd/c2389f_d081d7f5fcfc455cbe0bb560a900ecb6.pdf (accessed on 21 February 2022).
- Green, J.; Hoehler, T.; Neveu, M.; Domagal-Goldman, S.; Scalice, D.; Voytek, M. Call for a framework for reporting evidence for life beyond Earth. Nature 2021, 598, 575–579. [Google Scholar] [CrossRef]
- Guttenberg, N.; Chen, H.; Mochizuki, T.; Cleaves, H.J. Classification of the biogenicity of complex organic mixtures for the detection of extraterrestrial life. Life 2021, 11, 234. [Google Scholar] [CrossRef]
- Stevenson, D.J. Commentary: The habitability mantra: Hunting the Snark. Phys. Today 2018, 71, 10–12. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
{kind=link}
{kind=link}
| Parameter | Too Low | Too High |
|---|---|---|
| Energetic driving force | Unable to supply work needed to maintain complexity, growth, and innovation | Turbulence; immediately overcomes kinetic barriers, producing chaotic behavior |
| Informational driving force | No incentive for evolution; no need for a living system | Prohibitively complex environment requiring an impossibly sophisticated first, putative learning system |
| Combinatorial diversity | Set of available components cannot produce systems above biological complexity threshold | Component configuration space too random/dispersed for biological emergence |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wong, M.L.; Bartlett, S.; Chen, S.; Tierney, L. Searching for Life, Mindful of Lyfe’s Possibilities. Life 2022, 12, 783. https://doi.org/10.3390/life12060783
Wong ML, Bartlett S, Chen S, Tierney L. Searching for Life, Mindful of Lyfe’s Possibilities. Life. 2022; 12(6):783. https://doi.org/10.3390/life12060783
Chicago/Turabian StyleWong, Michael L., Stuart Bartlett, Sihe Chen, and Louisa Tierney. 2022. "Searching for Life, Mindful of Lyfe’s Possibilities" Life 12, no. 6: 783. https://doi.org/10.3390/life12060783
APA StyleWong, M. L., Bartlett, S., Chen, S., & Tierney, L. (2022). Searching for Life, Mindful of Lyfe’s Possibilities. Life, 12(6), 783. https://doi.org/10.3390/life12060783