
Membrane-Free Stem Cell Extract Enhances Blood–Brain Barrier Integrity by Suppressing NF-κB-Mediated Activation of NLRP3 Inflammasome in Mice with Ischemic Stroke

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of MFSCE
2.2. Mouse Model of Focal Cerebral Ischemia and Treatment with MFSCE
2.3. Infarct Volume and Edema
2.4. Neurological Deficit Score
2.5. Wire-Grip Test
2.6. Evans Blue Extravasation and Water Content
2.7. Real-Time Polymerase Chain Reaction
2.8. Western Blotting
2.9. Histological Examination
2.10. In Vivo Safety Evaluation
2.11. Statistical Analysis
3. Results
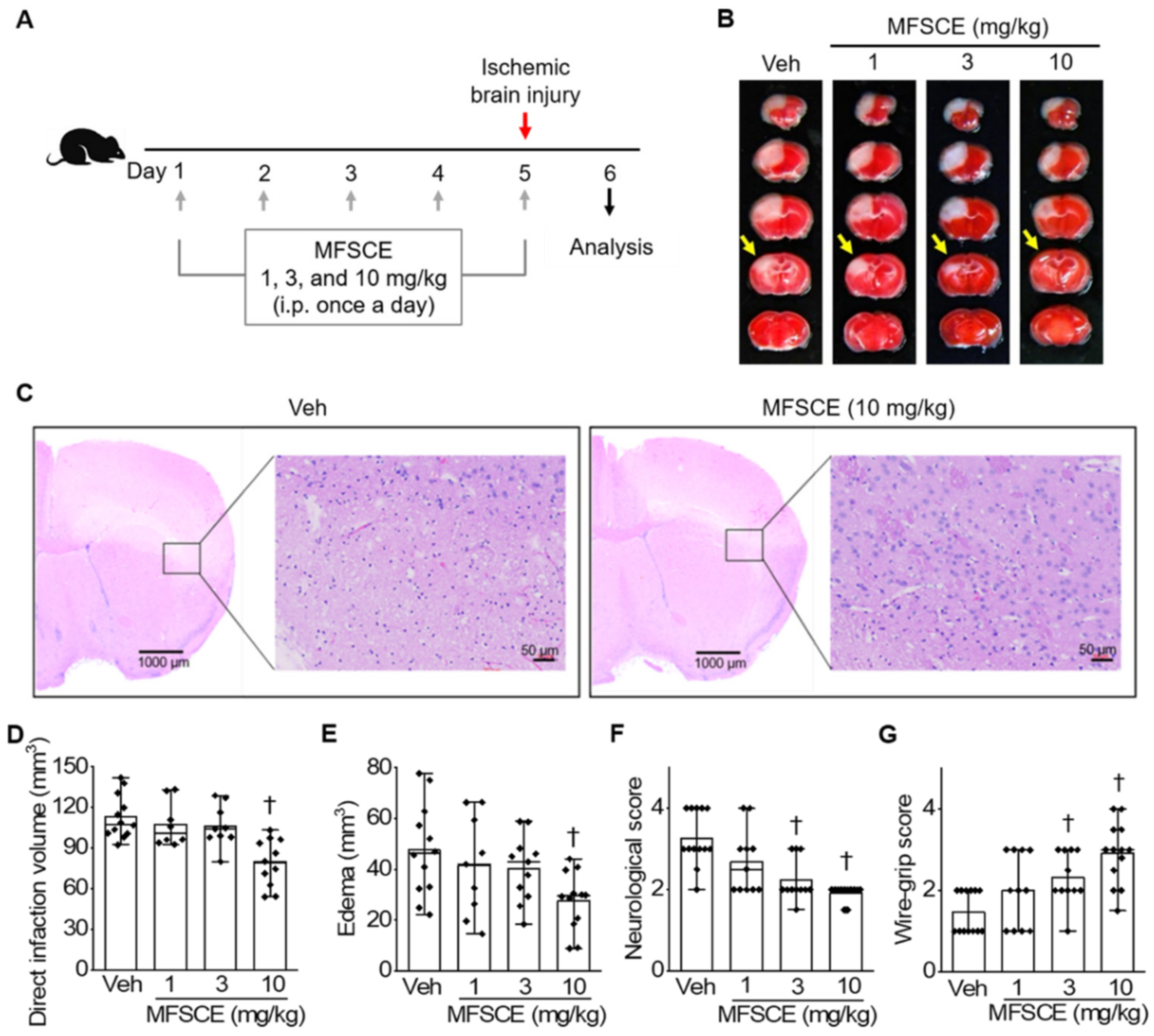
3.1. Pretreatment with MFSCE Improved Stroke Outcomes in Mice with Ischemic Stroke
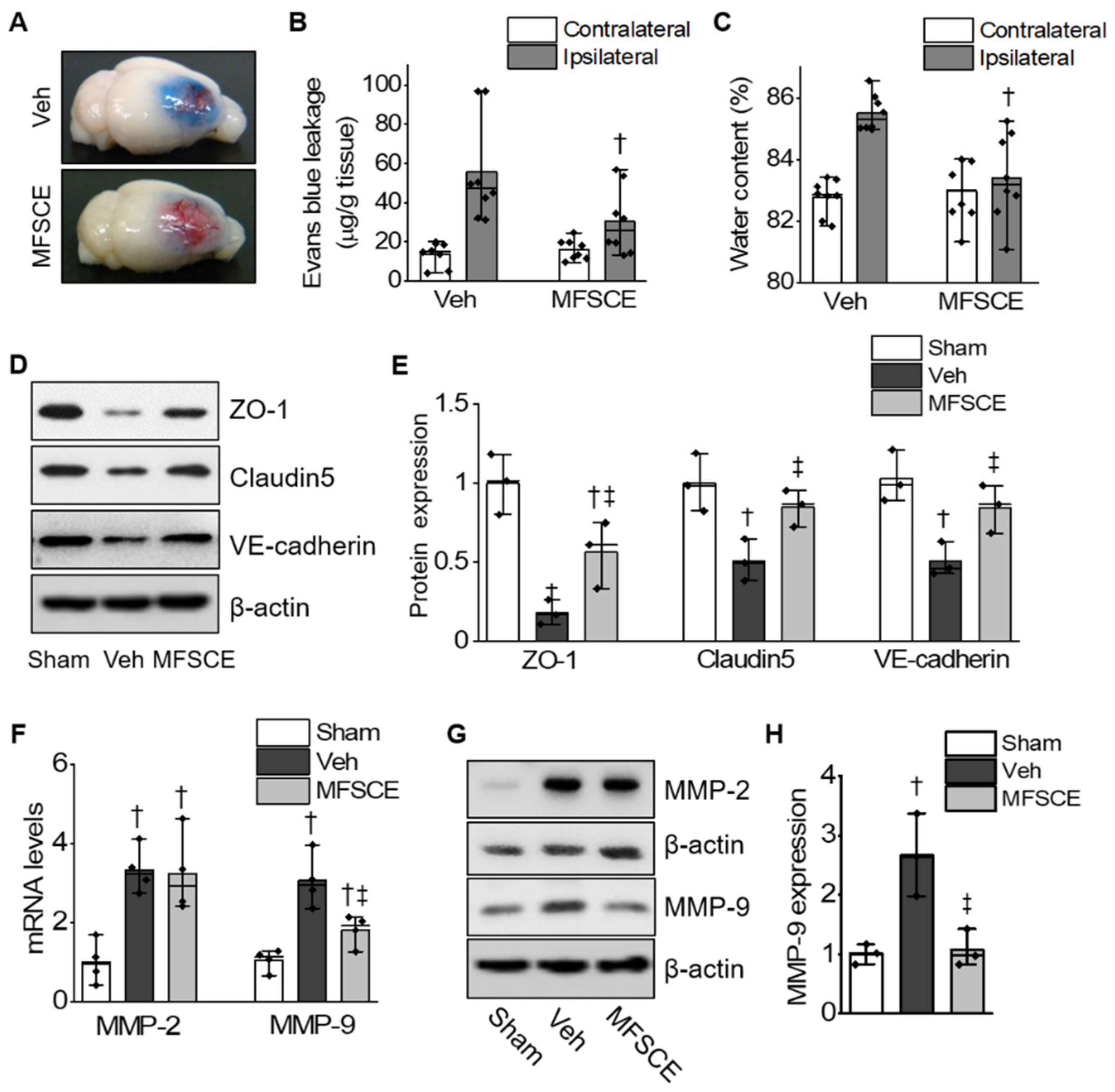
3.2. MFSCE Prevented BBB Destruction and Brain Edema by Ischemia
3.3. MFSCE Prevented Ischemic Stroke-Induced Neuronal Cell Death and Activation of the NLRP3 Inflammasome
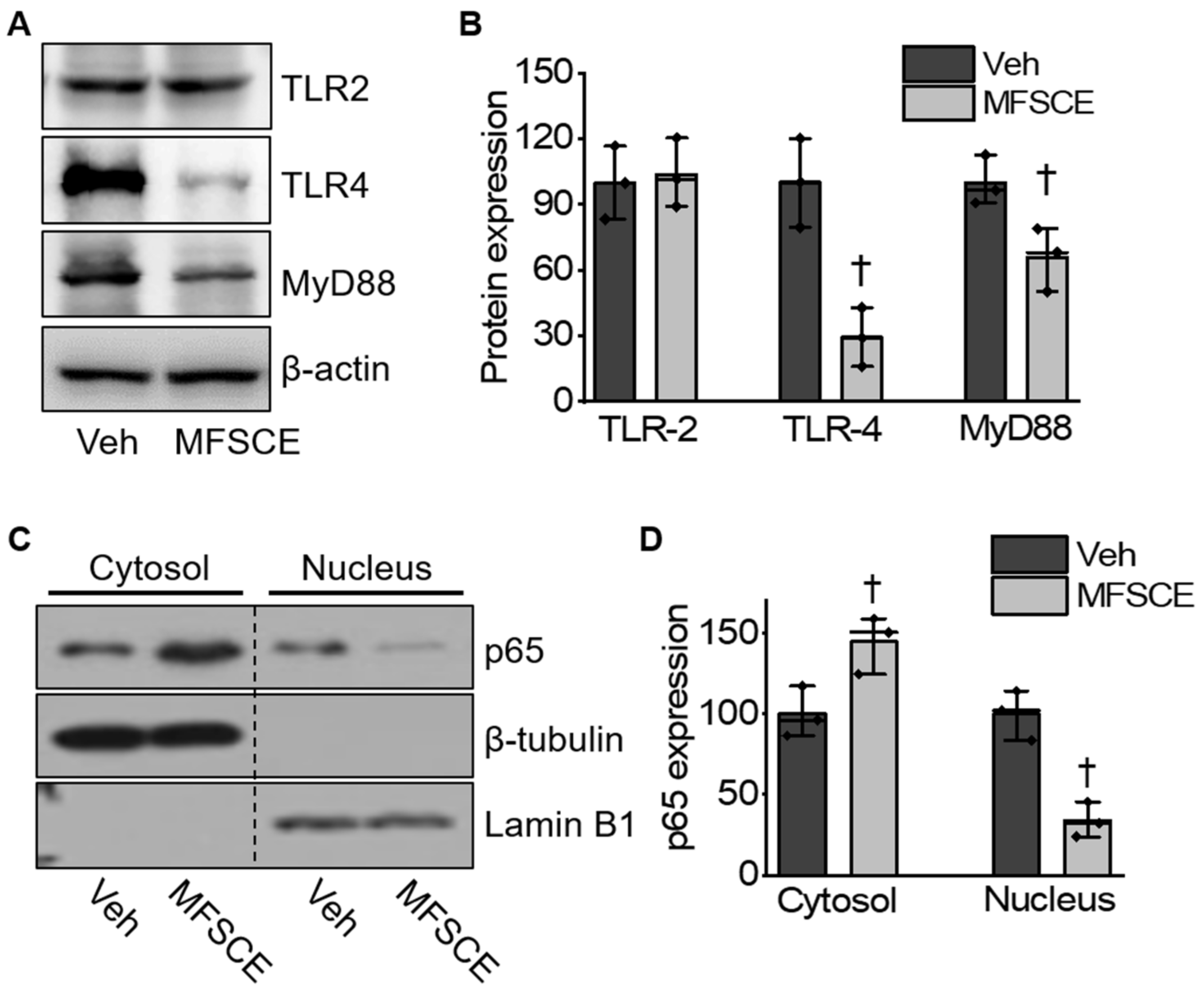
3.4. MFSCE Attenuated TLR-4/NF-κB-Mediated Inflammatory Response in Mice with Ischemic Stroke
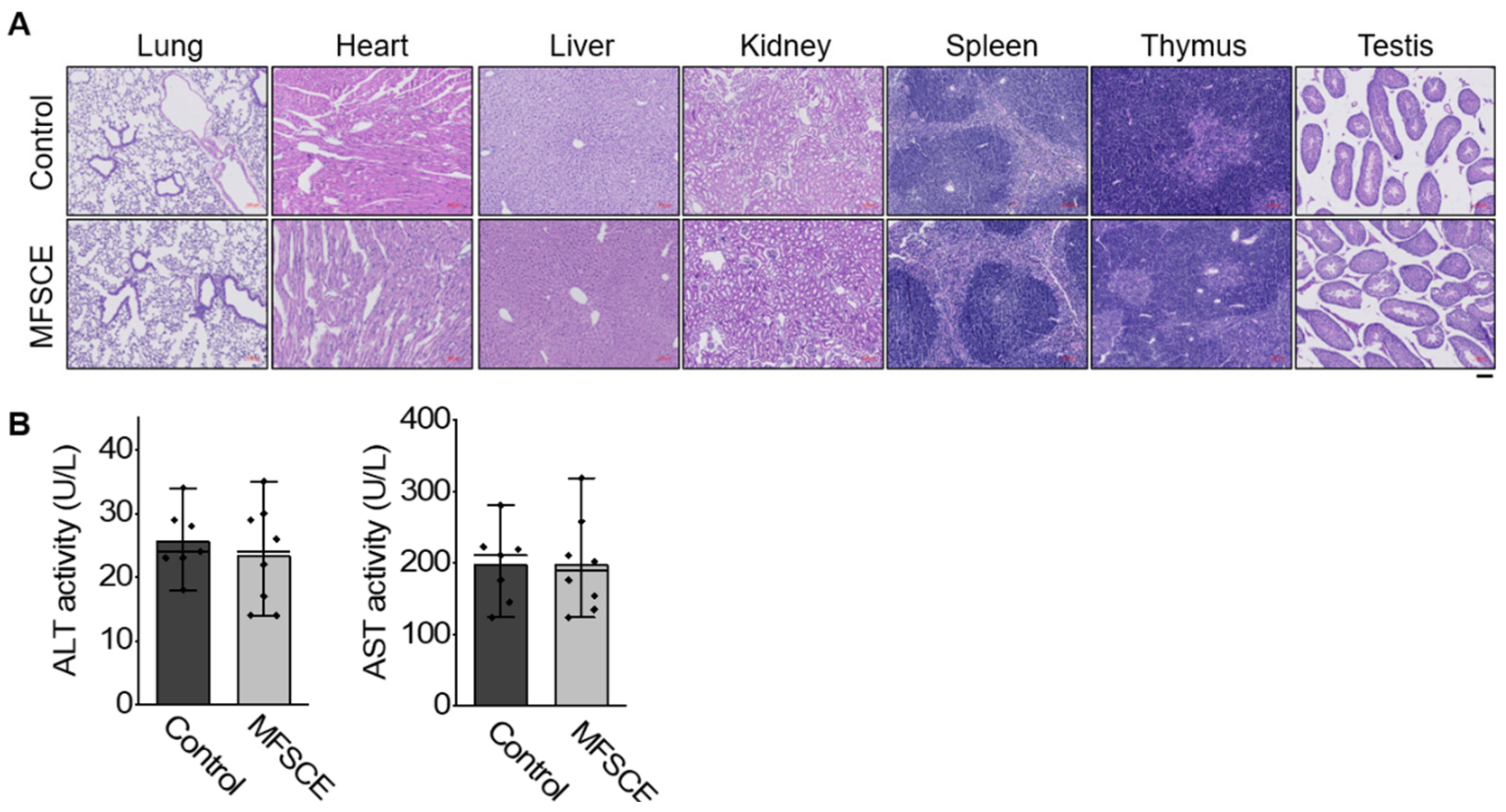
3.5. Preliminary Safety Test
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Feigin, V.L.; Norrving, B.; Mensah, G.A. Global burden of stroke. Circ. Res. 2017, 120, 439–448. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Levine, S.R.; Winn, H.R. Targeting ischemic penumbra: Part I-from pathophysiology to therapeutic strategy. J. Exp. Stroke Transl. Med. 2010, 3, 47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiang, H.; Zhang, Q.; Han, Y.; Yang, L.; Zhang, Y.; Liu, Q.; Zhang, Z.; Zhang, L. Novel brain-targeting 3-n-butylphthalide prodrugs for ischemic stroke treatment. J. Control. Release 2021, 335, 498–514. [Google Scholar] [CrossRef] [PubMed]
- Schwamm, L.H. Major advances across the spectrum of stroke care. Nat. Rev. Neurol. 2012, 8, 63–64. [Google Scholar] [CrossRef]
- Manning, N.W.; Campbell, B.C.; Oxley, T.J.; Chapot, R. Acute ischemic stroke: Time, penumbra, and reperfusion. Stroke 2014, 45, 640–644. [Google Scholar] [CrossRef] [Green Version]
- Pfefferkorn, T.; Rosenberg, G.A. Closure of the blood-brain barrier by matrix metalloproteinase inhibition reduces rtPA-mediated mortality in cerebral ischemia with delayed reperfusion. Stroke 2003, 34, 2025–2030. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Lee, S.-R.; Arai, K.; Lee, S.-R.; Tsuji, K.; Rebeck, G.W.; Lo, E.H. Lipoprotein receptor–mediated induction of matrix metalloproteinase by tissue plasminogen activator. Nat. Med. 2003, 9, 1313–1317. [Google Scholar] [CrossRef]
- Lapchak, P.A.; Chapman, D.F.; Zivin, J.A. Metalloproteinase inhibition reduces thrombolytic (tissue plasminogen activator)–induced hemorrhage after thromboembolic stroke. Stroke 2000, 31, 3034–3040. [Google Scholar] [CrossRef] [Green Version]
- Mazumder, M.K.; Bhattacharya, P.; Borah, A. Inhibition of matrix metalloproteinase-2 and 9 by Piroxicam confer neuroprotection in cerebral ischemia: An in silico evaluation of the hypothesis. Med. Hypotheses 2014, 83, 697–701. [Google Scholar] [CrossRef]
- McColl, B.W.; Rothwell, N.J.; Allan, S.M. Systemic inflammation alters the kinetics of cerebrovascular tight junction disruption after experimental stroke in mice. J. Neurosci. 2008, 28, 9451–9462. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.; Hawkins, K.E.; Doré, S.; Candelario-Jalil, E. Neuroinflammatory mechanisms of blood-brain barrier damage in ischemic stroke. Am. J. Physiol. -Cell Physiol. 2019, 316, C135–C153. [Google Scholar] [CrossRef] [PubMed]
- Martinon, F.; Burns, K.; Tschopp, J. The inflammasome: A molecular platform triggering activation of inflammatory caspases and processing of proIL-β. Mol. Cell 2002, 10, 417–426. [Google Scholar] [CrossRef]
- Schroder, K.; Tschopp, J. The inflammasomes. Cell 2010, 140, 821–832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gross, O.; Thomas, C.J.; Guarda, G.; Tschopp, J. The inflammasome: An integrated view. Immunol. Rev. 2011, 243, 136–151. [Google Scholar] [CrossRef]
- Tsuji, W.; Rubin, J.P.; Marra, K.G. Adipose-derived stem cells: Implications in tissue regeneration. World J. Stem Cells 2014, 6, 312. [Google Scholar] [CrossRef]
- Gutiérrez-Fernández, M.; Rodríguez-Frutos, B.; Ramos-Cejudo, J.; Vallejo-Cremades, M.T.; Fuentes, B.; Cerdán, S.; Díez-Tejedor, E. Effects of intravenous administration of allogenic bone marrow-and adipose tissue-derived mesenchymal stem cells on functional recovery and brain repair markers in experimental ischemic stroke. Stem Cell. Res. Ther. 2013, 4, 11. [Google Scholar] [CrossRef] [Green Version]
- Chi, L.; Huang, Y.; Mao, Y.; Wu, K.; Zhang, L.; Nan, G. Tail vein infusion of adipose-derived mesenchymal stem cell alleviated inflammatory response and improved blood brain barrier condition by suppressing endoplasmic reticulum stress in a middle cerebral artery occlusion rat model. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2018, 24, 3946. [Google Scholar] [CrossRef]
- Venkatarame Gowda Saralamma, V.; Vetrivel, P.; Kim, S.M.; Ha, S.E.; Lee, H.J.; Lee, S.J.; Kim, Y.S.; Pak, J.E.; Lee, H.J.; Heo, J.D. Proteome Profiling of Membrane-Free Stem Cell Components by Nano-LS/MS Analysis and Its Anti-Inflammatory Activity. Evid. Based Complement. Alternat. Med. 2019, 2019, 4683272. [Google Scholar] [CrossRef]
- Lee, J.-K.; Park, M.-S.; Kim, Y.-S.; Moon, K.-S.; Joo, S.-P.; Kim, T.-S.; Kim, J.-H.; Kim, S.-H. Photochemically induced cerebral ischemia in a mouse model. Surg. Neurol. 2007, 67, 620–625. [Google Scholar] [CrossRef]
- Lin, T.-N.; He, Y.Y.; Wu, G.; Khan, M.; Hsu, C.Y. Effect of brain edema on infarct volume in a focal cerebral ischemia model in rats. Stroke 1993, 24, 117–121. [Google Scholar] [CrossRef] [Green Version]
- Hattori, K.; Lee, H.; Hurn, P.D.; Crain, B.J.; Traystman, R.J.; DeVries, A.C. Cognitive deficits after focal cerebral ischemia in mice. Stroke 2000, 31, 1939–1944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uyama, O.; Okamura, N.; Yanase, M.; Narita, M.; Kawabata, K.; Sugita, M. Quantitative evaluation of vascular permeability in the gerbil brain after transient ischemia using Evans blue fluorescence. J. Cereb. Blood Flow Metab. 1988, 8, 282–284. [Google Scholar] [CrossRef] [Green Version]
- Kamoun, W.S.; Ley, C.D.; Farrar, C.T.; Duyverman, A.M.; Lahdenranta, J.; Lacorre, D.A.; Batchelor, T.T.; di Tomaso, E.; Duda, D.G.; Munn, L.L. Edema control by cediranib, a vascular endothelial growth factor receptor–targeted kinase inhibitor, prolongs survival despite persistent brain tumor growth in mice. J. Clin. Oncol. 2009, 27, 2542. [Google Scholar] [CrossRef] [PubMed]
- Hao, L.; Zou, Z.; Tian, H.; Zhang, Y.; Zhou, H.; Liu, L. Stem cell-based therapies for ischemic stroke. BioMed Res. Int. 2014, 2014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leu, S.; Lin, Y.-C.; Yuen, C.-M.; Yen, C.-H.; Kao, Y.-H.; Sun, C.-K.; Yip, H.-K. Adipose-derived mesenchymal stem cells markedly attenuate brain infarct size and improve neurological function in rats. J. Transl. Med. 2010, 8, 63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, X.; Du, Z.; Zhao, L.; Feng, D.; Wei, G.; He, Y.; Tan, J.; Lee, W.H.; Hampel, H.; Dodel, R. IFATS collection: The conditioned media of adipose stromal cells protect against hypoxia-ischemia-induced brain damage in neonatal rats. Stem Cells 2009, 27, 478–488. [Google Scholar] [CrossRef] [PubMed]
- Kalbermatten, D.F.; Schaakxs, D.; Kingham, P.J.; Wiberg, M. Neurotrophic activity of human adipose stem cells isolated from deep and superficial layers of abdominal fat. Cell Tissue Res. 2011, 344, 251–260. [Google Scholar] [CrossRef]
- Rehman, J.; Traktuev, D.; Li, J.; Merfeld-Clauss, S.; Temm-Grove, C.J.; Bovenkerk, J.E.; Pell, C.L.; Johnstone, B.H.; Considine, R.V.; March, K.L. Secretion of angiogenic and antiapoptotic factors by human adipose stromal cells. Circulation 2004, 109, 1292–1298. [Google Scholar] [CrossRef]
- Zhang, Y.; Deng, H.; Hu, Y.; Pan, C.; Wu, G.; Li, Q.; Tang, Z. Adipose-derived mesenchymal stem cells stereotactic transplantation alleviate brain edema from intracerebral hemorrhage. J. Cell. Biochem. 2019, 120, 14372–14382. [Google Scholar] [CrossRef]
- Lv, H.; Li, J.; Che, Y. miR-31 from adipose stem cell-derived extracellular vesicles promotes recovery of neurological function after ischemic stroke by inhibiting TRAF6 and IRF5. Exp. Neurol. 2021, 113611. [Google Scholar] [CrossRef]
- Shi, Y.; Zhang, L.; Pu, H.; Mao, L.; Hu, X.; Jiang, X.; Xu, N.; Stetler, R.A.; Zhang, F.; Liu, X. Rapid endothelial cytoskeletal reorganization enables early blood–brain barrier disruption and long-term ischaemic reperfusion brain injury. Nat. Commun. 2016, 7, 10523. [Google Scholar] [CrossRef] [PubMed]
- Lo, E.H.; Singhal, A.B.; Torchilin, V.P.; Abbott, N.J. Drug delivery to damaged brain. Brain Res. Rev. 2001, 38, 140–148. [Google Scholar] [CrossRef]
- Dejonckheere, E.; Vandenbroucke, R.E.; Libert, C. Matrix metalloproteinases as drug targets in ischemia/reperfusion injury. Drug Discov. Today 2011, 16, 762–778. [Google Scholar] [CrossRef] [PubMed]
- Rosell, A.; Cuadrado, E.; Ortega-Aznar, A.; Hernández-Guillamon, M.; Lo, E.H.; Montaner, J. MMP-9–positive neutrophil infiltration is associated to blood–brain barrier breakdown and basal lamina type iv collagen degradation during hemorrhagic transformation after human ischemic stroke. Stroke 2008, 39, 1121–1126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, G.; Liu, Y.; Zhang, Z.; Lu, Y.; Wang, Y.; Huang, J.; Li, Y.; Chen, X.; Gu, X.; Wang, Y. Mesenchymal stem cells maintain blood-brain barrier integrity by inhibiting aquaporin-4 upregulation after cerebral ischemia. Stem Cells 2014, 32, 3150–3162. [Google Scholar] [CrossRef] [PubMed]
- Obermeier, B.; Daneman, R.; Ransohoff, R.M. Development, maintenance and disruption of the blood-brain barrier. Nat. Med. 2013, 19, 1584–1596. [Google Scholar] [CrossRef]
- Jack, C.S.; Arbour, N.; Manusow, J.; Montgrain, V.; Blain, M.; McCrea, E.; Shapiro, A.; Antel, J.P. TLR signaling tailors innate immune responses in human microglia and astrocytes. J. Immunol. 2005, 175, 4320–4330. [Google Scholar] [CrossRef] [Green Version]
- Bsibsi, M.; Ravid, R.; Gveric, D.; van Noort, J.M. Broad expression of Toll-like receptors in the human central nervous system. J. Neuropathol. Exp. Neurol. 2002, 61, 1013–1021. [Google Scholar] [CrossRef] [Green Version]
- Olson, J.K.; Miller, S.D. Microglia initiate central nervous system innate and adaptive immune responses through multiple TLRs. J. Immunol. 2004, 173, 3916–3924. [Google Scholar] [CrossRef] [Green Version]
- Bowman, C.C.; Rasley, A.; Tranguch, S.L.; Marriott, I. Cultured astrocytes express toll-like receptors for bacterial products. Glia 2003, 43, 281–291. [Google Scholar] [CrossRef]
- Hyakkoku, K.; Hamanaka, J.; Tsuruma, K.; Shimazawa, M.; Tanaka, H.; Uematsu, S.; Akira, S.; Inagaki, N.; Nagai, H.; Hara, H. Toll-like receptor 4 (TLR4), but not TLR3 or TLR9, knock-out mice have neuroprotective effects against focal cerebral ischemia. Neuroscience 2010, 171, 258–267. [Google Scholar] [CrossRef] [PubMed]
- Tang, S.-C.; Arumugam, T.V.; Xu, X.; Cheng, A.; Mughal, M.R.; Jo, D.G.; Lathia, J.D.; Siler, D.A.; Chigurupati, S.; Ouyang, X. Pivotal role for neuronal Toll-like receptors in ischemic brain injury and functional deficits. Proc. Natl. Acad. Sci. USA 2007, 104, 13798–13803. [Google Scholar] [CrossRef] [Green Version]
- Seki, E.; Tsutsui, H.; Iimuro, Y.; Naka, T.; Son, G.; Akira, S.; Kishimoto, T.; Nakanishi, K.; Fujimoto, J. Contribution of Toll-like receptor/myeloid differentiation factor 88 signaling to murine liver regeneration. Hepatology 2005, 41, 443–450. [Google Scholar] [CrossRef] [PubMed]
- González, M.A.; Gonzalez–Rey, E.; Rico, L.; Büscher, D.; Delgado, M. Adipose-derived mesenchymal stem cells alleviate experimental colitis by inhibiting inflammatory and autoimmune responses. Gastroenterology 2009, 136, 978–989. [Google Scholar] [CrossRef] [PubMed]
- Yan, K.; Bian, J.-R.; He, L.; Song, B.-W.; Dong, L.; He, J.-W.; Shen, L.-H.; Zhou, X.-Z.; Zhen, Y. Adipose-derived Mesenchymal Stem Cells Protect Neurons by Inhibiting Microglial Inflammation. 2021. [Google Scholar] [CrossRef]
- Feng, N.; Jia, Y.; Huang, X. Exosomes from adipose-derived stem cells alleviate neural injury caused by microglia activation via suppressing NF-kB and MAPK pathway. J. Neuroimmunol. 2019, 334, 576996. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ryu, J.H.; Kim, Y.; Kim, M.J.; Park, J.; Kim, J.W.; Park, H.S.; Kim, Y.S.; Shin, H.K.; Shin, Y.-I. Membrane-Free Stem Cell Extract Enhances Blood–Brain Barrier Integrity by Suppressing NF-κB-Mediated Activation of NLRP3 Inflammasome in Mice with Ischemic Stroke. Life 2022, 12, 503. https://doi.org/10.3390/life12040503
Ryu JH, Kim Y, Kim MJ, Park J, Kim JW, Park HS, Kim YS, Shin HK, Shin Y-I. Membrane-Free Stem Cell Extract Enhances Blood–Brain Barrier Integrity by Suppressing NF-κB-Mediated Activation of NLRP3 Inflammasome in Mice with Ischemic Stroke. Life. 2022; 12(4):503. https://doi.org/10.3390/life12040503
Chicago/Turabian StyleRyu, Ji Hyeon, Yeonye Kim, Min Jae Kim, Jisu Park, Ji Won Kim, Hye Sook Park, Young Sil Kim, Hwa Kyoung Shin, and Yong-Il Shin. 2022. "Membrane-Free Stem Cell Extract Enhances Blood–Brain Barrier Integrity by Suppressing NF-κB-Mediated Activation of NLRP3 Inflammasome in Mice with Ischemic Stroke" Life 12, no. 4: 503. https://doi.org/10.3390/life12040503
APA StyleRyu, J. H., Kim, Y., Kim, M. J., Park, J., Kim, J. W., Park, H. S., Kim, Y. S., Shin, H. K., & Shin, Y.-I. (2022). Membrane-Free Stem Cell Extract Enhances Blood–Brain Barrier Integrity by Suppressing NF-κB-Mediated Activation of NLRP3 Inflammasome in Mice with Ischemic Stroke. Life, 12(4), 503. https://doi.org/10.3390/life12040503