
Life on Minerals: Binding Behaviors of Oligonucleotides on Zirconium Silicate and Its Inhibitory Activity for the Self-Cleavage of Hammerhead Ribozyme

 ,
, 
Abstract

1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Nucleic Acid Solutions Preparation
2.3. Self-Cleavage Behaviors of ASBVd(−):HHR on Minerals
2.4. HPLC Analysis for the Products of ASBVd(−):HHR Self-Cleavage
2.5. Secondary Structure Modeling of RNA and DNA
3. Results and Discussion
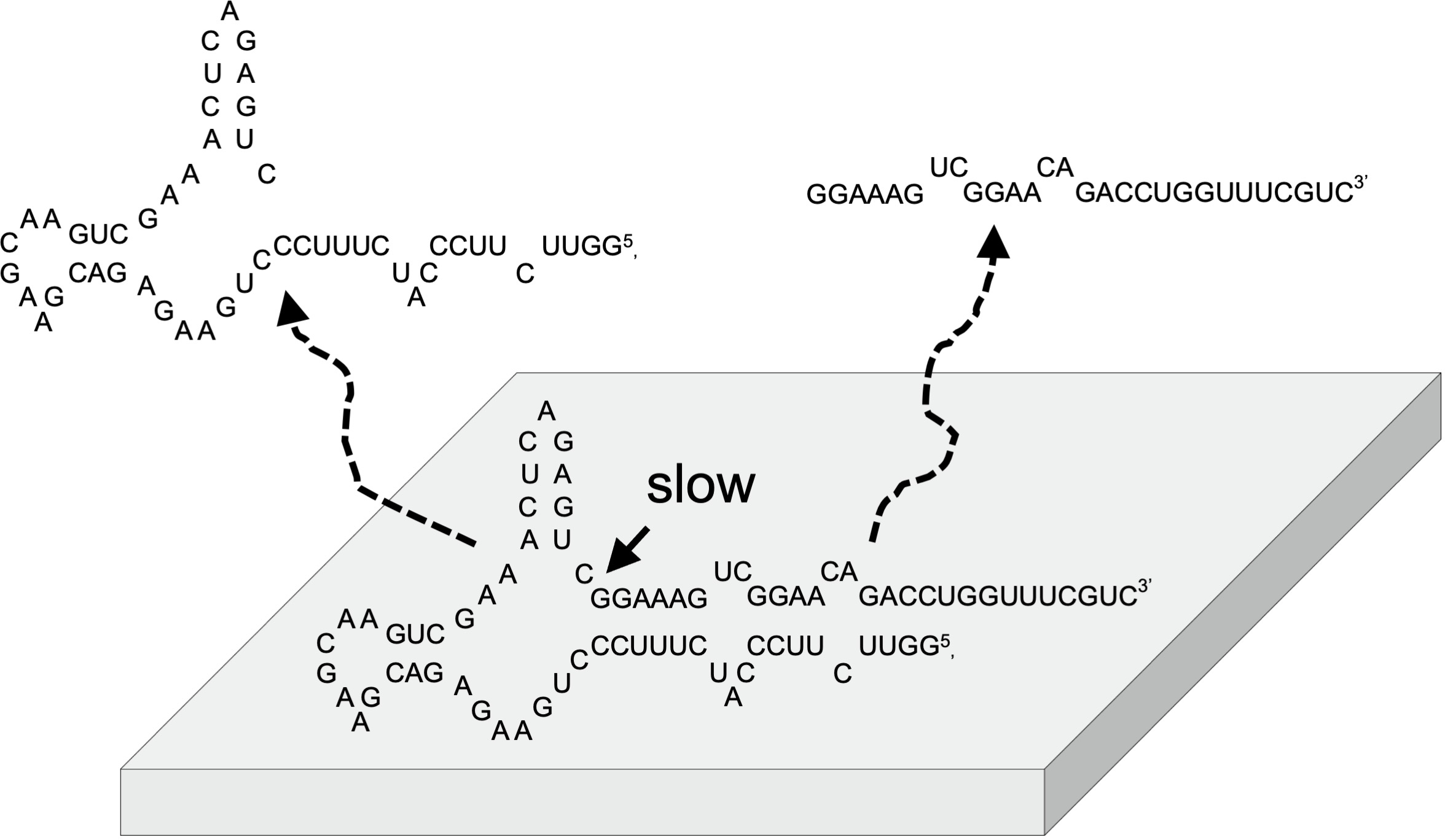
3.1. Concept of RNA Chemical Network Model on Mineral
3.2. Selection of Oligonucleotides and Minerals
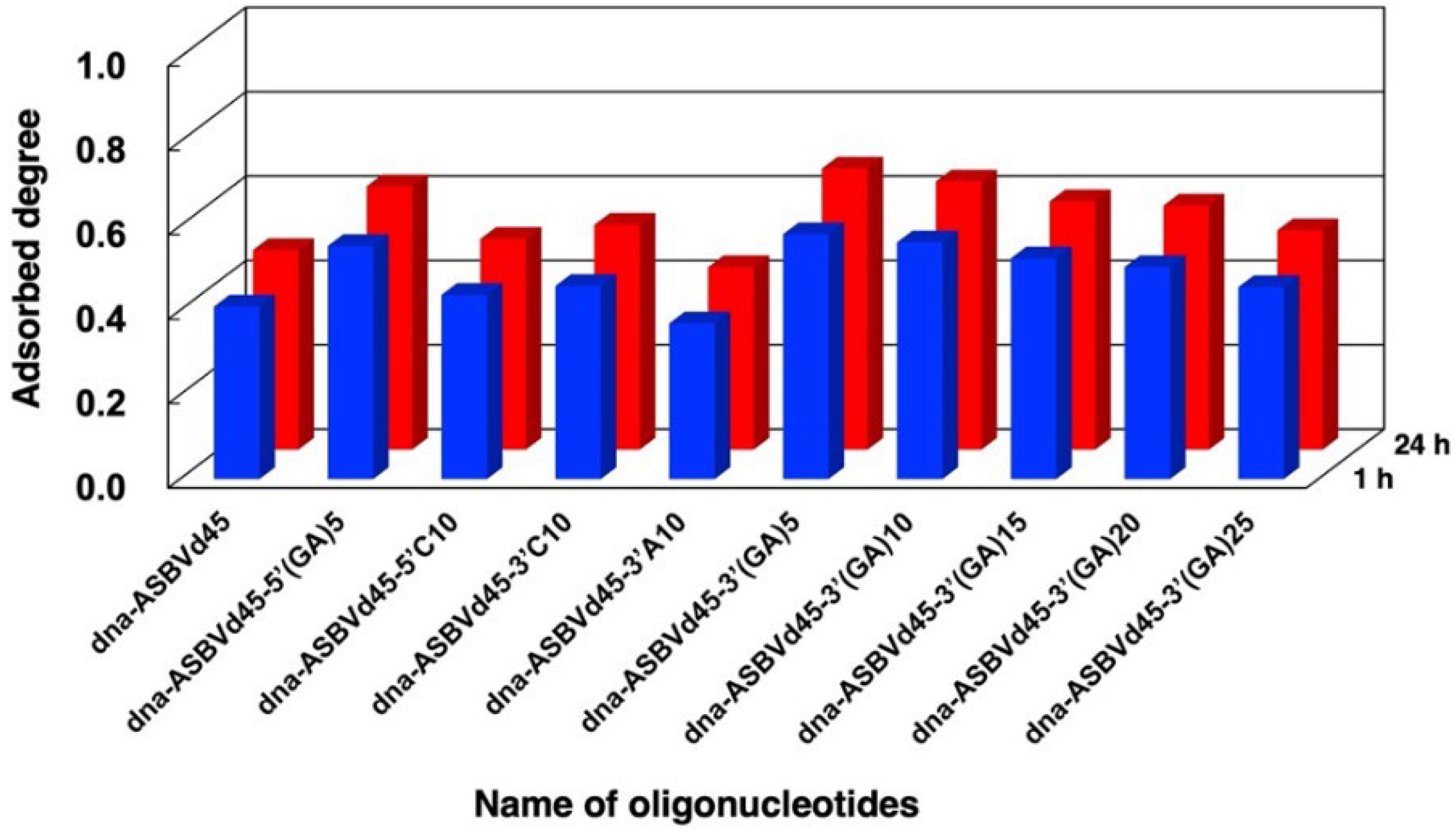
3.3. Binding Behaviors of DNA
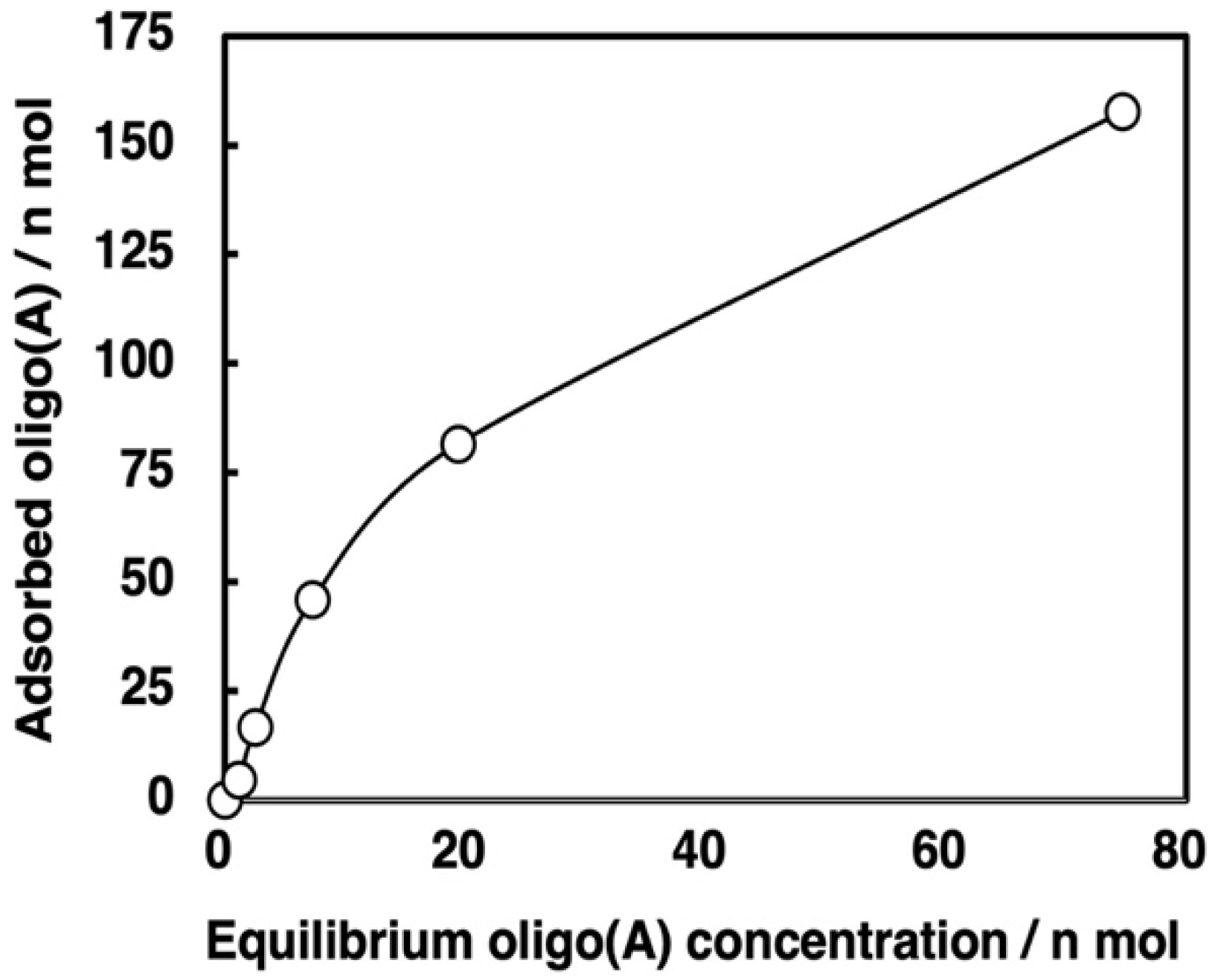
3.4. Binding Behaviors of Oligo(A)
3.5. Binding Behaviors of Ribozymes and Desorption by EDTA Solution
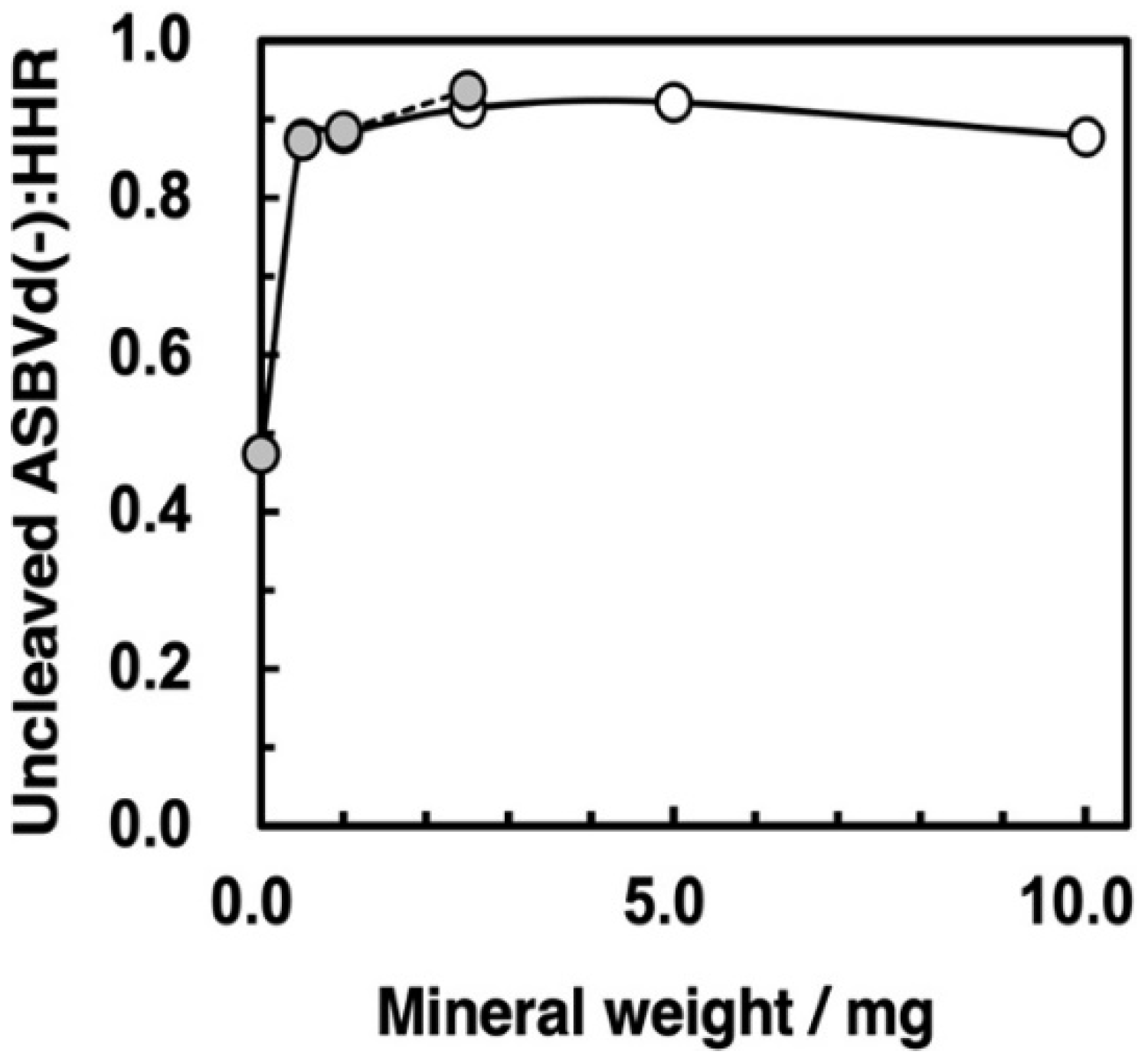
3.6. Self-Cleavage of ASBVd(−):HHR on Zirconium Silicate and Montmorillonite Surfaces
3.7. Role of Minerals for the Chemical Evolution of Ribozymes on Hadean Earth
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gilbert, W. Origins of life—The RNA world. Nature 1986, 319, 618. [Google Scholar] [CrossRef]
- Robertson, M.P.; Joyce, G.F. The origins of the RNA world. Cold Spring Harb. Perspect. Biol. 2012, 4, a003608. [Google Scholar] [CrossRef] [PubMed]
- Orgel, L.E. Prebiotic chemistry and the origin of the RNA world. Cri. Rev. Biochem. Mol. Biol. 2004, 39, 99–123. [Google Scholar] [CrossRef]
- Cafferty, B.J.; Hud, N.V. Abiotic synthesis of RNA in water: A common goal of prebiotic chemistry and bottom-up synthetic biology. Curr. Opin. Chem. Biol. 2014, 22, 146–157. [Google Scholar] [CrossRef] [PubMed]
- Ellington, A.D.; Szostak, J.W. In vitro selection of RNA molecules that bind specific ligands. Nature 1990, 346, 818–822. [Google Scholar] [CrossRef]
- Tjhunga, K.F.; Shokhireva, M.N.; Horninga, D.P.; Joyce, G.F. An RNA polymerase ribozyme that synthesizes its own ancestor. Proc. Natl. Acad. Sci. USA 2020, 116, 2906–2913. [Google Scholar] [CrossRef]
- Kawamura, K. Hydrothermal microflow technology as a research tool for origin-of-life studies in extreme earth environments. Life 2017, 7, 37. [Google Scholar] [CrossRef]
- Maurel, M.-C.; Leclerc, F.; Hervé, G. Ribozyme chemistry: To be or not to be under high pressure. Chem. Rev. 2020, 120, 4898–4918. [Google Scholar] [CrossRef]
- Kaddour, H.; Lucchi, H.; Hervé, G.; Vergne, J.; Maurel, M.-C. Kinetic study of the Avocado Sunblotch Viroid self-cleavage reaction reveals compensatory effects between high-pressure and high-temperature: Implications for origins of life on Earth. Biology 2021, 10, 720. [Google Scholar] [CrossRef]
- Kawamura, K.; Ogawa, M.; Konagaya, N.; Maruoka, Y.; Lambert, J.-F.; Ter-Ovanessian, L.M.P.; Vergne, J.; Hervé, G.; Maurel, M.-C. A high-pressure—Temperature flow reactor simulating the Hadean Earth environments applicable to pressure dependence of the cleavage of Avocado viroid hammerhead ribozyme. Life 2022, 12, 1224. [Google Scholar] [CrossRef]
- Ferris, J.P.; Ertem, G. Oligomerization of ribonucleotides on montmorillonite: Reaction of the 5′-phosphorimidazolide of adenosine. Science 1992, 257, 1387–1389. [Google Scholar] [CrossRef] [PubMed]
- Ferris, J.P. Mineral catalysis and prebiotic synthesis: Montmorillonite-catalyzed formation of RNA. Elements 2005, 1, 145–149. [Google Scholar] [CrossRef]
- Franchi, M.; Gallori, E. A surface-mediated origin of the RNA world: Biogenic activities of clay-adsorbed RNA molecules. Gene 2005, 346, 205–214. [Google Scholar] [CrossRef] [PubMed]
- Biondi, E.; Branciamore, S.S.; Maurel, M.-C.; Gallori, E. Montmorillonite protection of an UV-irradiated hairpin ribozyme: Evolution of the RNA world in a mineral environment. BMC Evol. Biol. 2007, 7 (Suppl. 2), S2. [Google Scholar] [CrossRef]
- Biondi, E.; Branciamore, S.; Fusi, L.; Gago, S.; Gallori, E. Catalytic activity of hammerhead ribozymes in a clay mineral environment: Implications for the RNA world. Gene 2007, 389, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Saladino, R.; Crestini, C.; Ciciriello, F.; Pino, S.; Costanzo, G.; Di Mauro, E. From formamide to RNA: The roles of formamide and water in the evolution of chemical information. Res. Microbiol. 2009, 160, 441–448. [Google Scholar] [CrossRef] [PubMed]
- Saladino, R.; Crestini, C.; Cossetti, C.; Di Mauro, E.; Deamer, D. Catalytic effects of Murchison material: Prebiotic synthesis and degradation of RNA precursors. Orig. Life Evol. Biosph. 2011, 41, 437–451. [Google Scholar] [CrossRef]
- Stephenson, J.D.; Popović, M.; Bristow, T.F.; Ditzler, M.A. Evolution of ribozymes in the presence of a mineral surface. RNA 2016, 22, 1893–1901. [Google Scholar] [CrossRef]
- Biondi, E.; Furukawa, Y.; Kawai, J.; Benner, S.A. Adsorption of RNA on mineral surfaces and mineral precipitates. Beilstein J. Org. Chem. 2017, 13, 393–404. [Google Scholar] [CrossRef]
- Ter-Ovanessian, L.M.P.; Rigaud, B.; Mezzetti, A.; Lambert, J.-F.; Maurel, M.-C. Carbamoyl phosphate and its substitutes for the uracil synthesis in origins of life scenarios. Sci. Rep. 2021, 11, 19356. [Google Scholar] [CrossRef]
- Rahman, M.M.; Matsumura, S.; Ikawa, Y. Oligomerization of a Bimolecular ribozyme modestly rescues its structural defects that disturb interdomain assembly to form the catalytic site. J. Mol. Evol. 2018, 86, 431–442. [Google Scholar] [CrossRef]
- Mizuuchi, R.; Blokhuis, A.; Vincent, L.; Nghe, P.; Lehman, N.; Baumd, D. Mineral surfaces select for longer RNA molecules. Chem. Comm. 2019, 55, 2090–2093. [Google Scholar] [CrossRef] [PubMed]
- Cairns-Smith, A.G. Genetic Takeover and the Mineral Origins of Life; Cambridge University Press: Cambridge, UK, 1982. [Google Scholar]
- Müller, U.F. Re-creating an RNA world. Cell Mol. Life Sci. 2006, 63, 1278–1293. [Google Scholar] [CrossRef]
- Yarus, M. Getting past the RNA world: The initial Darwinian ancestor. Cold Spring Harb. Perspect. Biol. 2011, 3, a003590. [Google Scholar] [CrossRef]
- El-Murr, N.; Maurel, M.-C.; Rihova, M.; Vergne, J.; Hervé, G.; Kato, M.; Kawamura, K. Behavior of a hammerhead ribozyme in aqueous solution at medium to high temperatures. Naturwissenschaften 2012, 99, 731–738. [Google Scholar] [CrossRef] [PubMed]
- Reuter, J.S.; Mathews, D.H. RNA structure: Software for RNA secondary structure prediction and analysis. BMC Bioinform. 2010, 11, 129. [Google Scholar] [CrossRef] [PubMed]
- Available online: https://rna.urmc.rochester.edu (accessed on 23 November 2021).
- Kawamura, K. Reality of the emergence of life-like systems from simple prebiotic polymers on primitive earth. In Genesis—In the Beginning: Precursors of Life, Chemical Models and Early Biological Evolution; Seckbach, J., Gordon, R., Eds.; Springer: Dordrecht, The Netherlands, 2012; pp. 123–144. [Google Scholar] [CrossRef]
- Kawamura, K. A hypothesis: Life initiated from two genes, as deduced from the RNA world hypothesis and the characteristics of life-like systems. Life 2016, 6, 29. [Google Scholar] [CrossRef]
- Hansma, H.G. Possible origin of life between mica sheets. J. Theor. Biol. 2010, 266, 175–188. [Google Scholar] [CrossRef]
- Kawamura, K.; Ferris, J.P. Kinetics and mechanistic analysis of dinucleotide and oligonucleotide formation from the 5’-phosphorimidazolide of adenosine on Na+-montmorillonite. JACS 1994, 116, 7564–7572. [Google Scholar] [CrossRef]
- Kawamura, K.; Ferris, J.P. Clay catalysis of oligonucleotide formation: Kinetics of the reaction of the 5′-phosphorimidazolides of nucleotides with the non-basic heterocycles uracil and hypoxanthine. Orig. Life Evol. Biosph. 1999, 29, 563–591. [Google Scholar] [CrossRef]
- de la Peña, M.; Gago, S.; Flores, R. Peripheral regions of natural hammerhead ribozymes greatly increase their self-cleavage activity. EMBO J. 2003, 22, 5561–5570. [Google Scholar] [CrossRef] [PubMed]
- Kaddour, H.; Vergne, J.; Guy Hervé, G.; Maurel, M.-C. High-pressure analysis of a hammerhead ribozyme from Chrysanthemum chlorotic mottle viroid reveals two different populations of self-cleaving molecule. FEBS J. 2011, 278, 3739–3747. [Google Scholar] [CrossRef] [PubMed]
- Turova, V.V.; Lebodab, R. Application of 1H NMR spectroscopy method for determination of characteristics of thin layers of water adsorbed on the surface of dispersed and porous adsorbents. Adv. Coll. Int. Sci. 1999, 79, 173–211. [Google Scholar] [CrossRef]
- Sulpizi, M.; Gaigeot, M.P.; Sprik, M. The silica−water interface: How the silanols determine the surface acidity and modulate the water properties. J. Chem. Theory Comput. 2012, 8, 1037–1047. [Google Scholar] [CrossRef]
- Mojzsis, S.; Harrison, T.; Pidgeon, R. Oxygen-isotope evidence from ancient zircons for liquid water at the Earth’s surface 4300 Myr ago. Nature 2001, 409, 178–181. [Google Scholar] [CrossRef]
- Wilde, S.; Valley, J.; Peck, W.; Graham, C.M. Evidence from detrital zircons for the existence of continental crust and oceans on the Earth 4.4 Gyr ago. Nature 2001, 409, 175–178. [Google Scholar] [CrossRef]
- Montgomery, W.; Tuff, J.; Kohn, S.C.; Jones, R.L. Reactions between organic acids and montmorillonite clay under Earth-forming conditions. Chem. Geol. 2011, 283, 171–176. [Google Scholar] [CrossRef]
- Mahmoud, M.E.; Nabil, G.M.; Mahmoud, S.M.E. High performance nano-zirconium silicate adsorbent for efficient removal of copper (II), cadmium (II) and lead (II). J. Env. Chem. Eng. 2016, 3, 1320–1328. [Google Scholar] [CrossRef]
- de Oliveira, L.H.; Trigueiro, P.; Rigaud, B.; da Silva-Filho, E.C.; Osajima, J.A.; Fonseca, M.G.; Lambert, J.-F.; Georgelin, T.; Jaber, M. When RNA meets montmorillonite: Influence of the pH and divalent cations. Appl. Clay Sci. 2021, 214, 106234. [Google Scholar] [CrossRef]
- Mao, M.; Fornasiero, D.; Ralston, J.; Smart, R.S.C.; Sobieraj, S. Electrochemistry of the zircon—Water interface. Coll. Surf. Int. A 1994, 85, 37–49. [Google Scholar] [CrossRef]
- Mathias, J.; Wannemacher, G. Basic characteristics and applications of aerosil: 30. The chemistry and physics of the aerosil surface. J. Coll. Inter. Sci. 1988, 125, 61–68. [Google Scholar] [CrossRef]
- Kitayama, Y.; Muraoka, M.; Konno, T.; Kodama, T.; Abe, J.; Okamura, M. Substitution of Mg2+ in sepiolite with Cu2+ in aqueous solutions of various cupric salts. Clay Sci. 1996, 10, 83–93. [Google Scholar] [CrossRef]
- Lazarević, S.; Janković-Častvan, I.; Djokić, V.; Radovanović, Z.; Janaćković, D.; Petrović, R. Iron-modified sepiolite for Ni2+ sorption from aqueous solution: An equilibrium, kinetic, and thermodynamic study. J. Chem. Eng. Data 2010, 55, 5681–5689. [Google Scholar] [CrossRef]
- Eren, E.; Afsin, B. Investigation of a basic dye adsorption from aqueous solution onto raw and pre-treated sepiolite surfaces. Dye. Pigment. 2007, 73, 162–167. [Google Scholar] [CrossRef]
- Swadling, J.B.; Wright, D.W.; Suter, J.L.; Coveney, P.V. Structure, dynamics, and function of the hammerhead ribozyme in bulk water and at a clay mineral surface from replica exchange molecular dynamics. Langmuir 2015, 31, 2493–2501. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| # | RNA Name | Sequence |
|---|---|---|
| 1 | ASBVd79(−):HHR | GGUUCUUCCC AUCUUUCCCU GAAGAGACGA AGCAAGUCGA AACUCAGAGU CGGAAAGUCG GAACAGACCU GGUUUCGUC |
| 2 | rna-ASBVd45 | CUUUCCCU GAAGAGACGA AGCAAGUCGA AACUCAGAGU CGGAAAG |
| 3 | rna-ASBVd45-3′(GA)5 | CUUUCCCU GAAGAGACGA AGCAAGUCGA AACUCAGAGU CGGAAAG GAGAGAGAGA |
| 4 | rna-ASBVd45-3′C10 | CUUUCCCU GAAGAGACGA AGCAAGUCGA AACUCAGAGU CGGAAAG CCCCCCCCCC |
| 5 | rna-ASBVd45-5′(GA)5 | GAGAGAGAGA CUUUCCCU GAAGAGACGA AGCAAGUCGA AACUCAGAGU CGGAAAG |
| 6 | rna-ASBVd45-5′C10 | CCCCCCCCCC CUUUCCCU GAAGAGACGA AGCAAGUCGA AACUCAGAGU CGGAAAG |
| # | DNA Name | Sequence |
|---|---|---|
| 1 | dna-ASBVd45 | CTTTCCCT GAAGAGACGA AGCAAGTCGA AACTCAGAGT CGGAAAG |
| 2 | dna-ASBVd45-5′(GA)5 | GAGAGAGAGA CTTTCCCT GAAGAGACGA AGCAAGTCGA AACTCAGAGT CGGAAAG |
| 3 | dna-ASBVd45-5′C10 | CCCCCCCCCC CTTTCCCT GAAGAGACGA AGCAAGTCGA AACTCAGAGT CGGAAAG |
| 4 | dna-ASBVd45-3′C10 | CTTTCCCT GAAGAGACGA AGCAAGTCGA AACTCAGAGT CGGAAAG CCCCCCCCCC |
| 5 | dna-ASBVd45-3′A10 | CTTTCCCT GAAGAGACGA AGCAAGTCGA AACTCAGAGT CGGAAAG AAAAAAAAAA |
| 6 | dna-ASBVd45-3′(GA)5 | CTTTCCCT GAAGAGACGA AGCAAGTCGA AACTCAGAGT CGGAAAG GAGAGAGAGA |
| 7 | dna-ASBVd45-3′(GA)10 | CTTTCCCT GAAGAGACGA AGCAAGTCGA AACTCAGAGT CGGAAAG GAGAGAGAGA GAGAGAGAGA |
| 8 | dna-ASBVd45-3′(GA)15 | CTTTCCCT GAAGAGACGA AGCAAGTCGA AACTCAGAGT CGGAAAG GAGAGAGAGA GAGAGAGAGA GAGAGAGAGA |
| 9 | dna-ASBVd45-3′(GA)20 | CTTTCCCT GAAGAGACGA AGCAAGTCGA AACTCAGAGT CGGAAAG GAGAGAGAGA GAGAGAGAGA GAGAGAGAGA GAGAGAGAGA |
| 10 | dna-ASBVd45-3′(GA)25 | CTTTCCCT GAAGAGACGA AGCAAGTCGA AACTCAGAGT CGGAAAG GAGAGAGAGA GAGAGAGAGA GAGAGAGAGA GAGAGAGAGA GAGAGAGAGA |
| Minerals | Area (BET)/m2 g−1 |
|---|---|
| Aerosil | 3.69 × 102 |
| Sepiolite | 3.51× 102 |
| Zirconium silicate | 4.82 × 100 |
| Montmorillonite | 2.29 × 102 |
| Conditions | k/min−1 * |
|---|---|
| Control | (80.0 ± 11.0) × 10−4 |
| In the presence of zirconium silicate | (2.46 ± 1.12) × 10−4 |
| In the presence of montmorillonite | (2.38 ± 0.55) × 10−4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kawamura, K.; Lambert, J.-F.; Ter-Ovanessian, L.M.P.; Vergne, J.; Hervé, G.; Maurel, M.-C. Life on Minerals: Binding Behaviors of Oligonucleotides on Zirconium Silicate and Its Inhibitory Activity for the Self-Cleavage of Hammerhead Ribozyme. Life 2022, 12, 1689. https://doi.org/10.3390/life12111689
Kawamura K, Lambert J-F, Ter-Ovanessian LMP, Vergne J, Hervé G, Maurel M-C. Life on Minerals: Binding Behaviors of Oligonucleotides on Zirconium Silicate and Its Inhibitory Activity for the Self-Cleavage of Hammerhead Ribozyme. Life. 2022; 12(11):1689. https://doi.org/10.3390/life12111689
Chicago/Turabian StyleKawamura, Kunio, Jean-François Lambert, Louis M. P. Ter-Ovanessian, Jacques Vergne, Guy Hervé, and Marie-Christine Maurel. 2022. "Life on Minerals: Binding Behaviors of Oligonucleotides on Zirconium Silicate and Its Inhibitory Activity for the Self-Cleavage of Hammerhead Ribozyme" Life 12, no. 11: 1689. https://doi.org/10.3390/life12111689
APA StyleKawamura, K., Lambert, J.-F., Ter-Ovanessian, L. M. P., Vergne, J., Hervé, G., & Maurel, M.-C. (2022). Life on Minerals: Binding Behaviors of Oligonucleotides on Zirconium Silicate and Its Inhibitory Activity for the Self-Cleavage of Hammerhead Ribozyme. Life, 12(11), 1689. https://doi.org/10.3390/life12111689