
Does a Single Exposure to General Anesthesia Have a Cumulative Effect on the Developing Brain after Mild Perinatal Asphyxia?

 ,
, 
 , , ,
, , ,  and
and 
Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
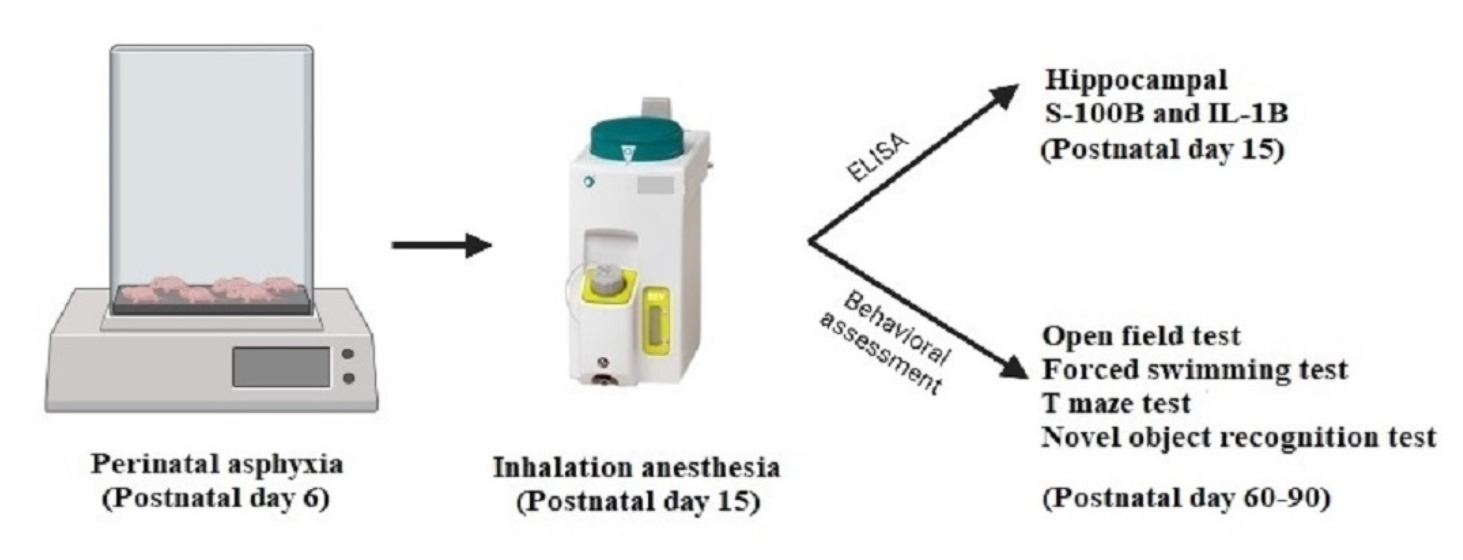
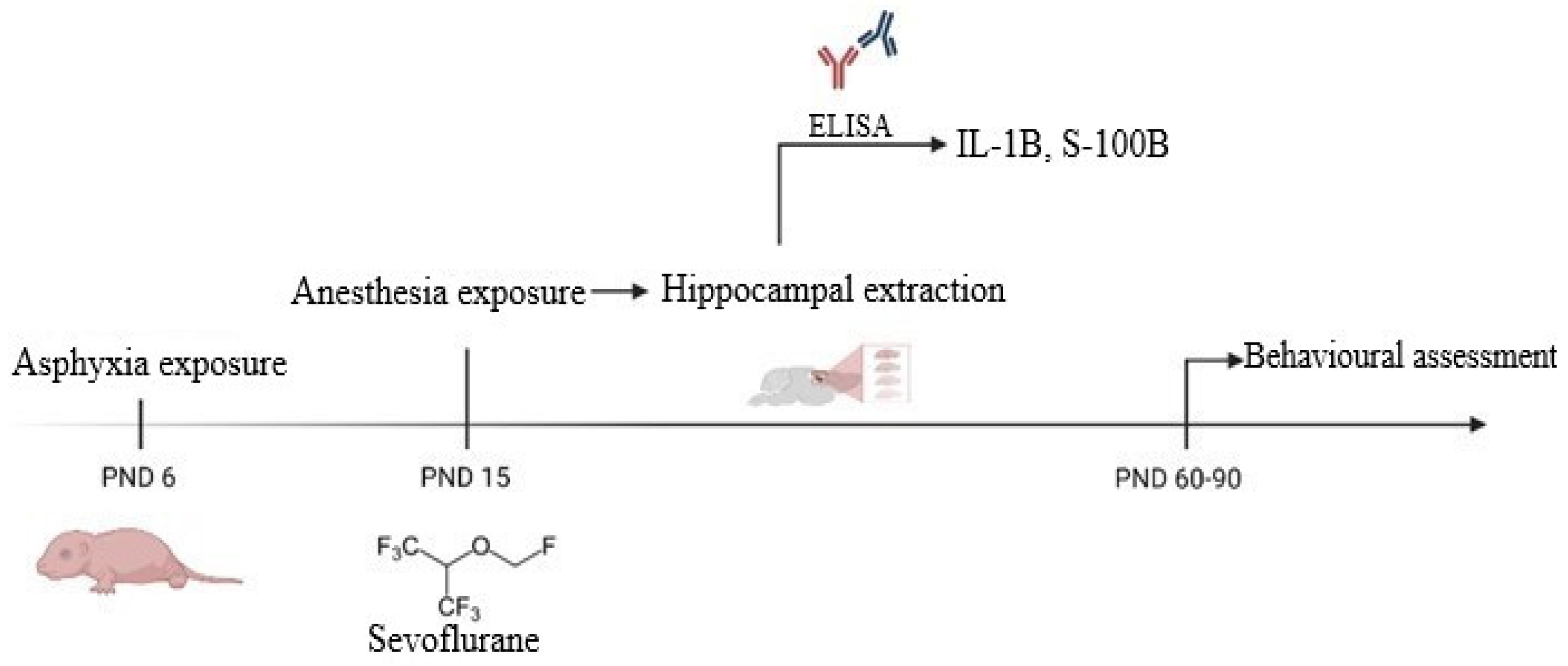
2.1. Asphyxia Exposure
2.2. General Anesthesia Exposure
2.3. Experimental Groups
2.4. Hippocampal Neuroinflammation and Injury Assessment by ELISA
2.5. Behavioral and Locomotor Assessment
2.6. Statistical Analysis
3. Results
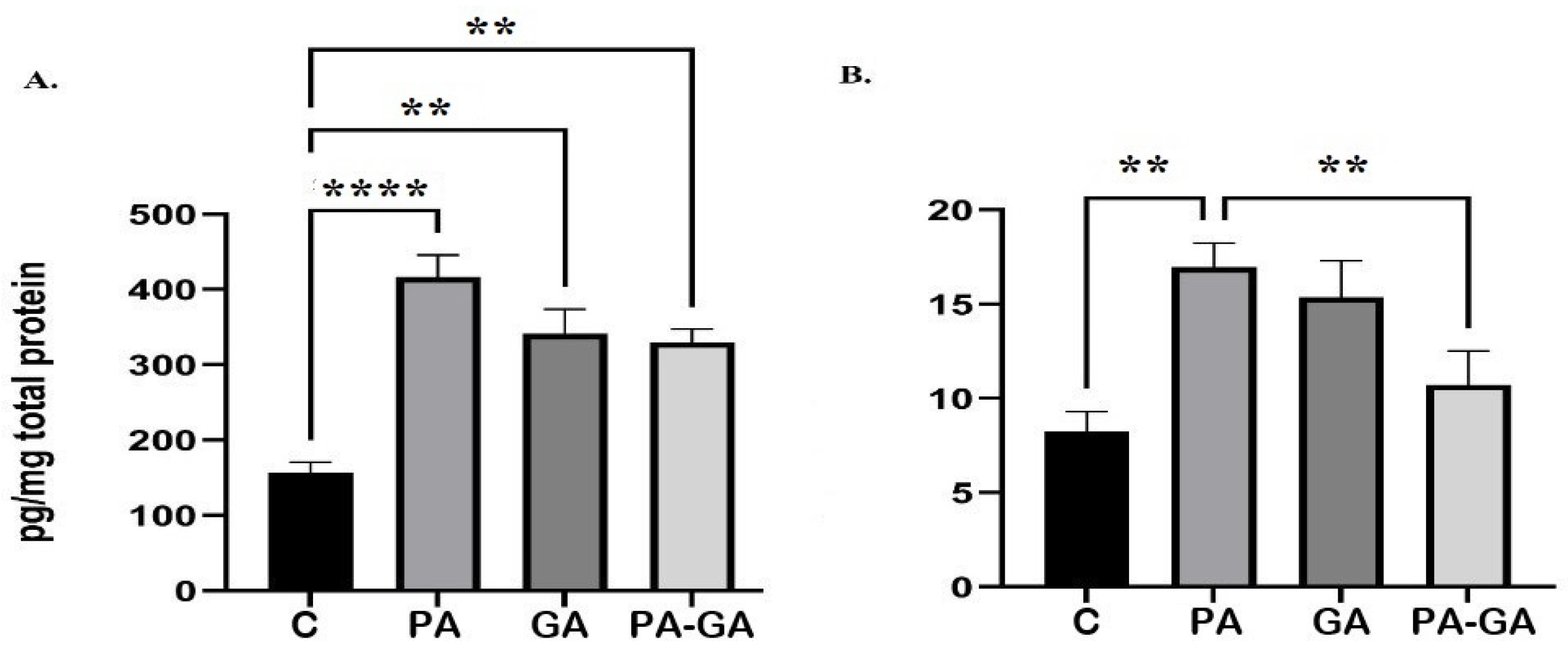
3.1. Hippocampal Concentration of S-100B and IL-1B
3.2. Behavioral and Locomotor Assessment
4. Discussion
Study Limitations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kornfeld, B.; Wild, B.M. Accounting for General Anesthesia: A Pre-Op Primer and Post-Op Factor. Pediatr. Ann. 2021, 50, e187–e189. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Michaelis, E. Selective Neuronal Vulnerability to Oxidative Stress in the Brain. Front. Aging Neurosci. 2010, 2, 12. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.-T.; Leu, D.; Zou, Y. Oxidative Stress and Redox Regulation on Hippocampal-Dependent Cognitive Functions. Arch. Biochem. Biophys. 2015, 576, 2–7. [Google Scholar] [CrossRef] [PubMed]
- Santos, I.M.S.; Tomé, A.R.; Saldanha, G.B.; Ferreira, P.M.P.; Militão, G.C.G.; de Freitas, R.M. Oxidative Stress in the Hippocampus during Experimental Seizures Can Be Ameliorated with the Antioxidant Ascorbic Acid. Oxid. Med. Cell. Longev. 2009, 2, 214–221. [Google Scholar] [CrossRef]
- Devroe, S.; Van der Veeken, L.; Bleeser, T.; Van der Merwe, J.; Meeusen, R.; Van de Velde, M.; Deprest, J.; Rex, S. The Effect of Xenon on Fetal Neurodevelopment Following Maternal Sevoflurane Anesthesia and Laparotomy in Rabbits. Neurotoxicol. Teratol. 2021, 87, 106994. [Google Scholar] [CrossRef]
- Ben-Ari, Y. GABA Excites and Sculpts Immature Neurons Well before Delivery: Modulation by GABA of the Development of Ventricular Progenitor Cells. Epilepsy Curr. 2007, 7, 167–169. [Google Scholar] [CrossRef]
- Kirmse, K.; Holthoff, K. Functions of GABAergic Transmission in the Immature Brain. e-Neuroforum 2017, 23, 27–33. [Google Scholar] [CrossRef]
- Kirmse, K.; Kummer, M.; Kovalchuk, Y.; Witte, O.W.; Garaschuk, O.; Holthoff, K. GABA Depolarizes Immature Neurons and Inhibits Network Activity in the Neonatal Neocortex in Vivo. Nat. Commun. 2015, 6, 7750. [Google Scholar] [CrossRef]
- Li, F.; Tsien, J.Z. Memory and the NMDA Receptors. N. Engl. J. Med. 2009, 361, 302–303. [Google Scholar] [CrossRef]
- Sha, H.; Peng, P.; Wei, G.; Wang, J.; Wu, Y.; Huang, H. Neuroprotective Effects of Dexmedetomidine on the Ketamine-Induced Disruption of the Proliferation and Differentiation of Developing Neural Stem Cells in the Subventricular Zone. Front. Pediatr. 2021, 9, 649284. [Google Scholar] [CrossRef]
- Raper, J.; Simon, H.K.; Kamat, P.P. Long-Term Evidence of Neonatal Anaesthesia Neurotoxicity Linked to Behavioural Phenotypes in Monkeys: Where Do We Go from Here? Br. J. Anaesth. 2021, 127, 343–345. [Google Scholar] [CrossRef] [PubMed]
- Remedios, E.; Sommerfield, D.; Fellingham, W.; Powers, N.; Stannage, K.; Hii, J.W.S. Paraspinal Desmoid Fibromatosis after Lumbar Epidural Analgesia. Anaesth. Rep. 2021, 9, e12129. [Google Scholar] [CrossRef]
- Biliškov, A.N.; Ivančev, B.; Pogorelić, Z. Effects on Recovery of Pediatric Patients Undergoing Total Intravenous Anesthesia with Propofol versus Ketofol for Short—Lasting Laparoscopic Procedures. Children 2021, 8, 610. [Google Scholar] [CrossRef] [PubMed]
- Aksenov, D.P.; Venkatasubramanian, P.N.; Miller, M.J.; Dixon, C.J.; Li, L.; Wyrwicz, A.M. Effects of Neonatal Isoflurane Anesthesia Exposure on Learning-Specific and Sensory Systems in Adults. Sci. Rep. 2020, 10, 13832. [Google Scholar] [CrossRef] [PubMed]
- Loepke, A.W. Developmental Neurotoxicity of Sedatives and Anesthetics: A Concern for Neonatal and Pediatric Critical Care Medicine? Pediatr. Crit. Care Med. 2010, 11, 217–226. [Google Scholar] [CrossRef]
- Isac, S.; Panaitescu, A.M.; Iesanu, M.I.; Zeca, V.; Cucu, N.; Zagrean, L.; Peltecu, G.; Zagrean, A.-M. Maternal Citicoline-Supplemented Diet Improves the Response of the Immature Hippocampus to Perinatal Asphyxia in Rats. Neonatology 2020, 117, 729–735. [Google Scholar] [CrossRef] [PubMed]
- Helmy, M.M.; Ruusuvuori, E.; Watkins, P.V.; Voipio, J.; Kanold, P.O.; Kaila, K. Acid Extrusion via Blood–Brain Barrier Causes Brain Alkalosis and Seizures after Neonatal Asphyxia. Brain 2012, 135, 3311–3319. [Google Scholar] [CrossRef][Green Version]
- Isac, S.; Panaitescu, A.M.; Iesanu, M.; Grigoras, I.F.; Totan, A.; Udriste, A.; Cucu, N.; Peltecu, G.; Zagrean, L.; Zagrean, A.-M. Maternal High-Fat Diet Modifies the Immature Hippocampus Vulnerability to Perinatal Asphyxia in Rats. Neonatology 2018, 114, 355–361. [Google Scholar] [CrossRef]
- Simpson, J.; Kelly, J.P. An Investigation of Whether There Are Sex Differences in Certain Behavioural and Neurochemical Parameters in the Rat. Behav. Brain Res. 2012, 229, 289–300. [Google Scholar] [CrossRef]
- Lovick, T.A.; Zangrossi, H. Effect of Estrous Cycle on Behavior of Females in Rodent Tests of Anxiety. Front. Psychiatry 2021, 12, 711065. [Google Scholar] [CrossRef]
- Lueptow, L.M. Novel Object Recognition Test for the Investigation of Learning and Memory in Mice. J. Vis. Exp. 2017, 126, e55718. [Google Scholar] [CrossRef] [PubMed]
- Antunes, M.; Biala, G. The Novel Object Recognition Memory: Neurobiology, Test Procedure, and Its Modifications. Cogn. Process. 2012, 13, 93–110. [Google Scholar] [CrossRef] [PubMed]
- Jablonski, S.A.; Schreiber, W.B.; Westbrook, S.R.; Brennan, L.E.; Stanton, M.E. Determinants of Novel Object and Location Recognition during Development. Behav. Brain Res. 2013, 256, 140–150. [Google Scholar] [CrossRef] [PubMed]
- Bogdanova, O.V.; Kanekar, S.; D’Anci, K.E.; Renshaw, P.F. Factors Influencing Behavior in the Forced Swim Test. Physiol. Behav. 2013, 118, 227–239. [Google Scholar] [CrossRef] [PubMed]
- Can, A.; Dao, D.T.; Arad, M.; Terrillion, C.E.; Piantadosi, S.C.; Gould, T.D. The Mouse Forced Swim Test. J. Vis. Exp. 2012, 59, e3638. [Google Scholar] [CrossRef] [PubMed]
- Yankelevitch-Yahav, R.; Franko, M.; Huly, A.; Doron, R. The Forced Swim Test as a Model of Depressive-like Behavior. J. Vis. Exp. 2015, 97, e52587. [Google Scholar] [CrossRef]
- Uzianbaeva, L.; Yan, Y.; Joshi, T.; Yin, N.; Hsu, C.-D.; Hernandez-Andrade, E.; Mehrmohammadi, M. Methods for Monitoring Risk of Hypoxic Damage in Fetal and Neonatal Brains: A Review. FDT 2022, 49, 1–24. [Google Scholar] [CrossRef]
- Min, Y.-J.; Ling, E.-A.; Li, F. Immunomodulatory Mechanism and Potential Therapies for Perinatal Hypoxic-Ischemic Brain Damage. Front. Pharmacol. 2020, 11, 580428. [Google Scholar] [CrossRef]
- Brandt, M.J.V.; Nijboer, C.H.; Nessel, I.; Mutshiya, T.R.; Michael-Titus, A.T.; Counotte, D.S.; Schipper, L.; van der Aa, N.E.; Benders, M.J.N.L.; de Theije, C.G.M. Nutritional Supplementation Reduces Lesion Size and Neuroinflammation in a Sex-Dependent Manner in a Mouse Model of Perinatal Hypoxic-Ischemic Brain Injury. Nutrients 2021, 14, 176. [Google Scholar] [CrossRef]
- Frank, M.G.; Weber, M.D.; Watkins, L.R.; Maier, S.F. Stress-Induced Neuroinflammatory Priming: A Liability Factor in the Etiology of Psychiatric Disorders. Neurobiol. Stress 2015, 4, 62–70. [Google Scholar] [CrossRef]
- Shen, X.; Dong, Y.; Xu, Z.; Wang, H.; Miao, C.; Soriano, S.G.; Sun, D.; Baxter, M.G.; Zhang, Y.; Xie, Z. Selective Anesthesia-Induced Neuroinflammation in Developing Mouse Brain and Cognitive Impairment. Anesthesiology 2013, 118, 502–515. [Google Scholar] [CrossRef] [PubMed]
- Michetti, F.; Corvino, V.; Geloso, M.C.; Lattanzi, W.; Bernardini, C.; Serpero, L.; Gazzolo, D. The S100B Protein in Biological Fluids: More than a Lifelong Biomarker of Brain Distress. J. Neurochem. 2012, 120, 644–659. [Google Scholar] [CrossRef] [PubMed]
- Locci, E.; Chighine, A.; Noto, A.; Ferino, G.; Baldi, A.; Varvarousis, D.; Xanthos, T.; De-Giorgio, F.; Stocchero, M.; d’Aloja, E. Metabolomics Improves the Histopathological Diagnosis of Asphyxial Deaths: An Animal Proof-of-Concept Model. Sci. Rep. 2021, 11, 10102. [Google Scholar] [CrossRef] [PubMed]
- Kleindienst, A.; Hesse, F.; Bullock, M.R.; Buchfelder, M. The Neurotrophic Protein S100B: Value as a Marker of Brain Damage and Possible Therapeutic Implications. Prog. Brain Res. 2007, 161, 317–325. [Google Scholar] [CrossRef] [PubMed]
- Panaitescu, A.M.; Isac, S.; Pavel, B.; Ilie, A.S.; Ceanga, M.; Totan, A.; Zagrean, L.; Peltecu, G.; Zagrean, A.M. Oxytocin reduces seizure burden and hippocampal injury in a rat model of perinatal asphyxia. Acta Endocrinol. (Buchar.) 2018, 14, 315–319. [Google Scholar] [CrossRef]
- Zhou, X.; Li, W.; Chen, X.; Yang, X.; Zhou, Z.; Lu, D.; Feng, X. Dose-Dependent Effects of Sevoflurane Exposure during Early Lifetime on Apoptosis in Hippocampus and Neurocognitive Outcomes in Sprague-Dawley Rats. Int. J. Physiol. Pathophysiol. Pharmacol. 2016, 8, 111–119. [Google Scholar]
- Simola, N.; Bustamante, D.; Pinna, A.; Pontis, S.; Morales, P.; Morelli, M.; Herrera-Marschitz, M. Acute Perinatal Asphyxia Impairs Non-Spatial Memory and Alters Motor Coordination in Adult Male Rats. Exp. Brain Res. 2008, 185, 595–601. [Google Scholar] [CrossRef]
- Daniel, D.G.; Daniel, N.G.; Daniel, D.T.; Flynn, L.C.; Allen, M.H. The Effect of Propofol on a Forced Swim Test in Mice at 24 Hours. Curr. Ther. Res. Clin. Exp. 2020, 92, 100590. [Google Scholar] [CrossRef]
- Herbst, L.S.; Gaigher, T.; Siqueira, A.A.; Joca, S.R.L.; Sampaio, K.N.; Beijamini, V. New Evidence for Refinement of Anesthetic Choice in Procedures Preceding the Forced Swimming Test and the Elevated Plus-Maze. Behav. Brain Res. 2019, 368, 111897. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Isac, S.; Pavel, B.; Dobre, M.; Milanesi, E.; Matache, I.-M.; Paun, R.-M.; Klimko, A.; Iesanu, M.I.; Droc, G.; Zagrean, A.-M. Does a Single Exposure to General Anesthesia Have a Cumulative Effect on the Developing Brain after Mild Perinatal Asphyxia? Life 2022, 12, 1568. https://doi.org/10.3390/life12101568
Isac S, Pavel B, Dobre M, Milanesi E, Matache I-M, Paun R-M, Klimko A, Iesanu MI, Droc G, Zagrean A-M. Does a Single Exposure to General Anesthesia Have a Cumulative Effect on the Developing Brain after Mild Perinatal Asphyxia? Life. 2022; 12(10):1568. https://doi.org/10.3390/life12101568
Chicago/Turabian StyleIsac, Sebastian, Bogdan Pavel, Maria Dobre, Elena Milanesi, Irina-Mihaela Matache, Raul-Mihai Paun, Artsiom Klimko, Mara Ioana Iesanu, Gabriela Droc, and Ana-Maria Zagrean. 2022. "Does a Single Exposure to General Anesthesia Have a Cumulative Effect on the Developing Brain after Mild Perinatal Asphyxia?" Life 12, no. 10: 1568. https://doi.org/10.3390/life12101568
APA StyleIsac, S., Pavel, B., Dobre, M., Milanesi, E., Matache, I.-M., Paun, R.-M., Klimko, A., Iesanu, M. I., Droc, G., & Zagrean, A.-M. (2022). Does a Single Exposure to General Anesthesia Have a Cumulative Effect on the Developing Brain after Mild Perinatal Asphyxia? Life, 12(10), 1568. https://doi.org/10.3390/life12101568