
Comparison of Chicken Immune Responses to Immunization with Vaccine La Sota or ZG1999HDS Strain of Newcastle Disease Virus

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Trial Animals
2.2. Viral Preparations
2.3. Animal Procedures
2.4. Laboratory Procedures
2.4.1. Determination of Humoral Immunity by Haemagglutination Inhibition (HI) Assay
2.4.2. Determination of Cell-Mediated Immunity by Differentiation of the Total Number of Chicken Leukocytes and Their Subpopulations in Flow Cytometry
2.5. Statistical Data Analysis
3. Results
3.1. Overall Animal Health in the Experiment
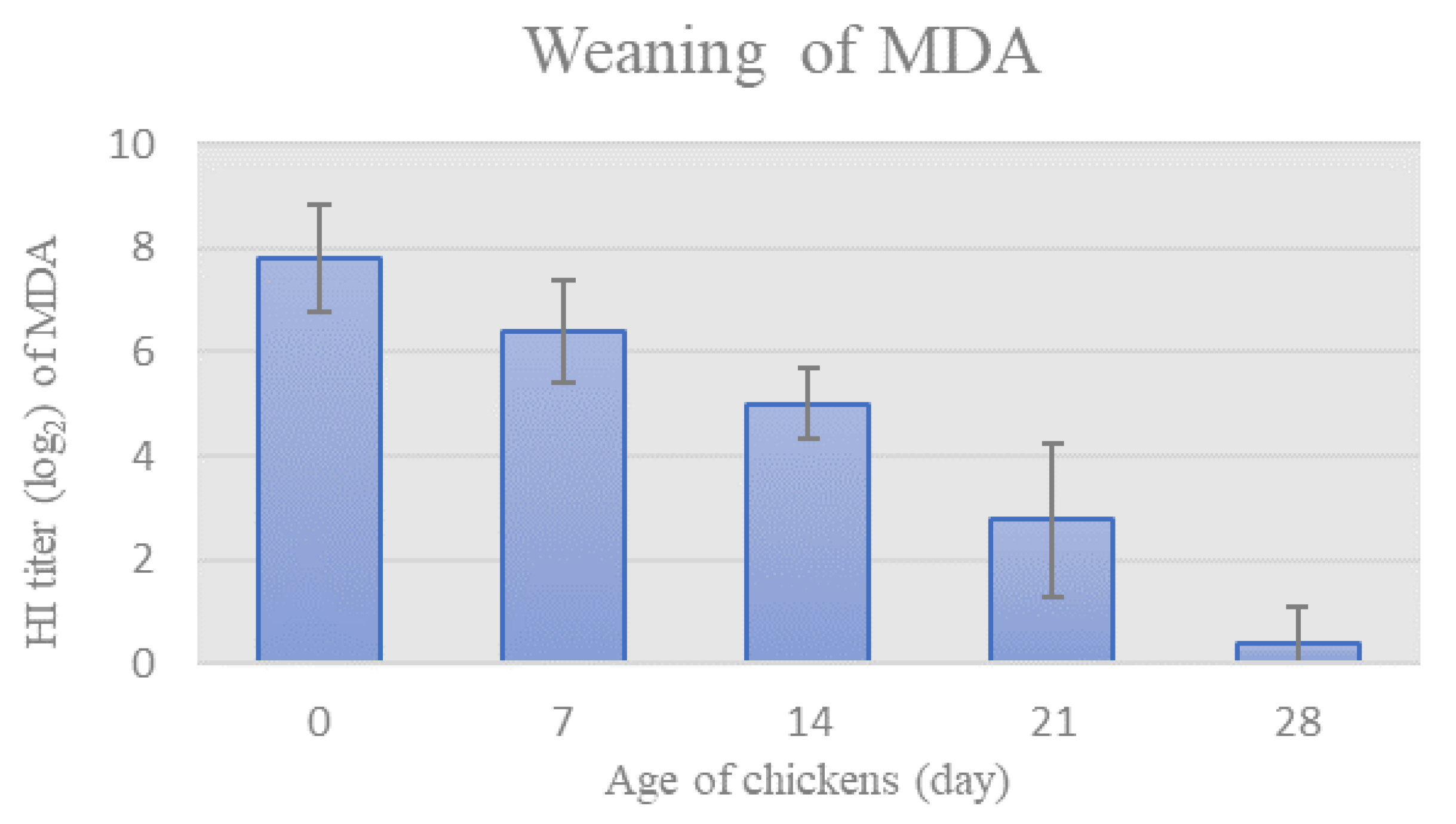
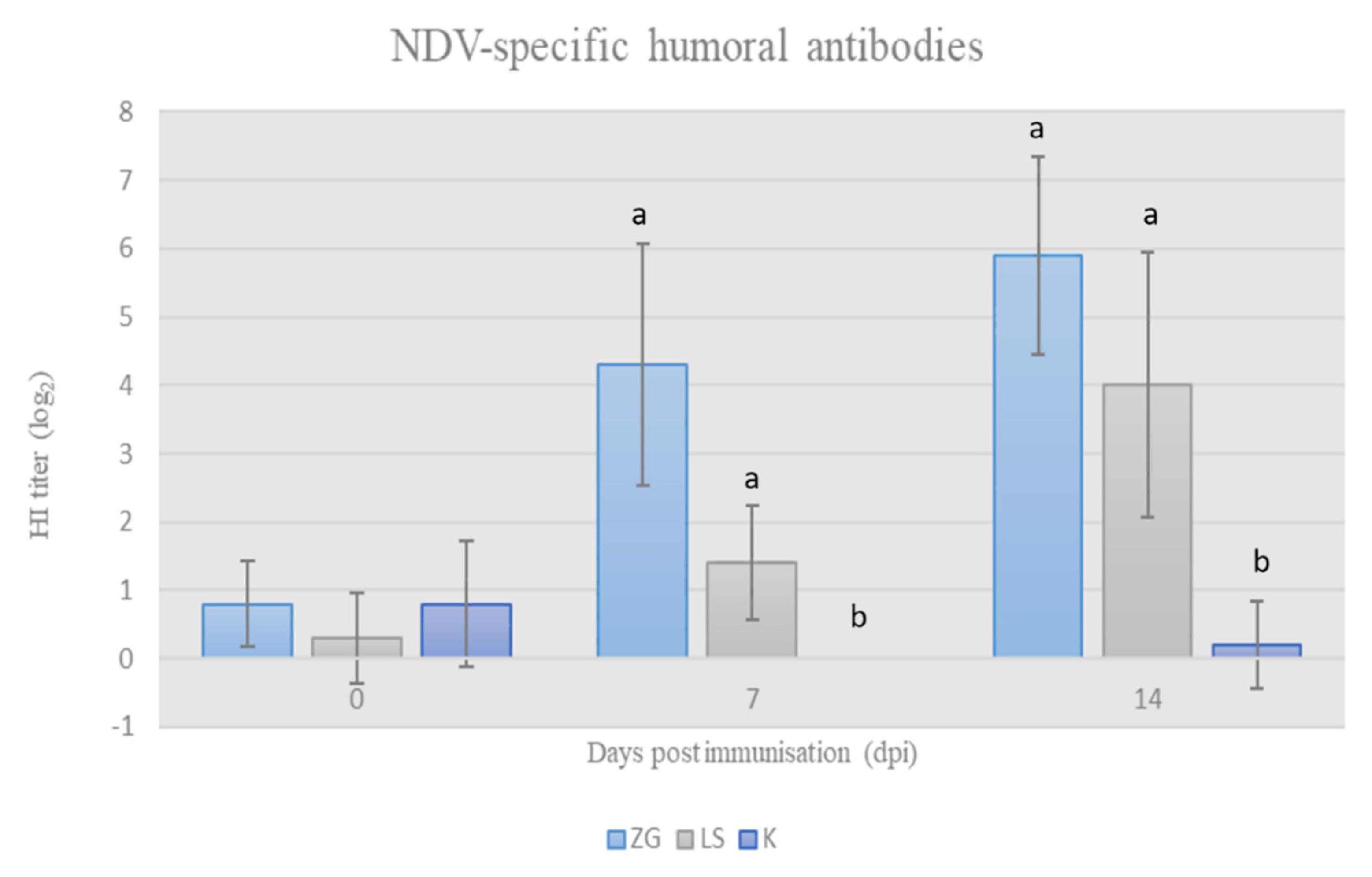
3.2. Humoral Immunity
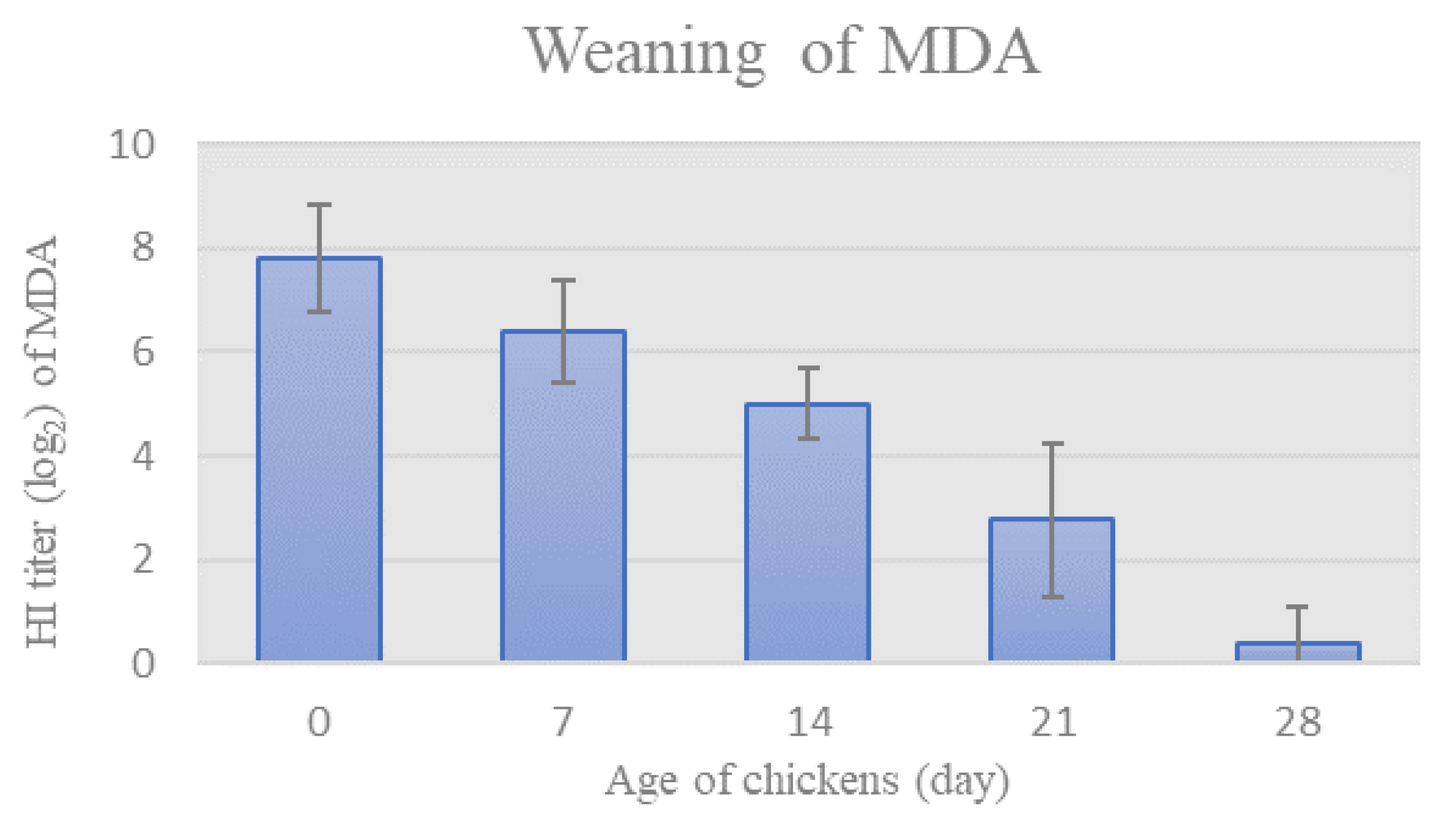
3.2.1. Weaning of MDA
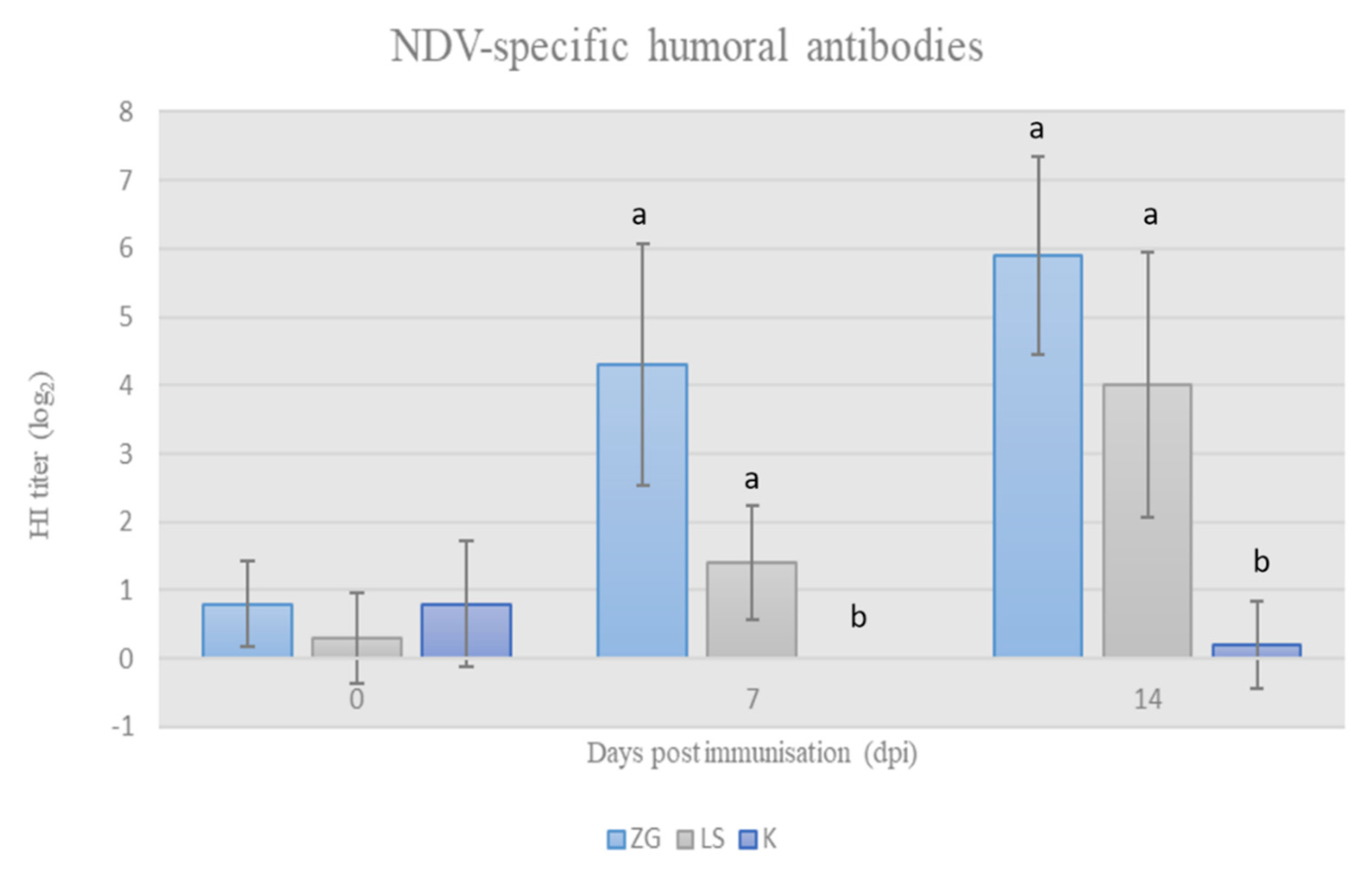
3.2.2. Acquired HI Antibodies
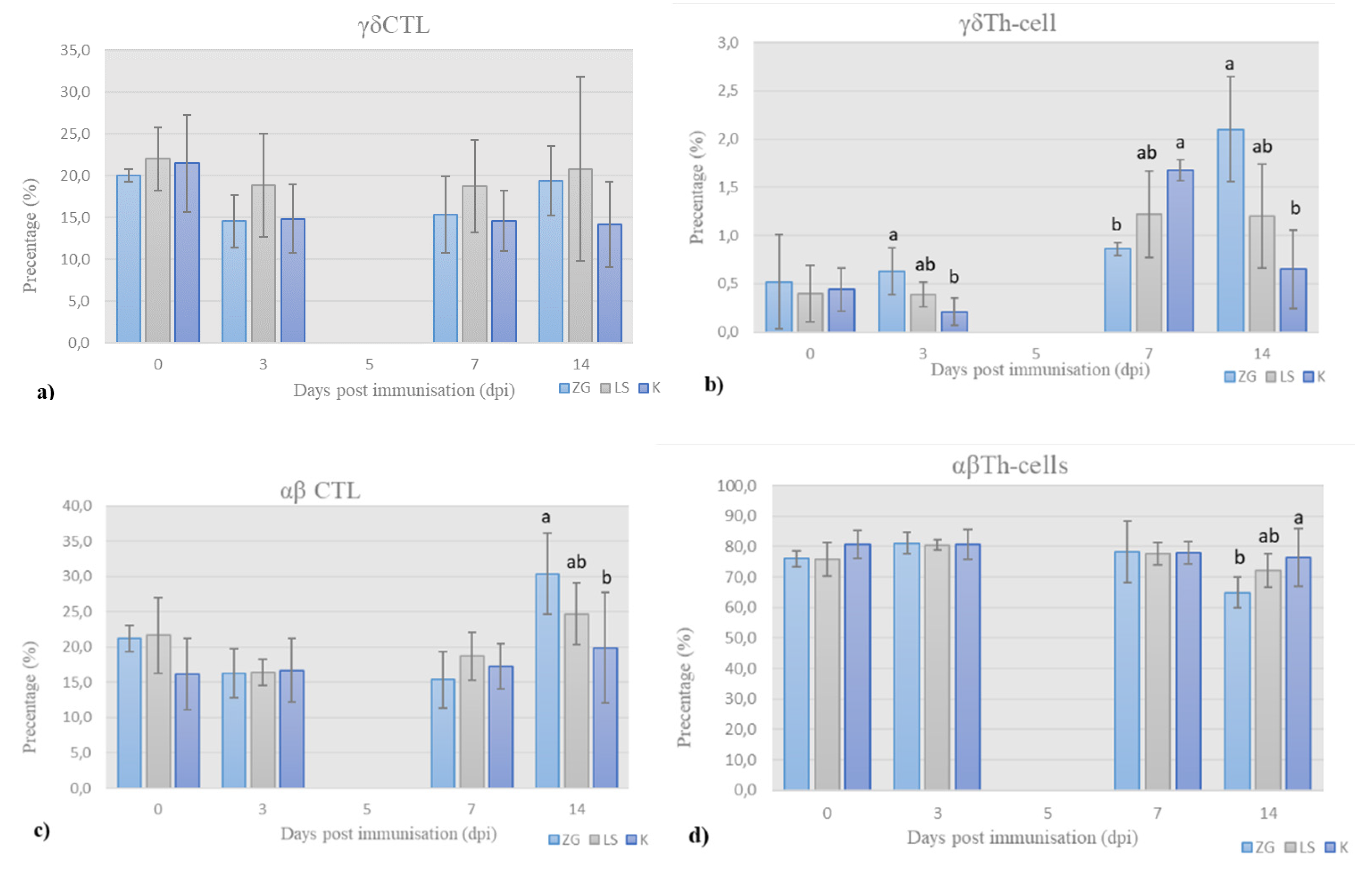
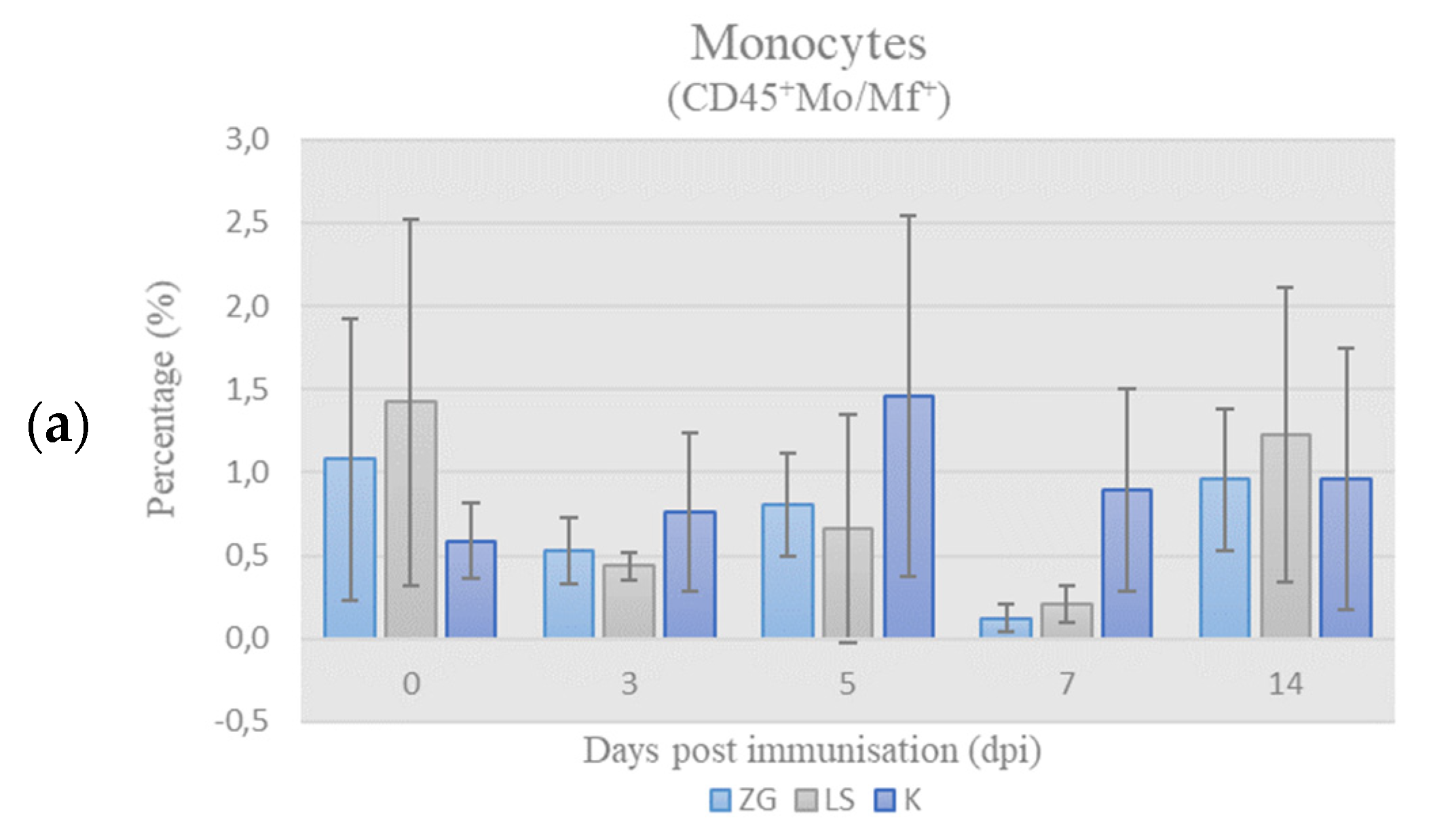
3.3. Cell-Mediated Immunity
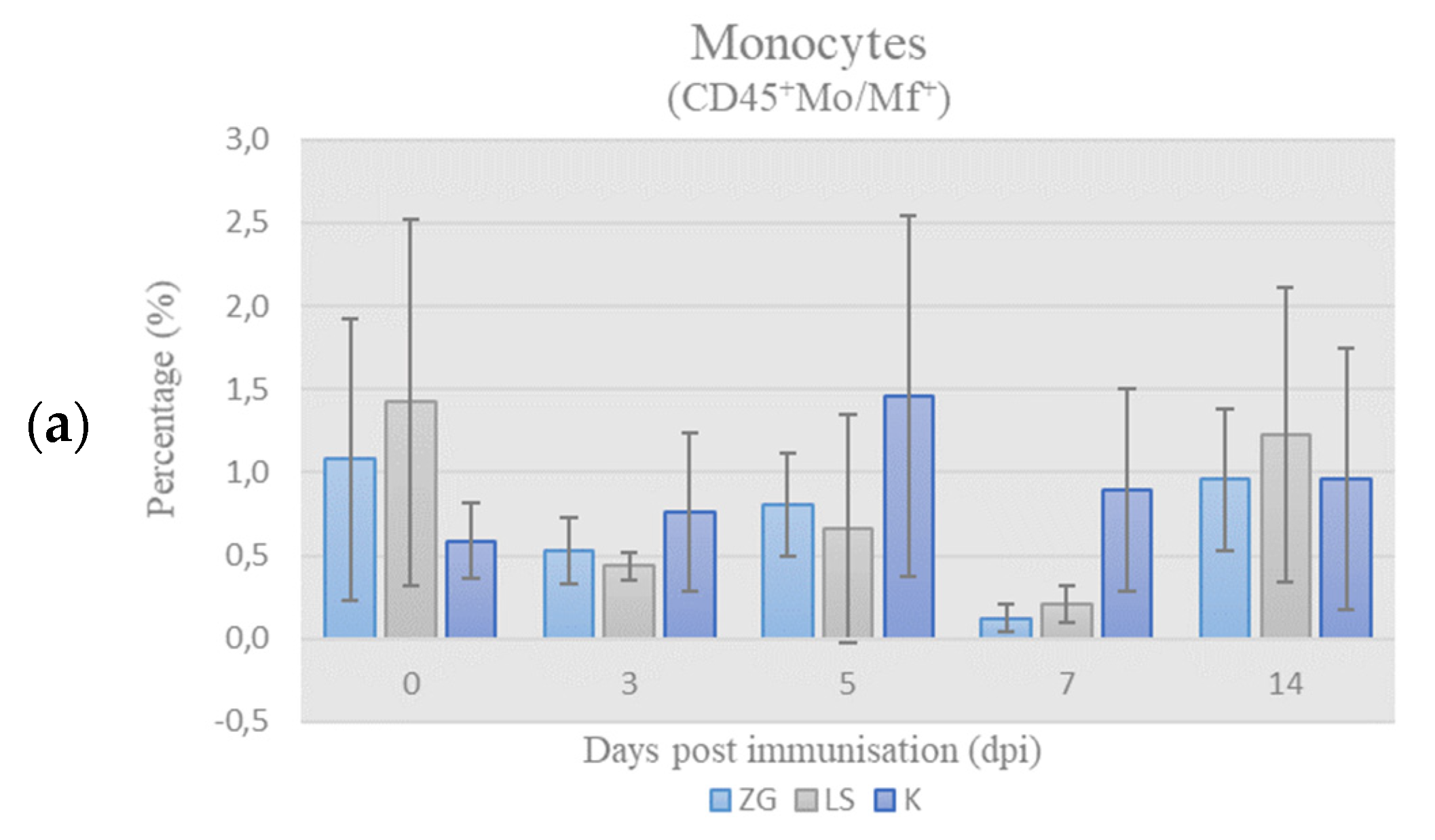
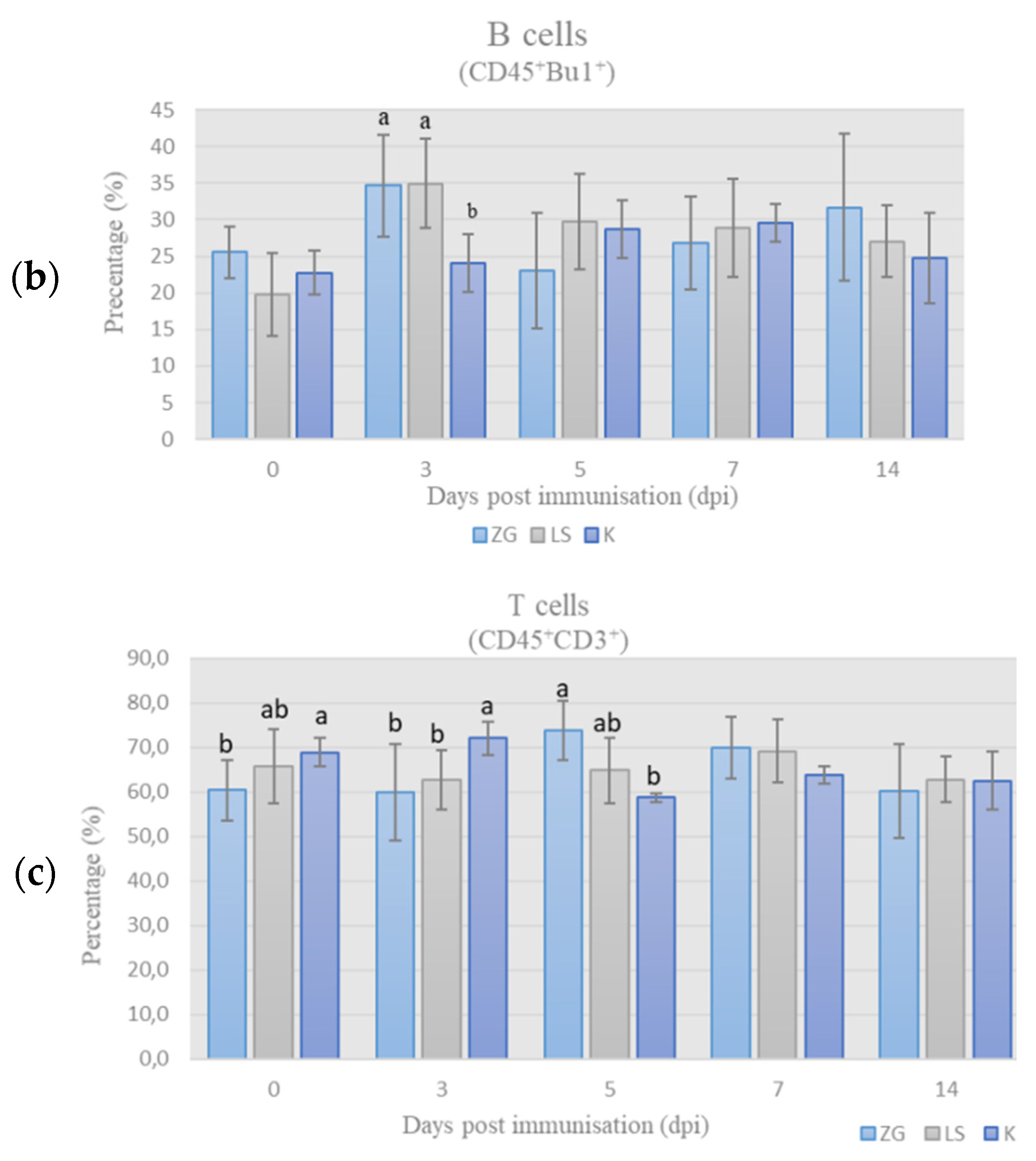
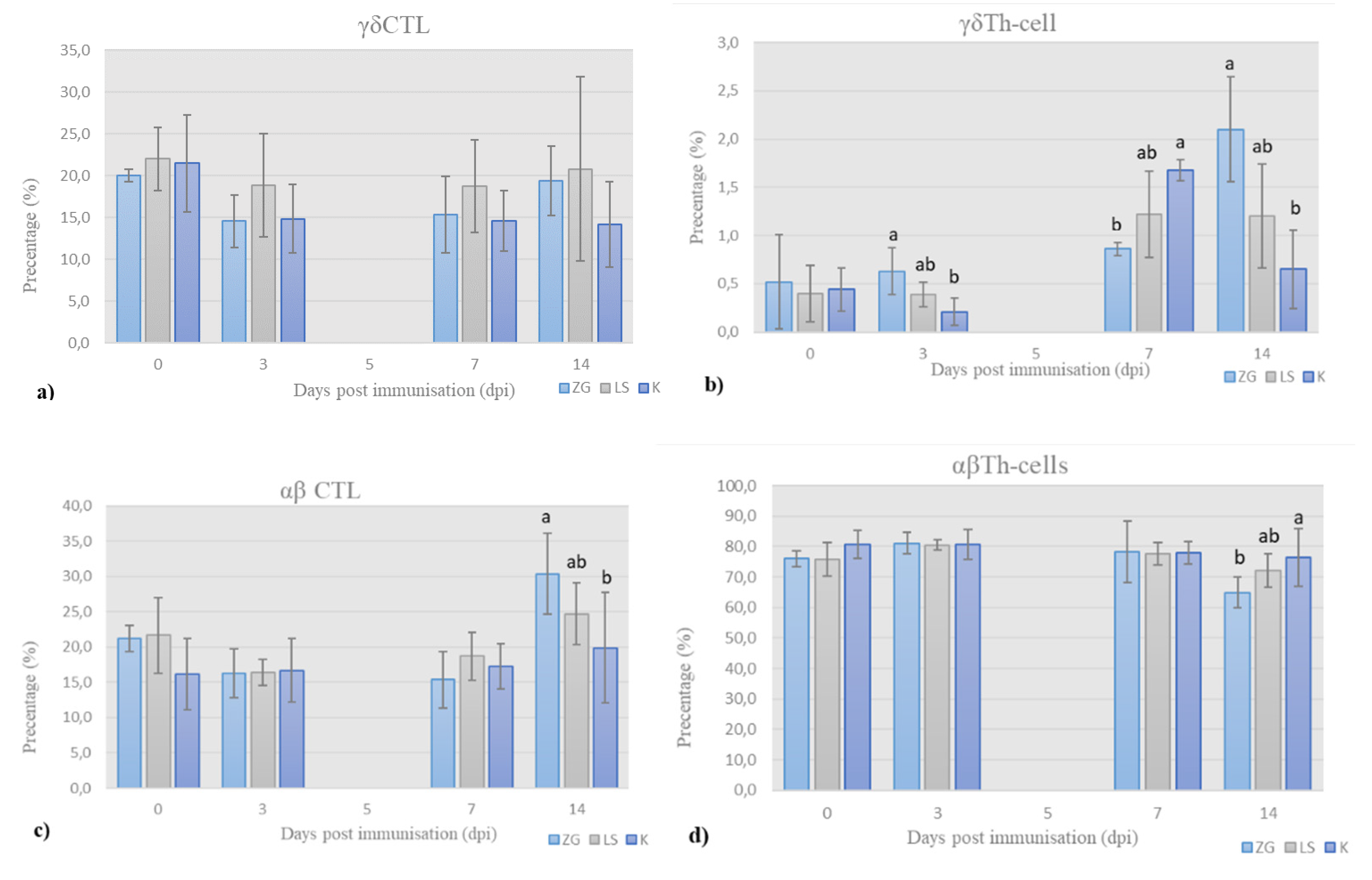
3.3.1. Immunophenotyping of Chicken PBMCs
Leukocyte-Panel (L-Panel)
T-Lymphocytes’ Panel (T-Panel)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Alexander, D.J. Gordon Memorial Lecture. Newcastle disease. Br. Poult. Sci. 2001, 42, 5–22. [Google Scholar] [CrossRef] [PubMed]
- Dimitrov, K.M.; Abolnik, C.; Afonso, C.L.; Albina, E.; Bahl, J.; Berg, M.; Briand, F.-X.; Brown, I.H.; Choi, K.-S.; Chvala, I.; et al. Updated unified phylogenetic classification system and revised nomenclature for Newcastle disease virus. Infect. Genet. Evol. 2019, 74, 103917. [Google Scholar] [CrossRef]
- Suarez, D.L.; Miller, P.J.; Koch, G.; Mundt, E.; Rautenschlein, S. Newcastle Disease, Other Avian Paramyxoviruses, and Avian Metapneumovirus Infections. In Diseases of Poultry, 14th ed.; David, E., Swayne, M.B., Catherine, M.L., Larry, R.M., Venugopal, N., David, L.S., de Wit, S., Grimes, T., Deirdre, J., Michelle, K., Eds.; Wiley-Blackwell: Hoboken, NJ, USA, 2020; pp. 109–166. [Google Scholar]
- Peeters, B.; Koch, G. Newcastle Disease Virus (Paramyxoviridae). In Encyclopedia of Virology; Elsevier: Amsterdam, The Netherlands, 2021; pp. 648–653. ISBN 9780128096338. [Google Scholar]
- Gallili, G.E.; Ben-Nathan, D. Newcastle disease vaccines. Biotechnol. Adv. 1998, 16, 343–366. [Google Scholar] [CrossRef]
- Liu, R.; Chen, C.; Huang, Y.; Cheng, L.; Lu, R.; Fu, G.; Shi, S.; Chen, H.; Wan, C.; Fu, Q.; et al. Microbiological identification and analysis of waterfowl livers collected from backyard farms in Southern China. J. Vet. Med. Sci. 2018, 80, 667–671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Igwe, A.O.; Agbakwuru, I.O. Haemagglutination inhibition antibody responses of pullet and broiler chickens (Gallus gallus domesticus) to Newcastle disease virus LaSota vaccination. Sokoto J. Vet. Sci. 2019, 16, 36. [Google Scholar] [CrossRef]
- Bello, M.B.; Yusoff, K.; Ideris, A.; Hair-Bejo, M.; Peeters, B.P.H.; Omar, A.R. Diagnostic and Vaccination Approaches for Newcastle Disease Virus in Poultry: The Current and Emerging Perspectives. BioMed Res. Int. 2018, 2018, 7278459. [Google Scholar] [CrossRef]
- Gowthaman, V.; Ganesan, V.; Gopala Krishna Murthy, T.R.; Nair, S.; Yegavinti, N.; Saraswathy, P.V.; Suresh Kumar, G.; Udhayavel, S.; Senthilvel, K.; Subbiah, M. Molecular phylogenetics of Newcastle disease viruses isolated from vaccinated flocks during outbreaks in Southern India reveals circulation of a novel sub-genotype. Transbound. Emerg. Dis. 2019, 66, 363–372. [Google Scholar] [CrossRef] [Green Version]
- Abolnik, C.; Mubamba, C.; Dautu, G.; Gummow, B. Complete Genome Sequence of a Newcastle Disease Genotype XIII Virus Isolated from Indigenous Chickens in Zambia. Genome Announc. 2017, 5, 2–3. [Google Scholar] [CrossRef] [Green Version]
- Abolnik, C. History of Newcastle disease in South Africa. Onderstepoort J. Vet. Res. 2017, 84, e1–e7. [Google Scholar] [CrossRef] [PubMed]
- Kapczynski, D.R.; Afonso, C.L.; Miller, P.J. Immune responses of poultry to Newcastle disease virus. Dev. Comp. Immunol. 2013, 41, 447–453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Garib, S.O.; Gielkens, A.L.J.; Gruys, E.; Koch, G. Review of Newcastle disease virus with particular references to immunity and vaccination. Worlds Poult. Sci. J. 2003, 59, 185–200. [Google Scholar] [CrossRef]
- Alexander, D.J.; Senne, D.A. Newcastle Disease. In Diseases of Poultry, 12th ed.; Aly, M., Saif, A.M., Fadly, J.R., Glisson, L.R., McDougald, L.K., Nolan, D.E.S., Eds.; Iowa State University Press: Ames, IA, USA, 2008; pp. 75–116. [Google Scholar]
- Balenović, M.; Popović, M.; Savić, V.; Kezić, D.; Vlahović, K.; Dovč, A.; Bezrok, G.; Popović, I.; Valpotić, I. Kvantitativna imunofenotipizacija T-limfocita periferne krvi kokoši nesilica. Prax. Vet. 2007, 55, 33–39. [Google Scholar]
- Dalgaard, T.S.; Norup, L.R.; Pedersen, A.R.; Handberg, K.J.; Jørgensen, P.H.; Juul-Madsen, H.R. Flow cytometric assessment of chicken T cell-mediated immune responses after Newcastle disease virus vaccination and challenge. Vaccine 2010, 28, 4506–4514. [Google Scholar] [CrossRef]
- Bidin, M.; Mazija, H. Immunogenicity of the field strain NDV Zg-2000 administered to the SPF chickens. In Proceedings of Pultry Days 2009; Mirta, B., Ed.; Poultry Centre: Zagreb, Croatia, 2009; pp. 241–245. [Google Scholar]
- Cvetić, Ž.; Nedeljković, G.; Jergović, M.; Bendelja, K.; Mazija, H.; Gottstein, Ž. Immunogenicity of Newcastle disease virus strain ZG1999HDS applied oculonasally or by means of nebulization to day-old chicks. Poult. Sci. 2021, 100, 101001. [Google Scholar] [CrossRef]
- Runjić, I. Genomic Characteristics of the Strain ZG—2000 of the Virus of Newcastle Diseas; Faculty of Science, University of Zagreb: Zagreb, Croatia, 2006. [Google Scholar]
- Stupar, M. Evidence of Horizontal Spread of Newcastle Disease Virus Strain ZG1999HDS after iv Application in Chickens; Faculty of Veterinary Medicine, University of Zagreb: Zagreb, Croatia, 2012. [Google Scholar]
- Nedeljković, G. (Veterinarski fakultet) Genomic Characterisation and Phylogenetic Analysis of a Newcastle Disease Virus Isolate (ZG1999HDS) from an Outbreak in 1999 in Croatia; Uppsala University: Uppsala, Sweden, 2011. [Google Scholar]
- Mazija, H.; Gottstein, Ž.; Ivanković, S.; Čović, D. Lentogenic cytolitic strain of the Newcastle Disease Virus isolated in Croatia. In IX Symposium Poultry Days 2011 with International Participation Croatia, Šibenik, 11–14 May 2011; Balenović, M., Ed.; Poultry Centre: Zagreb, Croatia, 2011; pp. 48–58. [Google Scholar]
- Anonymous. (Genera) Pestikal® Cjepivo Protiv Newcastleske Bolesti; Hrvatski veterinarski Institut: Rakov Potok, Croatia, 2013; Volume 10. [Google Scholar]
- Allan, W.; Gough, R. A standard haemagglutination inhibition test for Newcastle disease. (1). A comparison of macro and micro methods. Vet. Rec. 1974, 95, 120–123. [Google Scholar] [CrossRef]
- Anonnymous. Newcastle Disease (Infection with Newcastle disease virus). In Manual of Diagnostic Tests and Vaccines for Terrestrial Animals, 8th ed.; Organisation for Animal Health: Paris, France, 2018; Volume 3, pp. 1–22. [Google Scholar]
- Jergović, M.; Nedeljković, G.; Cvetić, Ž.; Gottstein, Ž.; Bendelja, K. Combined dextran and ficoll separation yields pure populations of chicken peripheral blood mononuclear cells—Short communicationia. Vet. Arh. 2017, 87, 623–631. [Google Scholar] [CrossRef]
- Rauw, F.; Gardin, Y.; Palya, V.; van Borm, S.; Gonze, M.; Lemaire, S.; van den Berg, T.; Lambrecht, B. Humoral, cell-mediated and mucosal immunity induced by oculo-nasal vaccination of one-day-old SPF and conventional layer chicks with two different live Newcastle disease vaccines. Vaccine 2009, 27, 3631–3642. [Google Scholar] [CrossRef]
- Sasipreeyajan, J.; Pohuang, T.; Sirikobkul, N. Efficacy of Different Vaccination Programs against Newcastle Disease Virus Challenge in Broiler Chickens. Thai J. Vet. Med. 2012, 42, 431–437. [Google Scholar]
- Lambrecht, B.; Gonze, M.; Meulemans, G.; van den Berg, T.P. Assessment of the cell-mediated immune response in chickens by detection of chicken interferon-gamma in response to mitogen and recall Newcastle disease viral antigen stimulation. Avian Pathol. 2004, 33, 343–350. [Google Scholar] [CrossRef]
- Norup, L.R.; Dalgaard, T.S.; Pedersen, A.R.; Juul-Madsen, H.R. Assessment of Newcastle disease-specific T cell proliferation in different inbred MHC chicken lines. Scand. J. Immunol. 2011, 74, 23–30. [Google Scholar] [CrossRef]
- Kaiser, P.; Stäheli, P. Avian Cytokines and Chemokines. In Avian Immunology; Elsevier: Amsterdam, The Netherlands, 2014; pp. 189–204. [Google Scholar]
- Tizard, I. Veterinary Immunology, 9th ed.; Tizard, I., Ed.; Elsevier Saunders: St. Louis, MO, USA, 2013; ISBN 9780323291699. [Google Scholar]
- Kristeen-Teo, Y.W.; Yeap, S.K.; Tan, S.W.; Omar, A.R.; Ideris, A.; Tan, S.G.; Alitheen, N.B. The effects of different velogenic NDV infections on the chicken bursa of Fabricius. BMC Vet. Res. 2017, 13, 151. [Google Scholar] [CrossRef] [Green Version]
- Degen, W.G.J.; Daal, N.V.; Rothwell, L.; Kaiser, P.; Schijns, V.E.J.C. Th1/Th2 polarization by viral and helminth infection in birds. Vet. Microbiol. 2005, 105, 163–167. [Google Scholar] [CrossRef] [Green Version]
- Kaiser, P. Advances in avian immunology-prospects for disease control: A review. Avian Pathol. 2010, 39, 309–324. [Google Scholar] [CrossRef]
- Zitzmann, C.; Kaderali, L. Mathematical Analysis of Viral Replication Dynamics and Antiviral Treatment Strategies: From Basic Models to Age-Based Multi-Scale Modeling. Front. Microbiol. 2018, 9, 1546. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Leucocyte Marker (CD) | Clone | Fluorescent Dye | Isotype | Cell Population with Marker |
|---|---|---|---|---|
| CD45 | LT-40 | APC | Mouse IgMκ | All leucocytes |
| Mo/Mf | KUL-01 | R-PE | Mouse IgG1κ | Macrophages and monocytes |
| Bu-1 | AV 20 | FITC | Mouse IgG1κ | B-cells, Bursal cells |
| CD3 | CT-3 | SPRD | Mouse IgG1κ | T cells |
| CD4 | CT-4 | R-PE | Mouse IgG1κ | T helper (Th) cells |
| CD8α | EP-72 | FITC | Mouse IgG2bκ | T cytotoxic (Tc) cells |
| γδ TCR | TCR 1 | UNLB(APC) | Mouse gG1κ | T cells with γδ surface receptor |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nedeljković, G.; Mazija, H.; Cvetić, Ž.; Jergović, M.; Bendelja, K.; Gottstein, Ž. Comparison of Chicken Immune Responses to Immunization with Vaccine La Sota or ZG1999HDS Strain of Newcastle Disease Virus. Life 2022, 12, 72. https://doi.org/10.3390/life12010072
Nedeljković G, Mazija H, Cvetić Ž, Jergović M, Bendelja K, Gottstein Ž. Comparison of Chicken Immune Responses to Immunization with Vaccine La Sota or ZG1999HDS Strain of Newcastle Disease Virus. Life. 2022; 12(1):72. https://doi.org/10.3390/life12010072
Chicago/Turabian StyleNedeljković, Gordana, Hrvoje Mazija, Željko Cvetić, Mladen Jergović, Krešo Bendelja, and Željko Gottstein. 2022. "Comparison of Chicken Immune Responses to Immunization with Vaccine La Sota or ZG1999HDS Strain of Newcastle Disease Virus" Life 12, no. 1: 72. https://doi.org/10.3390/life12010072
APA StyleNedeljković, G., Mazija, H., Cvetić, Ž., Jergović, M., Bendelja, K., & Gottstein, Ž. (2022). Comparison of Chicken Immune Responses to Immunization with Vaccine La Sota or ZG1999HDS Strain of Newcastle Disease Virus. Life, 12(1), 72. https://doi.org/10.3390/life12010072