
Simulated Microgravity Promotes Horizontal Gene Transfer of Antimicrobial Resistance Genes between Bacterial Genera in the Absence of Antibiotic Selective Pressure




Abstract
:1. Introduction
2. Methods
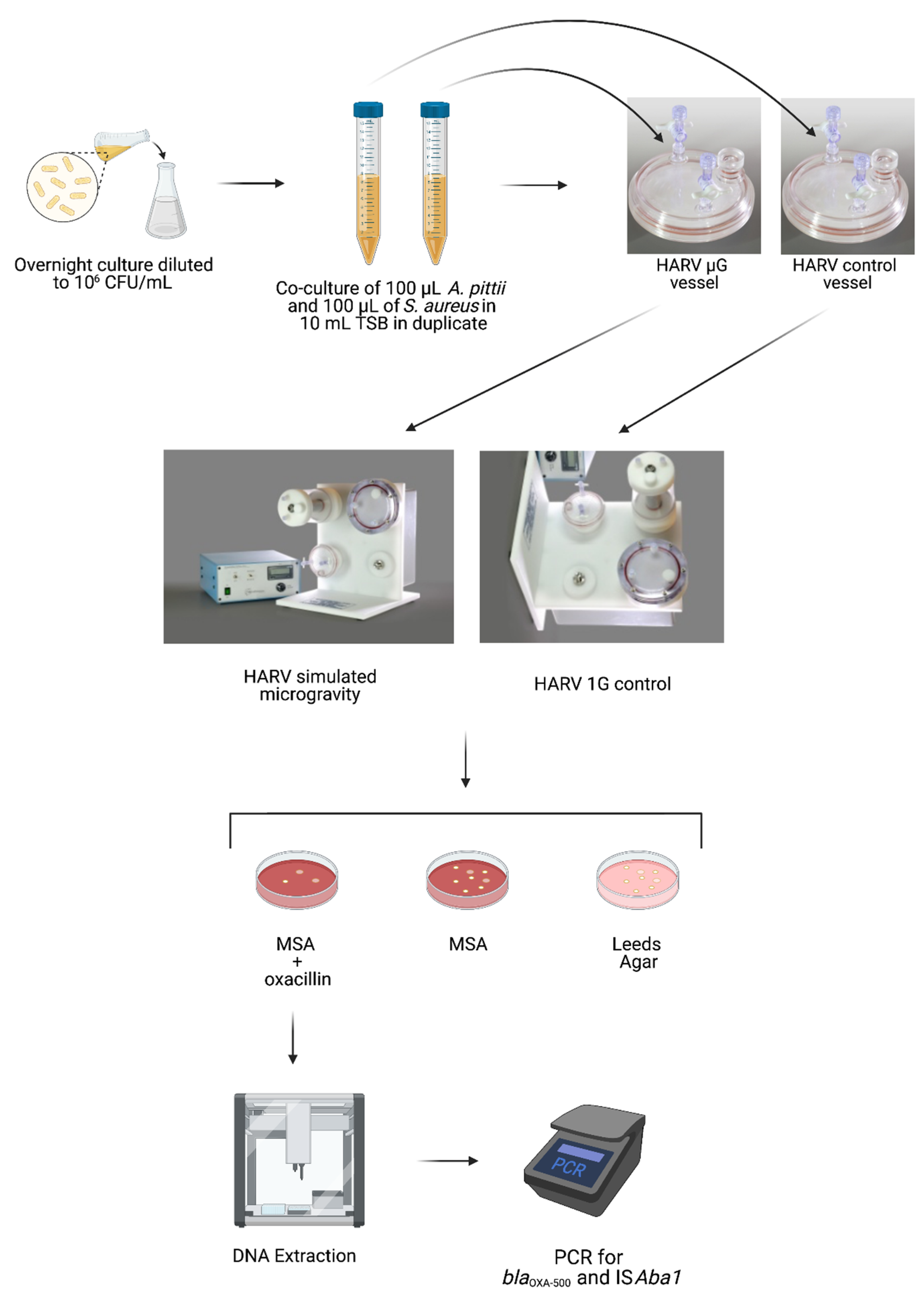
2.1. Co-Culture Experiments
2.2. Isolation of Colonies after HARV Growth
2.3. Designing Primers
2.4. PCR
- blaOXA-500_F (5′-CCGAGTTGTTCCAATCCCTTAT-3′) and
- blaOXA-500_R (5′-ATATGTTCCCGCCTCTACCT-3′) to amplify blaOXA-500
- and ISAba1_F (5′-ATGCAGCGCTTCTTTGCAGG-3′) and
- ISAba1_R (5′-AATGATTGGTGACAATGAAG-3′) to amplify ISAba1.
2.5. Calculating the Efficiency of Gene Transfer
2.6. Chromosome vs. Plasmid Location of blaOXA-500
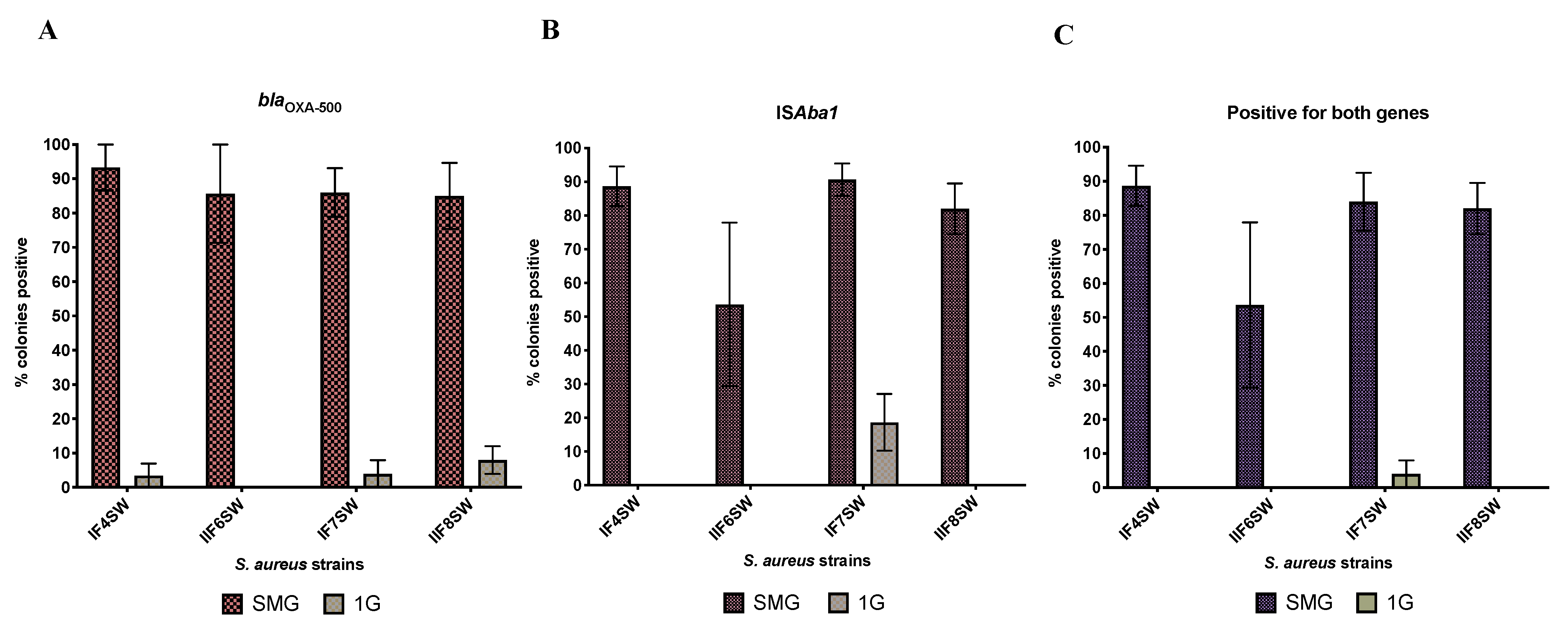
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Schulze-Makuch, D.; Wagner, D.; Kounaves, S.P.; Mangelsdorf, K.; Devine, K.G.; de Vera, J.-P.; Schmitt-Kopplin, P.; Grossart, H.-P.; Parro, V.; Kaupenjohann, M.; et al. Transitory microbial habitat in the hyperarid Atacama Desert. Proc. Natl. Acad. Sci. USA 2018, 115, 2670–2675. [Google Scholar] [CrossRef] [Green Version]
- Carpa, R.; Keul, A.; Muntean, V.; Dobrotă, C. Characterization of halophilic bacterial communities in Turda Salt Mine (Romania). Orig. Life Evol. Biosph. 2014, 44, 223–230. [Google Scholar] [CrossRef] [Green Version]
- Humayoun, S.B.; Bano, N.; Hollibaugh, J.T. Depth distribution of microbial diversity in Mono Lake, a meromictic soda lake in California. Appl. Environ. Microbiol. 2003, 69, 1030–1042. [Google Scholar] [CrossRef] [Green Version]
- Walker, J.J.; Spear, J.R.; Pace, N.R. Geobiology of a microbial endolithic community in the Yellowstone geothermal environment. Nature 2005, 434, 1011–1014. [Google Scholar] [CrossRef] [PubMed]
- Ji, M.; Greening, C.; Vanwonterghem, I.; Carere, C.R.; Bay, S.K.; Steen, J.A.; Montgomery, K.; Lines, T.; Beardall, J.; van Dorst, J.; et al. Atmospheric trace gases support primary production in Antarctic desert surface soil. Nature 2017, 552, 400. [Google Scholar] [CrossRef]
- Yamaguchi, N.; Roberts, M.; Castro, S.; Oubre, C.; Makimura, K.; Leys, N.; Grohmann, E.; Sugita, T.; Ichijo, T.; Nasu, M. Microbial monitoring of crewed habitats in space-current status and future perspectives. Microb. Environ. 2014, 29, 250–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urbaniak, C.; Sielaff, A.C.; Frey, K.G.; Allen, J.E.; Singh, N.; Jaing, C.; Wheeler, K.; Venkateswaran, K. Detection of antimicrobial resistance genes associated with the International Space Station environmental surfaces. Sci. Rep. 2018, 8, 814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Checinska Sielaff, A.; Urbaniak, C.; Mohan, G.B.M.; Stepanov, V.G.; Tran, Q.; Wood, J.M.; Minich, J.; McDonald, D.; Mayer, T.; Knight, R.; et al. Characterization of the total and viable bacterial and fungal communities associated with the International Space Station surfaces. Microbiome 2019, 7, 50. [Google Scholar] [CrossRef] [Green Version]
- Singh, N.K.; Wood, J.M.; Karouia, F.; Venkateswaran, K. Succession and persistence of microbial communities and antimicrobial resistance genes associated with International Space Station environmental surfaces. Microbiome 2018, 6, 204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soucy, S.M.; Huang, J.; Gogarten, J.P. Horizontal gene transfer: Building the web of life. Nat. Rev. Genet. 2015, 16, 472–482. [Google Scholar] [CrossRef]
- Frost, L.S.; Leplae, R.; Summers, A.O.; Toussaint, A. Mobile genetic elements: The agents of open source evolution. Nat. Rev. Microbiol. 2005, 3, 722–732. [Google Scholar] [CrossRef]
- Roberts, A.P.; Kreth, J. The impact of horizontal gene transfer on the adaptive ability of the human oral microbiome. Front. Cell. Infect. Microbiol. 2014, 4, 124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Webster, N.S.; Reusch, T.B.H. Microbial contributions to the persistence of coral reefs. ISME J. 2017, 11, 2167. [Google Scholar] [CrossRef] [PubMed]
- Koonin, E.V.; Makarova, K.S.; Aravind, L. Horizontal gene transfer in prokaryotes: Quantification and classification. Annu. Rev. Microbiol. 2001, 55, 709–742. [Google Scholar] [CrossRef] [PubMed]
- Brooks, A.N.; Turkarslan, S.; Beer, K.D.; Lo, F.Y.; Baliga, N.S. Adaptation of cells to new environments. Wiley Interdiscip. Rev. Syst. Biol. Med. 2011, 3, 544–561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vogan, A.A.; Higgs, P.G. The advantages and disadvantages of horizontal gene transfer and the emergence of the first species. Biol. Direct 2011, 6, 1. [Google Scholar] [CrossRef] [Green Version]
- Schiwon, K.; Arends, K.; Rogowski, K.M.; Fürch, S.; Prescha, K.; Sakinc, T.; Van Houdt, R.; Werner, G.; Grohmann, E. Comparison of antibiotic resistance, biofilm formation and conjugative transfer of Staphylococcus and Enterococcus isolates from International Space Station and Antarctic Research Station Concordia. Microb. Ecol. 2013, 65, 638–651. [Google Scholar] [CrossRef]
- Evans, B.A.; Amyes, S.G.B. OXA β-lactamases. Clin. Microbiol. Rev. 2014, 27, 241–263. [Google Scholar] [CrossRef] [Green Version]
- Khurshid, M.; Rasool, M.H.; Ashfaq, U.A.; Aslam, B.; Waseem, M. Emergence of ISAba1 harboring carbapenem-resistant Acinetobacter baumannii isolates in Pakistan. Future Microbiol. 2017, 12, 1261–1269. [Google Scholar] [CrossRef]
- Mugnier, P.D.; Poirel, L.; Nordmann, P. Functional analysis of insertion sequence ISAba1, responsible for genomic plasticity of Acinetobacter baumannii. J. Bacteriol. 2009, 191, 2414–2418. [Google Scholar] [CrossRef] [Green Version]
- Checinska Sielaff, A.; Singh, N.K.; Allen, J.E.; Thissen, J.; Jaing, C.; Venkateswaran, K. Draft Genome Sequences of Biosafety Level 2 Opportunistic Pathogens Isolated from the Environmental Surfaces of the International Space Station. Genome Announc. 2016, 4, e01263–e01316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brinkac, L.; Voorhies, A.; Gomez, A.; Nelson, K.E. The Threat of Antimicrobial Resistance on the Human Microbiome. Microb. Ecol. 2017, 74, 1001–1008. [Google Scholar] [CrossRef] [PubMed]
- Penders, J.; Stobberingh, E.E.; Savelkoul, P.H.M.; Wolffs, P.F.G. The human microbiome as a reservoir of antimicrobial resistance. Front. Microbiol. 2013, 4, 87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huddleston, J.R. Horizontal gene transfer in the human gastrointestinal tract: Potential spread of antibiotic resistance genes. Infect. Drug Resist. 2014, 7, 167–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wielders, C.L.C.; Vriens, M.R.; Brisse, S.; de Graaf-Miltenburg, L.A.M.; Troelstra, A.; Fleer, A.; Schmitz, F.J.; Verhoef, J.; Fluit, A.C. Evidence for in-vivo transfer of mecA DNA between strains of Staphylococcus aureus. Lancet 2001, 357, 1674–1675. [Google Scholar] [CrossRef]
- Charpentier, X.; Kay, E.; Schneider, D.; Shuman, H.A. Antibiotics and UV Radiation Induce Competence for Natural Transformation in Legionella pneumophila. J. Bacteriol. 2011, 193, 1114–1121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Claverys, J.-P.; Prudhomme, M.; Martin, B. Induction of Competence Regulons as a General Response to Stress in Gram-Positive Bacteria. Annu. Rev. Microbiol. 2006, 60, 451–475. [Google Scholar] [CrossRef]
- Manna, S.; Waring, A.; Papanicolaou, A.; Hall, N.E.; Bozinovski, S.; Dunne, E.M.; Satzke, C. The transcriptomic response of Streptococcus pneumoniae following exposure to cigarette smoke extract. Sci. Rep. 2018, 8, 15716. [Google Scholar] [CrossRef] [Green Version]
- Slager, J.; Kjos, M.; Attaiech, L.; Veening, J.W. Antibiotic-induced replication stress triggers bacterial competence by increasing gene dosage near the origin. Cell 2014, 157, 395–406. [Google Scholar] [CrossRef] [Green Version]
- Fagerlund, A.; Granum, P.E.; Håvarstein, L.S. Staphylococcus aureus competence genes: Mapping of the SigH, ComK1 and ComK2 regulons by transcriptome sequencing. Mol. Microbiol. 2014, 94, 557–579. [Google Scholar] [CrossRef]
- Morikawa, K.; Takemura, A.J.; Inose, Y.; Tsai, M.; Nguyen Thi le, T.; Ohta, T.; Msadek, T. Expression of a cryptic secondary sigma factor gene unveils natural competence for DNA transformation in Staphylococcus aureus. PLoS Pathog. 2012, 8, e1003003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, K.L.; Roberts, C.; Disz, T.; Vonstein, V.; Hwang, K.; Overbeek, R.; Olson, P.D.; Projan, S.J.; Dunman, P.M. Characterization of the Staphylococcus aureus heat shock, cold shock, stringent, and SOS responses and their effects on log-phase mRNA turnover. J. Bacteriol. 2006, 188, 6739–6756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engelmoer, D.J.; Rozen, D.E. Competence increases survival during stress in Streptococcus pneumoniae. Evol. Int. J. Org. Evol. 2011, 65, 3475–3485. [Google Scholar] [CrossRef] [PubMed]
- Schultz, D.; Wolynes, P.G.; Jacob, E.B.; Onuchic, J.N. Deciding fate in adverse times: Sporulation and competence in Bacillus subtilis. Proc. Natl. Acad. Sci. USA 2009, 106, 21027–21034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Kim, Y.; Ma, Q.; Hong, S.H.; Pokusaeva, K.; Sturino, J.M.; Wood, T.K. Cryptic prophages help bacteria cope with adverse environments. Nat. Commun. 2010, 1, 147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du Toit, A. Phage induction in different contexts. Nat. Rev. Microbiol. 2019, 17, 126–127. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| S. aureus Recipient Strain | SMG (Range) | 1G (Range) |
|---|---|---|
| IF4SW | 4.21 × 10−7 (1.1 × 10−6 to 5.2 × 10−8) | 2.05 × 10−9 (0.0 to 4.1 × 10−9) |
| IIF6SW | 4.54 × 10−8 (7.5 × 10−8 to 6.3 × 10−9) | 0.0 (no transfer) |
| IF7SW | 3.20 × 10−7 (7.2 × 10−7 to 9.9 × 10−8) | 4.33 × 10−9 (7.5 × 10−9 to 2.2 × 10−9) |
| IIF8SW | 4.38 × 10−7 (6.8 × 10−7 to 3.1 × 10−9) | 6.07 × 10−9 (0.0 to 1.2 × 10−9) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Urbaniak, C.; Grams, T.; Mason, C.E.; Venkateswaran, K. Simulated Microgravity Promotes Horizontal Gene Transfer of Antimicrobial Resistance Genes between Bacterial Genera in the Absence of Antibiotic Selective Pressure. Life 2021, 11, 960. https://doi.org/10.3390/life11090960
Urbaniak C, Grams T, Mason CE, Venkateswaran K. Simulated Microgravity Promotes Horizontal Gene Transfer of Antimicrobial Resistance Genes between Bacterial Genera in the Absence of Antibiotic Selective Pressure. Life. 2021; 11(9):960. https://doi.org/10.3390/life11090960
Chicago/Turabian StyleUrbaniak, Camilla, Tristan Grams, Christopher E. Mason, and Kasthuri Venkateswaran. 2021. "Simulated Microgravity Promotes Horizontal Gene Transfer of Antimicrobial Resistance Genes between Bacterial Genera in the Absence of Antibiotic Selective Pressure" Life 11, no. 9: 960. https://doi.org/10.3390/life11090960
APA StyleUrbaniak, C., Grams, T., Mason, C. E., & Venkateswaran, K. (2021). Simulated Microgravity Promotes Horizontal Gene Transfer of Antimicrobial Resistance Genes between Bacterial Genera in the Absence of Antibiotic Selective Pressure. Life, 11(9), 960. https://doi.org/10.3390/life11090960