
Salinity Stress in Potato: Understanding Physiological, Biochemical and Molecular Responses

 , , , ,
, , , ,  ,
, 
Abstract
1. Introduction
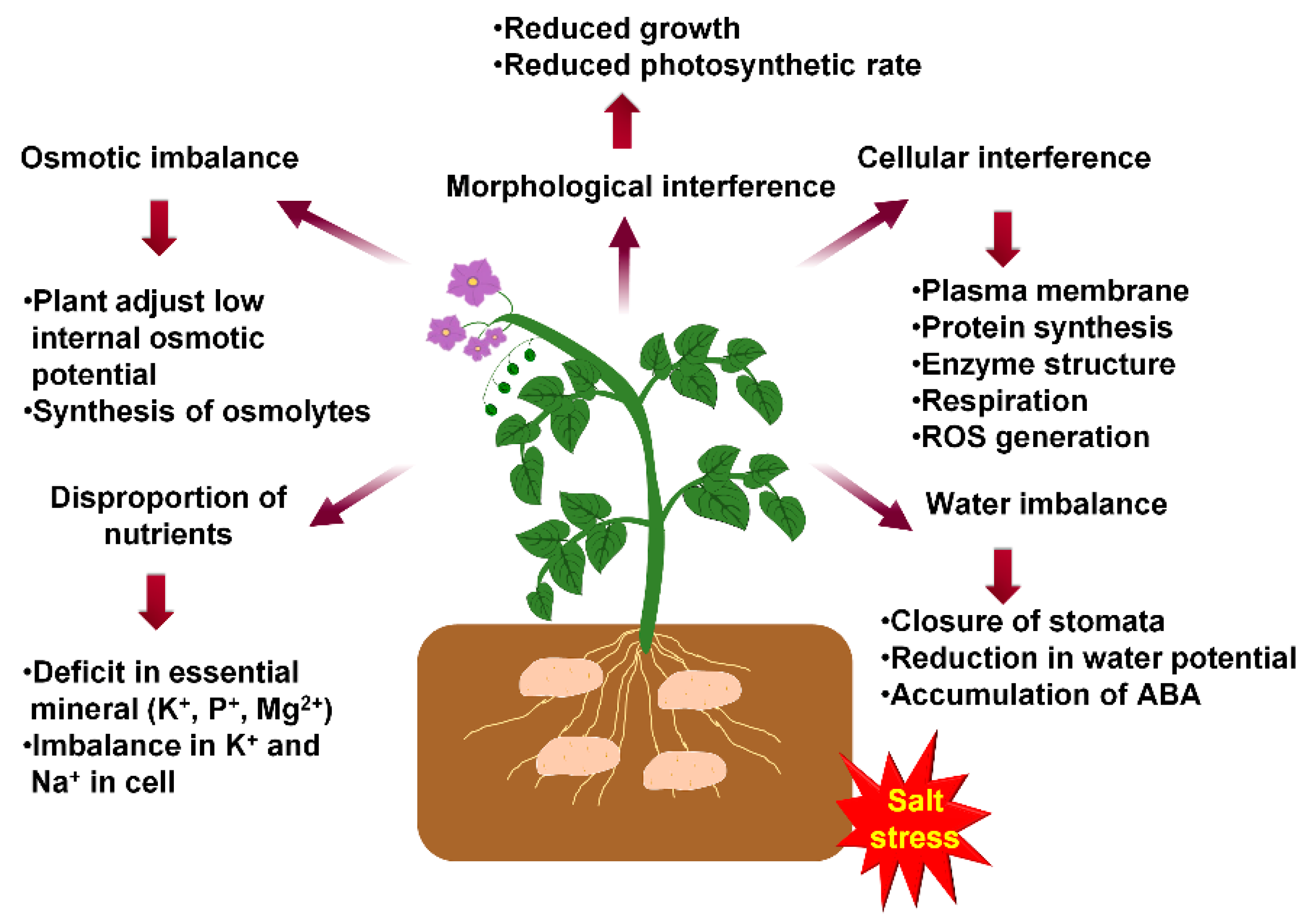
2. Effects of Salt Stress on Potato Growth and Development
3. Effect of Salinity on the Physiology of Potato
3.1. Effect of Salinity on Source-Sink Relationship
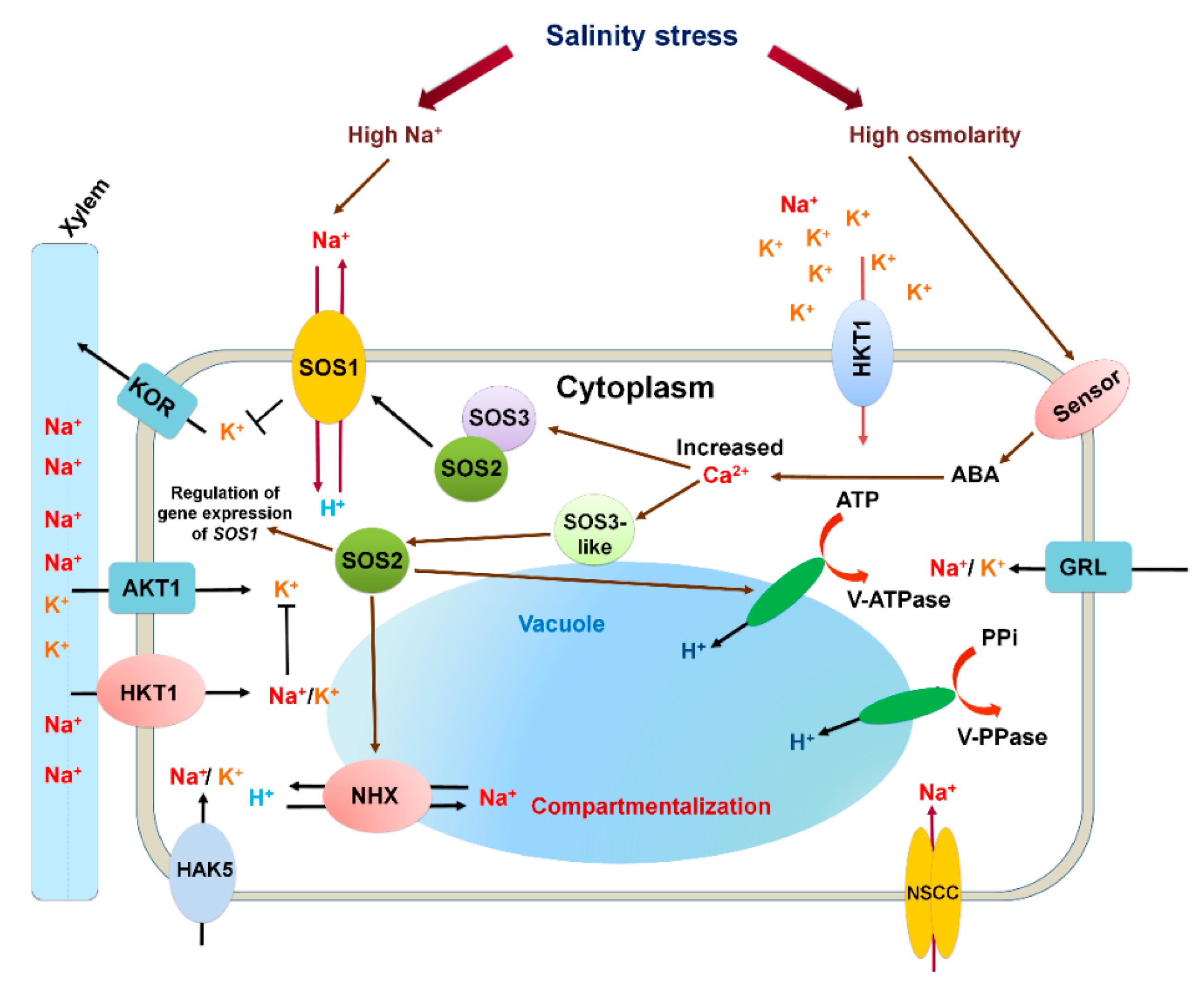
3.2. Ion Homeostasis
3.3. Hormonal Effect
3.4. Water Status in Plants
3.5. Anatomical Changes and Ultrastructural Changes
3.6. Photosynthesis
4. Effect of Salinity on Biochemical Traits of Potato
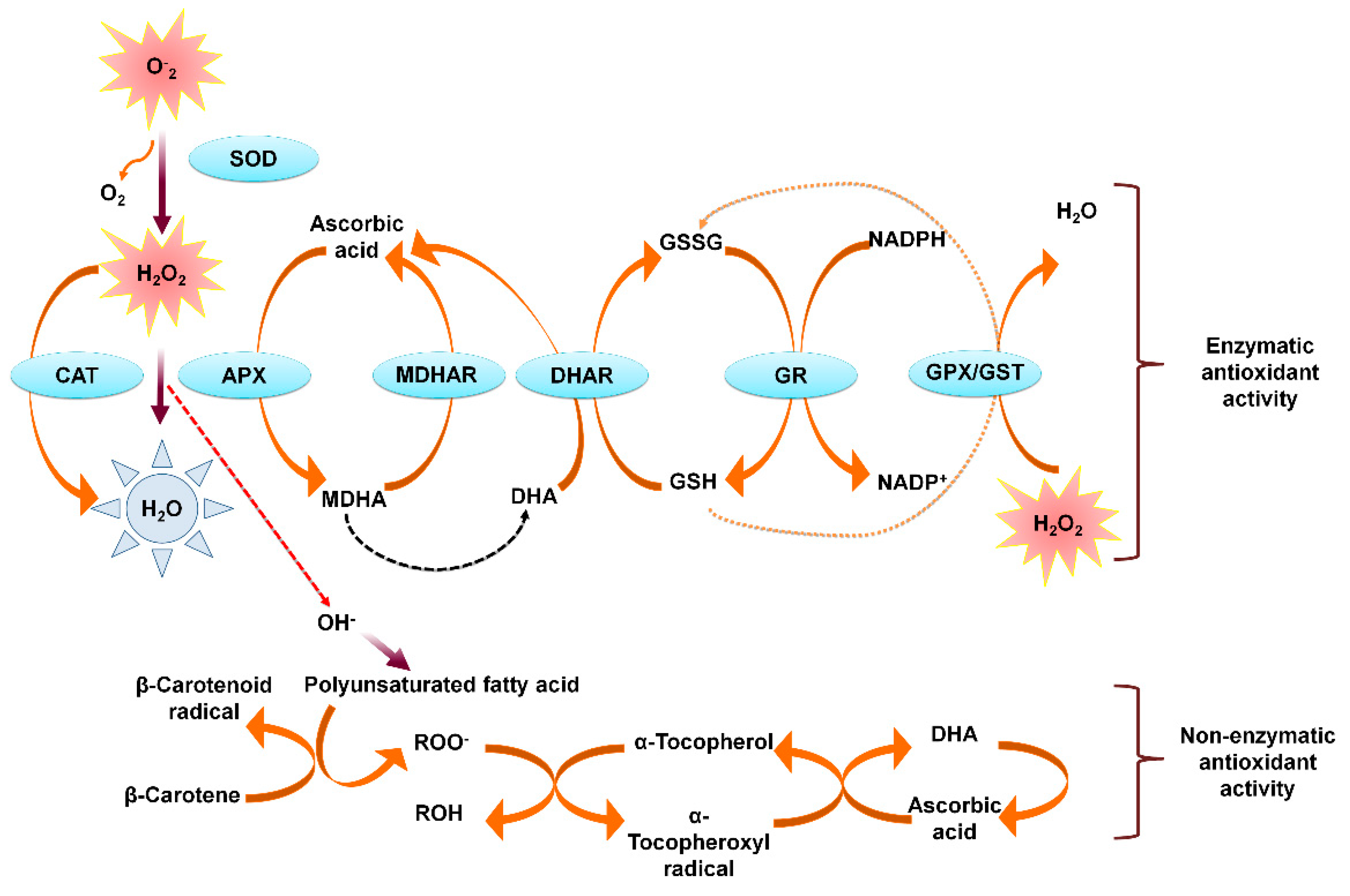
4.1. ROS and Antioxidant System
4.2. Osmolyte Function
4.3. Proline
4.4. Polyols
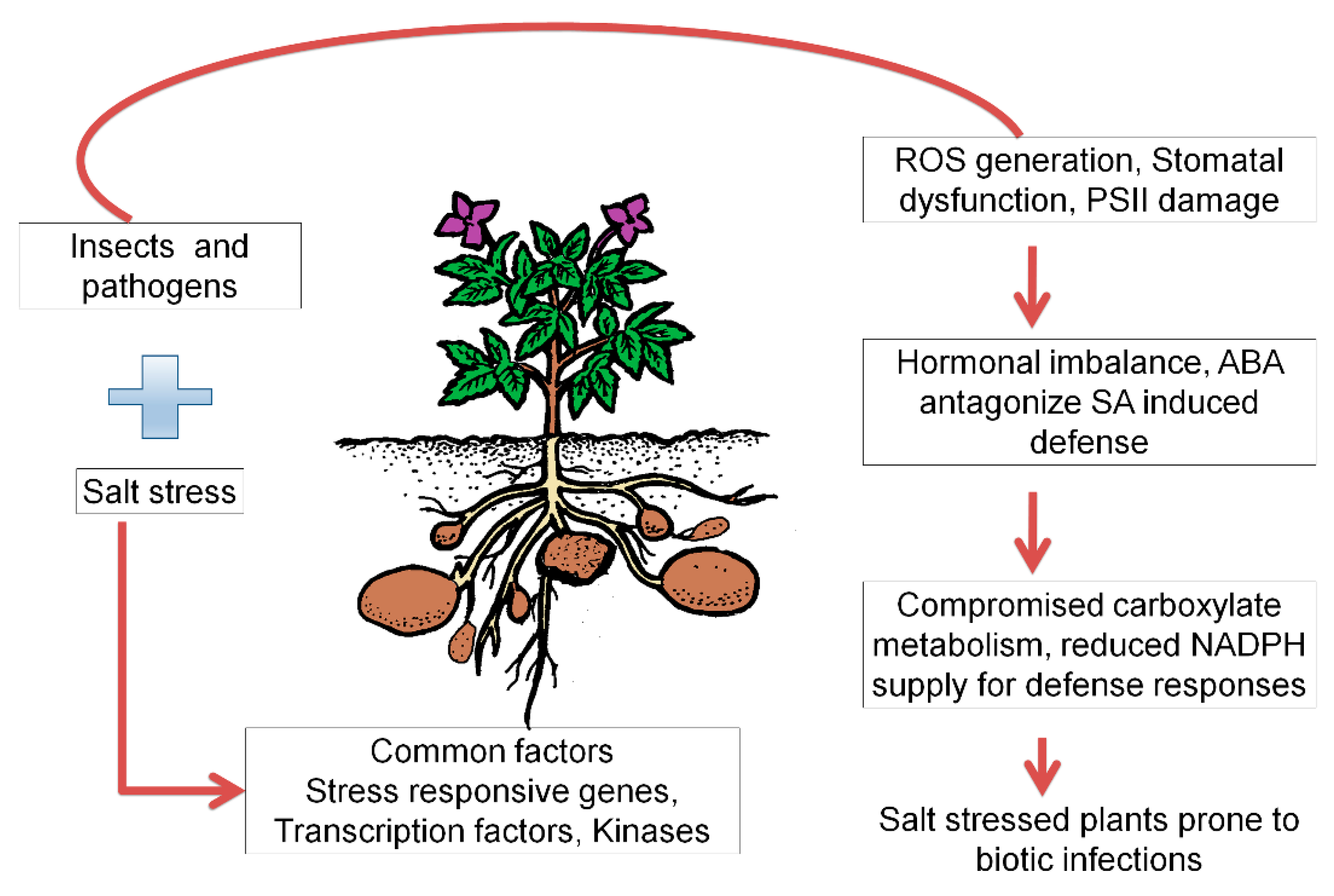
5. Disease and Pest Susceptibility
6. Improving Salt Tolerance in Potato
6.1. Wild Relatives and Primitive Potato Species
6.2. Breeding Approach for Enhancing Salt Tolerance
6.3. Genetic Engineering Approach to Improve Salt Tolerance in Potato
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lutaladio, N.; Castaldi, L. Potato: The Hidden Treasure. J. Food Compost. Anal. 2009, 22, 491–493. [Google Scholar] [CrossRef]
- Raigond, P.; Atkinson, F.S.; Lal, M.K.; Thakur, N.; Singh, B.; Mishra, T. Potato Carbohydrates. In Potato; Raigond, P., Singh, B., Dutt, S., Chakrabarti, S., Eds.; Springer: Singapore, 2020; pp. 13–36. [Google Scholar]
- Changan, S.S.; Taylor, M.A.; Raigond, P.; Dutt, S.; Kumar, D.; Lal, M.K.; Kumar, M.; Tomar, M.; Singh, B. Potato Carotenoids. In Potato; Raigond, P., Singh, B., Dutt, S., Chakrabarti, S., Eds.; Springer: Singapore, 2020; pp. 151–171. [Google Scholar]
- Lal, M.K.; Kumar, A.; Jena, R.; Dutt, S.; Thakur, N.; Parmar, V.; Kumar, V.; Singh, B. Lipids in Potato. In Potato; Raigond, P., Singh, B., Dutt, S., Chakrabarti, S., Eds.; Springer: Singapore, 2020; pp. 73–85. [Google Scholar]
- Lal, M.K.; Kumar, A.A.; Kumar, A.A.; Raigond, P.; Oko, A.O.; Thakur, N.; Parmar, V.; Thakur, A.; Singh, B. Dietary Fibres in Potato. In Potato; Raigond, P., Singh, B., Dutt, S., Chakrabarti, S., Eds.; Springer: Singapore, 2020; pp. 37–50. [Google Scholar]
- Cleland, J. World Population Growth; Past, Present and Future. Environ. Resour. Econ. 2013, 55, 543–554. [Google Scholar] [CrossRef]
- Devaux, A.; Kromann, P.; Ortiz, O. Potatoes for Sustainable Global Food Security. Potato Res. 2014, 57, 185–199. [Google Scholar] [CrossRef]
- Hijmans, R.J. The Effect of Climate Change on Global Potato Production. Am. J. Potato Res. 2003, 80, 271–279. [Google Scholar] [CrossRef]
- Tiwari, R.K.; Lal, M.K.; Naga, K.C.; Kumar, R.; Chourasia, K.N.; Subhash, S.; Kumar, D.; Sharma, S.U. Emerging roles of melatonin in mitigating abiotic and biotic stresses of horticultural crops. Sci. Hortic. 2020, 272, 109592. [Google Scholar] [CrossRef]
- Levy, D.; Veilleux, R.E. Adaptation of Potato to High Temperatures and Salinity—A review. Am. J. Potato Res. 2007, 84, 487–506. [Google Scholar] [CrossRef]
- Qadir, M.; Quillérou, E.; Nangia, V.; Murtaza, G.; Singh, M.; Thomas, R.J.; Drechsel, P.; Noble, A.D. Economics of Salt-induced Land Degradation and Restoration. Nat. Resour. Forum. 2014, 38, 282–295. [Google Scholar] [CrossRef]
- Eliazer Nelson, A.R.L.; Ravichandran, K.; Antony, U. The Impact of the Green Revolution on Indigenous Crops of India. J. Ethn. Foods 2019, 6, 8. [Google Scholar] [CrossRef]
- Sharma, D.K.; Singh, A. Salinity Research in India-achievements, Challenges and Future Prospects. Water Energy Int. 2015, 58, 35–45. [Google Scholar]
- Neumann, P. Salinity Resistance and Plant Growth Revisited. Plant Cell Environ. 1997, 20, 1193–1198. [Google Scholar] [CrossRef]
- Shao, H.; Chu, L.; Lu, H.; Qi, W.; Chen, X.; Liu, J.; Kuang, S.; Tang, B.; Wong, V. Towards sustainable agriculture for the salt-affected soil. Land Degrad. Dev. 2019, 30, 574–579. [Google Scholar] [CrossRef]
- Shrivastava, P.; Kumar, R. Soil salinity: A Serious Environmental Issue and Plant Growth Promoting Bacteria as one of the Tools for its Alleviation. Saudi J. Biol. Sci. 2015, 22, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Rengasamy, P. World salinization with emphasis on Australia. J. Exp. Bot. 2006, 57, 1017–1023. [Google Scholar] [CrossRef] [PubMed]
- Ahanger, M.A.; Tomar, N.S.; Tittal, M.; Argal, S.; Agarwal, R.M. Plant Growth under Water/Salt Stress: ROS Production; Antioxidants and Significance of added Potassium under such Conditions. Physiol. Mol. Biol. Plants 2017, 23, 731–744. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, R.K.; Kumar, R.; Sharma, S.; Sagar, V.; Aggarwal, R.; Naga, K.C.; Lal, M.K.; Chourasia, K.N.; Kumar, D.; Kumar, M. Potato Dry Rot disease: Current Status, Pathogenomics and Management. 3 Biotech 2020, 10, 503. [Google Scholar] [CrossRef]
- Babu, M.A.; Singh, D.; Gothandam, K.M. The Effect of Salinity on Growth, Hormones and Mineral Elements in Leaf and Fruit of Tomato Cultivar PKM1. J. Anim. Plant Sci. 2012, 22, 159–164. [Google Scholar]
- Heuer, B.; Nadler, A. Growth and Development of Potatoes under Salinity and Water deficit. Aust. J. Agric. Res. 1995, 46, 1477–1486. [Google Scholar] [CrossRef]
- Jha, G.; Choudhary, O.P.; Sharda, R. Comparative effects of saline water on yield and quality of potato under drip and furrow irrigation. Cogent Food Agric. 2017, 3, 369345. [Google Scholar] [CrossRef]
- Ma, Y.; Dias, M.C.; Freitas, H. Drought and Salinity Stress Responses and Microbe-Induced Tolerance in Plants. Front. Plant Sci. 2020, 11, 1750. [Google Scholar] [CrossRef]
- Zhang, Z.; Mao, B.; Li, H.; Zhou, W.; Takeuchi, Y.; Yoneyama, K. Effect of Salinity on Physiological Characteristics, Yield and Quality of Microtubers in vitro in Potato. Acta Physiol. Plant 2005, 27, 481–489. [Google Scholar] [CrossRef]
- Machado, R.M.A.; Serralheiro, R.P. Soil salinity: Effect on Vegetable Crop Growth. Management Practices to Prevent and Mitigate Soil Salinization. Horticulturae 2017, 3, 30. [Google Scholar] [CrossRef]
- Blom-Zandstra, M.; Wolters, W.; Heinen, M.; Roest, C.W.J.; Smit, R.W.; Smit, A.L. Perspectives for the Growth of Salt tolerant cash Crops: A Case Study with Potato. Plant Res. Int. Bus. Unit Agrosystems Res. 2014, 572, 36. [Google Scholar]
- Tavakkoli, E.; Rengasamy, P.; McDonald, G.K. High Concentrations of Na+ and Cl- ions in Soil Solution have Simultaneous Detrimental effects on Growth of Faba bean under Salinity Stress. J. Exp. Bot. 2010, 61, 4449–4459. [Google Scholar] [CrossRef] [PubMed]
- Hirasawa, T.; Sato, K.; Yamaguchi, M.; Narita, R.; Kodama, A.; Adachi, S.; Ookawa, T.; Sato, K. Differences in Dry matter Production, Grain production, and Photosynthetic Rate in Barley cultivars under Long-term Salinity. Plant Prod. Sci. 2017, 20, 288–299. [Google Scholar] [CrossRef]
- Dahal, K.; Li, X.-Q.; Tai, H.; Creelman, A.; Bizimungu, B. Improving Potato Stress Tolerance and Tuber Yield Under a Climate Change Scenario—A Current Overview. Front. Plant Sci. 2019, 10, 563. [Google Scholar] [CrossRef]
- Chakraborty, K.; Mondal, S.; Ray, S.; Samal, P.; Pradhan, B.; Chattopadhyay, K.; Kar, M.K.; Swain, P.; Sarkar, R.K. Tissue Tol-erance Coupled with Ionic Discrimination Can Potentially Minimize the Energy Cost of Salinity Tolerance in Rice. Front. Plant Sci. 2020, 11, 265. [Google Scholar] [CrossRef]
- Roy, S.J.; Negrão, S.; Tester, M. Salt Resistant Crop Plants. Curr. Opin. Biotechnol. 2014, 26, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Bündig, C.; Jozefowicz, A.M.; Mock, H.-P.; Winkelmann, T. Proteomic Analysis of two Divergently Responding Potato Genotypes (Solanum tuberosum L.) following Osmotic Stress Treatment in vitro. J. Proteomics 2016, 143, 227–241. [Google Scholar] [CrossRef]
- Sarker, U.; Oba, S. Drought Stress Effects on Growth, ROS Markers, Compatible Solutes, Phenolics, Flavonoids, and Antioxidant Activity in Amaranthus tricolor. Appl. Biochem. Biotechnol. 2018, 186, 999–1016. [Google Scholar] [CrossRef]
- Sarker, U.; Oba, S. Catalase, Superoxide dismutase and Ascorbate-glutathione Cycle Enzymes confer Drought Tolerance of Amaranthus tricolor. Sci. Rep. 2018, 8, 1–12. [Google Scholar] [CrossRef]
- Ödemiş, B.; Çalişkan, M.E. Photosynthetic Response of Potato Plants to Soil Salinity. Türk Tarım ve Doğa Bilim. Dergisi-Turkish J. Agric. Nat. Sci. 2014, 1, 1429–1439. [Google Scholar]
- Katerji, N.; van Hoorn, J.W.; Hamdy, A.; Mastrorilli, M. Salinity Effect on Crop Development and Yield, Analysis of Salt tolerance according to several Classification Methods. Agric. Water Manag. 2003, 62, 37–66. [Google Scholar] [CrossRef]
- Londhe, S. Cultivation of Potato on Abiotic Stress-affected Soils of India. Curr. Sci. 2016, 111, 21–22. [Google Scholar] [CrossRef]
- Flowers, T.J.; Flowers, S.A. Why Does Salinity Pose such a difficult Problem for Plant Breeders? Agric. Water Manag. 2005, 78, 15–24. [Google Scholar] [CrossRef]
- Ashraf, M.; Foolad, M.R. Crop Breeding for Salt Tolerance in the era of Molecular Markers and Marker-assisted Selection. Plant Breed. 2013, 132, 10–20. [Google Scholar] [CrossRef]
- Velásquez, B.; Balzarini, M.; Taleisnik, E. Salt Tolerance Variability amongst Argentine Andean Potatoes (Solanum tuberosum L. subsp. andigena). Potato Res. 2005, 48, 59–67. [Google Scholar] [CrossRef]
- Naik, P.S.; Widholm, J.M. Comparison of Tissue culture and Whole plant Responses to Salinity in Potato. Plant Cell. Tissue Organ Cult. 1993, 33, 273–280. [Google Scholar] [CrossRef]
- Alhagdow, M.M.; Barthakur, N.N.; Donnelly, D.J. Salinity Stress and Sodium-potassium interactions in Micropropagated Potatoes. Potato Res. 1999, 42, 73–78. [Google Scholar] [CrossRef]
- Ghosh, S.C.; Asanuma, K.I.; Kusutani, A.; Toyota, M. Effect of Salt Stress on some Chemical Components and Yield of Potato. Soil Sci. Plant Nutr. 2001, 47, 467–475. [Google Scholar] [CrossRef]
- Jbir-Koubaa, R.; Charfeddine, S.; Ellouz, W.; Saidi, M.N.; Drira, N.; Gargouri-Bouzid, R.; Nouri-Ellouz, O. Investigation of the Response to Salinity and to Oxidative Stress of Interspecific Potato Somatic hybrids grown in a Greenhouse. Plant Cell. Tissue Organ Cult. 2015, 120, 933–947. [Google Scholar] [CrossRef]
- Hossain, M.; Kawochar, A.; Rahaman, E. Screening of CIP Potato clones for Salinity Tolerance in Pot and Field Condition. Adv. Plants Agric. Res. 2018, 8, 573–580. [Google Scholar] [CrossRef]
- Levy, D. The Response of Potatoes (Solunum tuberosum L.) to Salinity: Plant growth and Tuber yields in the arid Desert of Israel. Ann. Appl. Biol. 1992, 120, 547–555. [Google Scholar] [CrossRef]
- Kumar, D.; Ezekiel, R. Developmental Changes in Sugars and Dry matter content of Potato Tuber under Sub-tropical Climates. Sci. Hortic. 2006, 110, 129–134. [Google Scholar] [CrossRef]
- Dobránszki, J.; Magyar-Tábori, K.; Hudák, I. In vitro Tuberization in Hormone-free systems on Solidified Medium and Dormancy of Potato Microtubers. Fruit Veg. Cereal Sci. Biotechnol. 2008, 2, 82–94. [Google Scholar]
- Richardson, K.V.A.; Wetten, A.C.; Caligari, P.D.S. Cell and Nuclear Degradation in Root meristems following exposure of Potatoes (Solarium tuberosum L.) to salinity. Potato Res. 2001, 44, 389–399. [Google Scholar] [CrossRef]
- Murshed, R.; Najla, S.; Albiski, F.; Kassem, I.; Jbour, M.; Al-Said, H. Using Growth Parameters for in-vitro Screening of Potato varieties Tolerant to Salt stress. J. Agric. Sci. Technol. 2015, 17, 483–494. [Google Scholar]
- Faried, H.N.; Ayyub, C.M.; Amjad, M.; Ahmed, R. Salinity impairs Ionic, Physiological and Biochemical attributes in Potato. Pakistan J. Agric. Sci. 2016, 53, 17–25. [Google Scholar] [CrossRef]
- Acosta-Motos, J.R.; Ortuño, M.F.; Bernal-Vicente, A.; Diaz-Vivancos, P.; Sanchez-Blanco, M.J.; Hernandez, J.A. Plant responses to salt stress: Adaptive mechanisms. Agronomy 2017, 7, 18. [Google Scholar] [CrossRef]
- Tiwari, R.K.; Lal, M.K.; Kumar, R.; Chourasia, K.N.; Naga, K.C.; Kumar, D.; Das, S.K.; Zinta, G. Mechanistic insights on melatonin-mediated drought stress mitigation in plants. Physiol. Plant 2020. [Google Scholar] [CrossRef]
- Levy, D.; Fogelman, E.; Itzhak, Y. The Effect of Water Salinity on Potatoes (Solanum tuberosum L.): Physiological indices and Yielding Capacity. Potato Res. 1988, 31, 601–610. [Google Scholar] [CrossRef]
- Sarker, U.; Oba, S. The Response of Salinity Stress-Induced A. tricolor to Growth, Anatomy, Physiology, Non-Enzymatic and Enzymatic Antioxidants. Front. Plant Sci. 2020, 11, 1–14. [Google Scholar] [CrossRef]
- Keisham, M.; Mukherjee, S.; Bhatla, S.C. Mechanisms of Sodium Transport in Plants-Progresses and Challenges. Int. J. Mol. Sci. 2018, 19, 647. [Google Scholar] [CrossRef]
- Colmer, T.D.; Epstein, E.; Dvorak, J. Differential Solute Regulation in Leaf Blades of Various Ages in Salt Sensitive Wheat and Salt Tolerant Wheat×Lophopyrum elongatum (Host) Love Lamphiploid. Plant Physiol. 1995, 108, 1715–1724. [Google Scholar] [CrossRef]
- Santa-Maria, G.E.; Epstein, E. Potassium/Sodium Selectivity in Wheat and Amphiploid Cross Wheat×Lophopyrum elongatum. Plant Sci. 2001, 160, 523–534. [Google Scholar] [CrossRef]
- Flowers, T.J.; Yeo, A.R. Ion Relations of Plants under Drought and Salinity. Aust. J. Plant Physiol. 1986, 13, 75–91. [Google Scholar] [CrossRef]
- Chindi, A.; Woldegiorgis, G.; Solomon, A.; Tesema, L.; Negash, K.; Lemaga, B.; Schulz, S. Enhancing Potato Seed Production using Rapid Multiplication Techniques. In Proceedings of the National Workshop on Seed Potato Tuber Production and Dissemination, Bahir Dar, Ethiopia, 12–14 March 2012; Ethiopian Institute of Agricultural Research (EIAR): Addis Ababa, Ethiopia; Amhara Regional Agricultural Research Institute (ARARI): Bahir Dar, Ethiopia, 2013; pp. 91–100. [Google Scholar]
- Lal, M.K.; Kumar, A.; Raigond, P.; Dutt, S.; Changan, S.S.; Chourasia, K.N.; Tiwari, R.K.; Kumar, D.; Sharma, S.; Chakrabarti, S.K.; et al. Impact of Starch Storage Condition on Glycemic Index and Resistant Starch of Cooked Potato (Solanum tuberosum) Tubers. Starch Stärke 2020, 73, 1900281. [Google Scholar] [CrossRef]
- Zaman, M.; Mannan, M.; Samanta, S. Salt Induced Changes in Dry Matter Accumulation and Yield of Mustard (Brassica juncea). Bangladesh Agron. J. 2015, 18, 65–70. [Google Scholar] [CrossRef]
- Barbieri, G.; Vallone, S.; Orsini, F.; Paradiso, R.; De Pascale, S.; Negre-Zakharov, F.; Maggio, A. Stomatal density and Metabolic Determinants mediate Salt Stress Adaptation and Water use efficiency in Basil (Ocimum basilicum L.). J. Plant Physiol. 2012, 169, 1737–1746. [Google Scholar] [CrossRef] [PubMed]
- Gholizadeh, A.; Saberioon, M.; Borůvka, L.; Wayayok, A.; Mohd Soom, M.A. Leaf chlorophyll and Nitrogen Dynamics and their Relationship to Lowland Rice yield for Site-specific Paddy Management. Inf. Process. Agric. 2017, 4, 259–268. [Google Scholar] [CrossRef]
- Munns, R.; Tester, M. Mechanisms of Salinity Tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [PubMed]
- Gupta, B.; Huang, B. Mechanism of Salinity Tolerance in Plants: Physiological, Biochemical, and Molecular Characterization. Int. J. Genom. 2014, 2014, 701596. [Google Scholar] [CrossRef] [PubMed]
- Sarker, U.; Oba, S. Response of Nutrients, Minerals, Antioxidant Leaf Pigments, Vitamins, Polyphenol, Flavonoid and Antioxidant Activity in Selected Vegetable Amaranth under four Soil Water Content. Food Chem. 2018, 252, 72–83. [Google Scholar] [CrossRef]
- Sarker, U.; Oba, S. Drought stress enhances nutritional and bioactive compounds, phenolic acids and antioxidant capacity of Amaranthus leafy vegetable. BMC Plant Biol. 2018, 18, 1–15. [Google Scholar] [CrossRef]
- Albacete, A.A.; Martínez-Andújar, C.; Pérez-Alfocea, F. Hormonal and Metabolic Regulation of Source-sink Relations Under Salinity and Drought: From Plant Survival to Crop Yield Stability. Biotechnol. Adv. 2014, 32, 12–30. [Google Scholar] [CrossRef]
- Suwa, R.; Nguyen, N.T.; Saneoka, H.; Moghaieb, R.; Fujita, K. Effect of Salinity Stress on Photosynthesis and Vegetative Sink in Tobacco Plants. Soil Sci. Plant Nutr. 2006, 52, 243–250. [Google Scholar] [CrossRef]
- Aliche, E.B.; Theeuwen, T.P.J.M.; Oortwijn, M.; Visser, R.G.F.; van der Linden, C.G. Carbon Partitioning Mechanisms in Potato under Drought Stress. Plant Physiol. Biochem. 2020, 146, 211–219. [Google Scholar] [CrossRef] [PubMed]
- Rahnama, H.; Vakilian, H.; Fahimi, H.; Ghareyazie, B. Enhanced Salt Stress Tolerance in Transgenic Potato Plants (Solanum tuberosum L.) expressing a bacterial mtlD gene. Acta Physiol. Plant 2011, 33, 1521–1532. [Google Scholar] [CrossRef]
- Ishikawa, T.; Shabala, S. Control of Xylem Na+ loading and Transport to the Shoot in Rice and Barley as a Determinant of differential Salinity Stress Tolerance. Physiol. Plant 2019, 165, 619–631. [Google Scholar] [CrossRef] [PubMed]
- Sahoo, M.R.; Devi, T.R.; Dasgupta, M.; Nongdam, P.; Prakash, N. Reactive Oxygen Species Scavenging Mechanisms Associated with Polyethylene glycol mediated Osmotic Stress Tolerance in Chinese Potato. Sci. Rep. 2020, 10, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Si, H.J.; Wen, G.; Du, H.H.; Liu, B.L.; Wang, D. Enhanced Drought and Salinity Tolerance in Transgenic Potato Plants with a BADH gene from Spinach. Plant Biotechnol. Rep. 2011, 5, 71–77. [Google Scholar] [CrossRef]
- Jaarsma, R.; de Boer, A.H. Salinity Tolerance of two Potato Cultivars (Solanum tuberosum) correlates with differences in Vacuolar Transport Activity. Front. Plant Sci. 2018, 9, 737. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Liu, Y.; Li, D.; Feng, S.; Yang, J.; Zhang, J.; Wang, D.; Gan, Y. Improving Salt Tolerance in Potato through Overexpression of AtHKT1 gene. BMC Plant Biol. 2019, 19, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Shabala, S.; Mackay, A. Ion Transport in Halophytes; Academic Press: Cambridge, MA, USA, 2011; Volume 57. [Google Scholar]
- Rodriguez-Rosales, M.P.; Jiang, X.; Gálvez, F.J.; Aranda, M.N.; Cubero, B.; Venema, K. Overexpression of the Tomato K+/H+ antiporter LeNHX2 confers Salt Tolerance by improving Potassium Compartmentalization. New Phytol. 2008, 179, 366–377. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Qin, Y.; Hu, X.; Li, G.; Ding, H.; Xiong, X.; Wang, W. Transcriptome analysis Uncovers the Gene expression Profile of Salt-stressed Potato (Solanum tuberosum L.). Sci. Rep. 2020, 10, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Volkov, V. Salinity tolerance in plants. Quantitative Approach to Ion transport starting from Halophytes and Stepping to Genetic and Protein Engineering for Manipulating Ion fluxes. Front. Plant Sci. 2015, 6, 873. [Google Scholar] [CrossRef] [PubMed]
- Maathuis, F.J.M.; Ahmad, I.; Patishtan, J. Regulation of Na+ fluxes in Plants. Front. Plant Sci. 2014, 5, 467. [Google Scholar] [CrossRef]
- Etehadnia, M.; Waterer, D.; De Jong, H.; Tanino, K.K. Scion and Rootstock effects on ABA-mediated Plant Growth Regulation and Salt Tolerance of acclimated and unacclimated Potato Genotypes. J. Plant Growth Regul. 2008, 27, 125–140. [Google Scholar] [CrossRef]
- Zhu, J.K. Abiotic Stress Signaling and Responses in Plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef]
- Wang, Z.; Hong, Y.; Zhu, G.; Li, Y.; Niu, Q.; Yao, J.; Hua, K.; Bai, J.; Zhu, Y.; Shi, H.; et al. Loss of Salt Tolerance during Tomato Domestication Conferred by Variation in a Na+ /K+ transporter. EMBO J. 2020, 1–14. [Google Scholar] [CrossRef]
- Bouaziz, D.; Jbir, R.; Charfeddine, S.; Saidi, M.N.; Gargouri-Bouzid, R. The StDREB1 transcription factor is involved in Oxidative Stress Response and Enhances Tolerance to Salt stress. Plant Cell. Tissue Organ Cult. 2015, 121, 237–248. [Google Scholar] [CrossRef]
- Suzuki, N.; Rivero, R.M.; Shulaev, V.; Blumwald, E.; Mittler, R. Abiotic and Biotic Stress Combinations. New Phytol. 2014, 203, 32–43. [Google Scholar] [CrossRef]
- Kim, T.H.; Hauser, F.; Ha, T.; Xue, S.; Böhmer, M.; Nishimura, N.; Munemasa, S.; Hubbard, K.; Peine, N.; Lee, B.H.; et al. Chemical Genetics reveals Negative regulation of Abscisic acid Signaling by a Plant Immune Response Pathway. Curr. Biol. 2011, 21, 990–997. [Google Scholar] [CrossRef] [PubMed]
- Clouse, S.D.; Sasse, J.M. BRASSINOSTEROIDS: Essential Regulators of Plant Growth and Development. Annu. Rev. Plant Biol. 1998, 49, 427–451. [Google Scholar] [CrossRef]
- Hu, Y.; Xia, S.; Su, Y.; Wang, H.; Luo, W.; Su, S.; Xiao, L. Brassinolide Increases Potato Root Growth in Vitro in a Dose-dependent Way and Alleviates Salinity Stress. BioMed Res. Int. 2016. [Google Scholar] [CrossRef]
- Efimova, M.V.; Khripach, V.A.; Boyko, E.V.; Malofii, M.K.; Kolomeichuk, L.V.; Murgan, O.K.; Vidershpun, A.N.; Mukhamatdinova, E.A.; Kuznetsov, V.V. The Priming of Potato Plants Induced by Brassinosteroids Reduces Oxidative Stress and Increases Salt Tolerance. Dokl. Biol. Sci. 2018, 478, 33–36. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Zhang, N.; Yang, J.; Tang, X.; Wen, Y.; Si, H. Functional analysis of StDWF4 gene in response to salt stress in potato. Plant Physiol. Biochem. 2018, 125, 63–73. [Google Scholar] [CrossRef]
- Azaizeh, H.; Gunse, B.; Steudle, E. Effects of NaCl and CaCl2 on Water Transport across Root Cells Of Maize (Zea mays L.) seedlings. Plant Physiol. 1992, 99, 886–894. [Google Scholar] [CrossRef] [PubMed]
- Levy, D.; Coleman, W.K.; Veilleux, R.E. Adaptation of Potato to Water Shortage: Irrigation Management and Enhancement of Tolerance to Drought and Salinity. Am. J. Potato Res. 2013, 90, 186–206. [Google Scholar] [CrossRef]
- Ruiz-Lozano, J.M.; Porcel, R.; Azcón, C.; Aroca, R. Regulation by Arbuscular mycorrhizae of the integrated Physiological Response to Salinity in Plants: New Challenges in Physiological and Molecular Studies. J. Exp. Bot. 2012, 63, 4033–4044. [Google Scholar] [CrossRef]
- Negrão, S.; Schmöckel, S.M.; Tester, M. Evaluating Physiological Responses of Plants to Salinity Stress. Ann. Bot. 2017, 119, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Aliche, E.B.; Prusova-Bourke, A.; Ruiz-Sanchez, M.; Oortwijn, M.; Gerkema, E.; Van As, H.; Visser, R.G.F.; van der Linden, C.G. Morphological and Physiological Responses of the Potato Stem Transport tissues to Dehydration Stress. Planta 2020, 251, 1–15. [Google Scholar] [CrossRef]
- Ali, E.F.; Hassan, F.A.S. Alleviatory Effects of Salt Stress by Mycorrhizal Fungi and Gibberellic Acid on Chamomile Plant. Int. J. Sci. Res. 2014, 3, 109–118. [Google Scholar]
- Antunes, W.C.; Provart, N.J.; Williams, T.C.R.; Loureiro, M.E. Changes in Stomatal Function and Water use efficiency in Potato Plants with altered Sucrolytic Activity. Plant Cell Environ. 2012, 35, 747–759. [Google Scholar] [CrossRef]
- Gao, H.J.; Yang, H.Y.; Bai, J.P.; Liang, X.Y.; Lou, Y.; Zhang, J.L.; Wang, D.; Zhang, J.L.; Niu, S.Q.; Chen, Y.L. Ultrastructural and Physiological Responses of Potato (Solanum tuberosum L) Plantlets to Gradient Saline Stress. Front. Plant Sci. 2015, 5. [Google Scholar] [CrossRef] [PubMed]
- Neumann, G.; Römheld, V. Root Excretion of Carboxylic acids and Protons in Phosphorus-deficient Plants. Plant Soil. 1999, 211, 121–130. [Google Scholar] [CrossRef]
- Céccoli, G.; Ramos, J.C.; Ortega, L.I.; Acosta, J.M.; Perreta, M.G. Salinity Induced Anatomical and Morphological Changes in Chloris gayana Kunth roots. Biocell 2011, 35, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Atabayeva, S.; Nurmahanova, A.; Minocha, S.; Ahmetova, A.; Kenzhebayeva, S.; Aidosova, S.; Nurzhanova, A.; Zhardamalieva, A.; Asrandina, S.; Alybayeva, R.; et al. The Effect of Salinity on growth and Anatomical attributes of Barley Seedling (Hordeum vulgare L.). Afr. J. Biotechnol. 2013, 12, 2366. [Google Scholar]
- Sarker, U.; Oba, S. Salinity Stress Enhances Color Parameters, Bioactive Leaf Pigments, Vitamins, Polyphenols, Flavonoids and Antioxidant activity in selected Amaranthus leafy vegetables. J. Sci. Food Agric. 2019, 99, 2275–2284. [Google Scholar] [CrossRef] [PubMed]
- Kaya, C.; Tuna, A.L.; Yokaş, I. The Role of Plant Hormones in Plants Under Salinity Stress. Salinity Water Stress. 2009, 44, 45–50. [Google Scholar]
- Abdel Latef, A.A.H.; Chaoxing, H. Effect of Arbuscular mycorrhizal fungi on Growth, Mineral nutrition, Antioxidant enzymes activity and Fruit Yield of Tomato grown under Salinity Stress. Sci. Hortic. 2011, 127, 228–233. [Google Scholar] [CrossRef]
- Ashraf, M.; Akram, N.A. Improving Salinity Tolerance of Plants through Conventional Breeding and Genetic Engineering: An analytical Comparison. Biotechnol. Adv. 2009, 27, 744–752. [Google Scholar] [CrossRef] [PubMed]
- Charfeddine, M.; Charfeddine, S.; Ghazala, I.; Bouaziz, D.; Bouzid, R.G. Investigation of the Response to Salinity of Transgenic Potato Plants Overexpressing the Transcription factor StERF94. J. Biosci. 2019, 44. [Google Scholar] [CrossRef]
- Kolomeichuk, L.V.; Efimova, M.V.; Zlobin, I.E.; Kreslavski, V.D.; Murgan, O.K.; Kovtun, I.S.; Khripach, V.A.; Kuznetsov, V.V.; Allakhverdiev, S.I. 24-Epibrassinolide Alleviates the Toxic effects of NaCl on Photosynthetic Processes in Potato Plants. Photosynth. Res. 2020, 146, 151–163. [Google Scholar] [CrossRef]
- Mehta, P.; Jajoo, A.; Mathur, S.; Bharti, S. Chlorophyll a Fluorescence Study Revealing Effects of High Salt Stress on Photosystem II in wheat leaves. Plant Physiol. Biochem. 2010, 48, 16–20. [Google Scholar] [CrossRef] [PubMed]
- Ramegowda, V.; Da Costa, M.V.J.; Harihar, S.; Karaba, N.N.; Sreeman, S.M. Abiotic and Biotic Stress Interactions in Plants: A Cross-tolerance Perspective. In Priming-Mediated Stress and Cross-Stress Tolerance in Crop Plants; Hossain, M.A., Liu, F., Burritt, D., Fujita, M., Huang, B., Eds.; Elsevier: Amsterdam, The Netherlands, 2020; pp. 267–302. [Google Scholar]
- Choudhary, A.; Kumar, A.; Kaur, N. ROS and Oxidative Burst: Roots in Plant Development. Plant Divers. 2020, 42, 33–43. [Google Scholar] [CrossRef]
- Okon, O.G. Effect of Salinity on Physiological Processes in Plants. In Microorganisms in Saline Environments: Strategies and Functions; Giri, B., Varma, A., Eds.; Springer: Berlin, Germany, 2019; pp. 237–262. [Google Scholar]
- Zandalinas, S.I.; Mittler, R.; Balfagón, D.; Arbona, V.; Gómez-Cadenas, A. Plant Adaptations to the Combination of Drought and High Temperatures. Physiol. Plant 2018, 162, 2–12. [Google Scholar] [CrossRef] [PubMed]
- Kim, I.S.; Park, S. Ultrastructural Characteristics of three Chenopod Halophytes lacking Salt excretion Structures. J. Plant Biol. 2010, 53, 314–320. [Google Scholar] [CrossRef]
- Benavídes, M.P.; Marconi, P.L.; Gallego, S.M.; Comba, M.E.; Tomaro, M.L. Relationship between Antioxidant Defence Systems and Salt Tolerance in Solanum tuberosum. Aust. J. Plant Physiol. 2000, 27, 273–278. [Google Scholar] [CrossRef]
- Eskandari, H.; Ehsanpour, A.A.; Al-Mansour, N.; Bardania, H.; Sutherland, D.; Mohammad-Beigi, H. Rosmarinic acid inhibits programmed cell death in Solanum tuberosum L. calli under high salinity. Plant Physiol. Biochem. 2020, 147, 54–65. [Google Scholar] [CrossRef]
- Upadhyaya, C.P.; Venkatesh, J.; Gururani, M.A.; Asnin, L.; Sharma, K.; Ajappala, H.; Park, S.W. Transgenic Potato Overproducing L-ascorbic acid resisted an increase in Methylglyoxal under Salinity Stress via maintaining higher reduced Glutathione level and Glyoxalase Enzyme activity. Biotechnol. Lett. 2011, 33, 2297–2307. [Google Scholar] [CrossRef]
- Singh, M.; Kumar, J.; Singh, S.; Singh, V.P.; Prasad, S.M. Roles of Osmoprotectants in improving Salinity and Drought Tolerance in Plants: A review. Rev. Environ. Sci. Biotechnol. 2015, 14, 407–426. [Google Scholar] [CrossRef]
- Ashraf, M.; Harris, P.J.C. Potential biochemical indicators of salinity tolerance in plants. Plant Sci. 2004, 166, 3–16. [Google Scholar] [CrossRef]
- Hussain Wani, S.; Brajendra Singh, N.; Haribhushan, A.; Iqbal Mir, J. Compatible Solute Engineering in Plants for Abiotic Stress Tolerance—Role of Glycine Betaine. Curr. Genomics 2013, 14, 157–165. [Google Scholar] [CrossRef] [PubMed]
- Parray, J.A.; Yaseen Mir, M.; Shameem, N.; Parray, J.A.; Yaseen Mir, M.; Shameem, N. Stress Management: Sustainable Approach Towards Resilient Agriculture. In Sustainable Agriculture: Biotechniques in Plant Biology; Parray, J., Yaseen Mir, M., Shameem, N., Eds.; Springer: Singapore, 2019; pp. 231–270. [Google Scholar]
- Dikilitas, M.; Simsek, E.; Roychoudhury, A. Role of Proline and Glycine Betaine in Overcoming Abiotic Stresses. Prot. Chem. Agents Amelior. Plant Abiotic Stress Biochem. Mol. Perspect. 2020, 1–23. [Google Scholar]
- Joshi, R.; Sahoo, K.K.; Singh, A.K.; Anwar, K.; Pundir, P.; Gautam, R.K.; Krishnamurthy, S.L.; Sopory, S.K.; Pareek, A.; Singla-Pareek, S.L. Enhancing Trehalose biosynthesis improves Yield potential in Marker-free Transgenic Rice under Drought, Saline, and Sodic Conditions. J. Exp. Bot. 2020, 71, 653–668. [Google Scholar] [CrossRef]
- Jain, N.K.; Roy, I. Effect of Trehalose on Protein Structure. Protein Sci. 2009, 18, 24–36. [Google Scholar] [CrossRef] [PubMed]
- Anamul Hoque, M.; Okuma, E.; Nasrin Akhter Banu, M.; Nakamura, Y.; Shimoishi, Y.; Murata, Y. Exogenous Proline Mitigates the detrimental effects of Salt Stress more than Exogenous Betaine by increasing Antioxidant Enzyme Activities. J. Plant Physiol. 2007, 164, 553–561. [Google Scholar] [CrossRef]
- Deinlein, U.; Stephan, A.B.; Horie, T.; Luo, W.; Xu, G.; Schroeder, J.I. Plant salt-tolerance mechanisms. Trends Plant Sci. 2014, 19, 371–379. [Google Scholar] [CrossRef]
- Daneshmand, F.; Arvin, M.J.; Kalantari, K.M. Physiological Responses to NaCl stress in three wild species of Potato in vitro. Acta Physiol. Plant 2010, 32, 91–101. [Google Scholar] [CrossRef]
- Liu, J.; Zhu, J.K. Proline accumulation and Salt-stress-induced gene expression in a Salt-hypersensitive mutant of Arabidopsis. Plant Physiol. 1997, 114, 591–596. [Google Scholar] [CrossRef]
- Rasool, S.; Hameed, A.; Azooz, M.M.; Muneeb-U-Rehman; Siddiqi, T.O.; Ahmad, P. Salt stress: Causes, types and responses of Plants. In Ecophysiology and Responses of Plants under Salt Stress; Ahmad, P., Azooz, M.M., Prasad, M.N.V., Eds.; Springer: New York, NY, USA, 2012; pp. 1–24. [Google Scholar]
- Kanayama, Y. Physiological roles of polyols in horticultural crops. J. Jpn. Soc. Hortic. Sci. 2009, 78, 158–168. [Google Scholar] [CrossRef]
- Lal, M.K.; Tiwari, R.K.; Kumar, R.; Naga, K.C.; Kumar, A.; Singh, B.; Raigond, P.; Dutt, S.; Chourasia, K.N.; Kumar, D.; et al. Effect of potato apical leaf curl disease on glycemic index and resistant starch of potato (Solanum tuberosum L.) tubers. Food Chem. 2021, 359, 129939. [Google Scholar] [CrossRef]
- Kissoudis, C.; Chowdhury, R.; van Heusden, S.; van de Wiel, C.; Finkers, R.; Visser, R.G.F.; Bai, Y.; van der Linden, G. Combined Biotic and Abiotic stress resistance in Tomato. Euphytica 2015, 202, 317–332. [Google Scholar] [CrossRef]
- Munns, R.; Gilliham, M. Salinity tolerance of crops—What is the cost? New Phytol. 2015, 208, 668–673. [Google Scholar] [CrossRef]
- Saijo, Y.; Loo, E.P. iian Plant Immunity in Signal Integration between Biotic and Abiotic Stress Responses. New Phytol. 2020, 225, 87–104. [Google Scholar] [CrossRef]
- Martinez, V.; Nieves-Cordones, M.; Lopez-Delacalle, M.; Rodenas, R.; Mestre, T.C.; Garcia-Sanchez, F.; Rubio, F.; Nortes, P.A.; Mittler, R.; Rivero, R.M. Tolerance to Stress combination in Tomato Plants: New Insights in the Protective Role of Melatonin. Molecules 2018, 23, 535. [Google Scholar] [CrossRef] [PubMed]
- Snapp, S.S.; Shennan, C.; van Bruggen, A.H.C. Effects of Salinity on Severity of Infection by Phytophthora parasitica Dast., Ion Concentrations and Growth of Tomato, Lycopersicon esculentum Mill. New Phytol. 1991, 119, 275–284. [Google Scholar] [CrossRef] [PubMed]
- Chojak, J.; Kuźniak, E.; Świercz, U.; Sekulska-Nalewajko, J.; Gocławski, J. Interaction Between Salt Stress and Angular Leaf Spot (Pseudomonas Syringae Pv Lachrymans) in Cucumber. Veg. Crop. Res. Bull. 2012, 77, 5–16. [Google Scholar] [CrossRef]
- You, M.; Colmer, T.; Biology, M.B.-F.P. Undefined Salinity drives Host reaction in Phaseolus vulgaris (common bean) to Macrophomina phaseolina. CSIRO 2011, 38, 984–992. [Google Scholar]
- Rasmussen, S.L. Effect of Salinity Stress on Development of Pythium Blight in Agrostis palustris. Phytopathology 1988, 78, 1495. [Google Scholar] [CrossRef]
- Howell, A.; Francois, L.; Research, D.E.-F.C. Undefined Interactive effect of salinity and Verticillium albo-atrum on Verticillium Wilt disease Severity and Yield of two alfalfa cultivars. Field Crop. Res. 1994, 37, 247–251. [Google Scholar] [CrossRef]
- Souman, M.F.; Kostandi, S.F. Effect of Saline environment on Yield and Smut Disease Severity of different Corn Genotypes (Zea mays L.). J. Phytopathol. 1998, 146, 185–189. [Google Scholar] [CrossRef]
- Nayem, S.A.; Chowdhury, M.S.M.; Sultana, N.; Masum, G.Z.H.; Rahman, M.S.; Jamal, M.A.H.M. Combined effect of salt stress and Xanthomonas axonopodis pv citri on citrus (Citrus aurantifolia). Heliyon 2020, 6, e03403. [Google Scholar] [CrossRef] [PubMed]
- Nachmias, A.; Kaufman, Z.; Livescu, L.; Tsror, L.; Meiri, A.; Caligari, P.D.S. Effects of Salinity and its Interactions with Disease incidence on Potatoes grown in Hot Climates. Phytoparasitica 1993, 21, 245–255. [Google Scholar] [CrossRef]
- DiLeo, M.V.; Pye, M.F.; Roubtsova, T.V.; Duniway, J.M.; MacDonald, J.D.; Rizzo, D.M.; Bostock, R.M. Abscisic acid in salt stress predisposition to phytophthora root and crown rot in tomato and chrysanthemum. Phytopathology 2010, 100, 871–879. [Google Scholar] [CrossRef] [PubMed]
- Triky-Dotan, S.; Yermiyahu, U.; Katan, J.; Gamliel, A. Development of Crown and Root Rot Disease of Tomato under Irrigation with Saline water. Phytopathology 2005, 95, 1438–1444. [Google Scholar] [CrossRef]
- Bai, Y.; Kissoudis, C.; Yan, Z.; Visser, R.G.F.; van der Linden, G. Plant behaviour under combined stress: Tomato responses to combined salinity and pathogen stress. Plant J. 2018, 93, 781–793. [Google Scholar] [CrossRef]
- Thaler, J.S.; Bostock, R.M. Interactions between abscisic-acid-mediated responses and plant resistance to pathogens and insects. Ecology 2004, 85, 48–58. [Google Scholar] [CrossRef]
- Han, P.; Wang, Z.J.; Lavoir, A.V.; Michel, T.; Seassau, A.; Zheng, W.Y.; Niu, C.Y.; Desneux, N. Increased Water Salinity applied to Tomato Plants Accelerates the development of the Leaf Miner Tuta absoluta through Bottom-up Effects. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef]
- Eichele-Nelson, J.; DeSutter, T.; Wick, A.F.; Harmon, E.L.; Harmon, J.P. Salinity Improves Performance and Alters Distribution of Soybean Aphids. Environ. Entomol. 2018, 47, 875–880. [Google Scholar] [CrossRef] [PubMed]
- Polack, L.A.; Pereyra, P.C.; Sarandón, S.J. Effects of Plant Stress and Habitat Manipulation on Aphid Control in Greenhouse Sweet Peppers. J. Sustain. Agric. 2011, 35, 699–725. [Google Scholar] [CrossRef]
- Quais, M.K.; Ansari, N.A.; Wang, G.-Y.; Zhou, W.-W.; Zhu, Z.-R. Host Plant Salinity Stress Affects the Development and Population Parameters of Nilaparvata lugens (Hemiptera: Delphacidae). Environ. Entomol. 2019, 48, 1149–1161. [Google Scholar] [CrossRef]
- Chen, M.; Yang, Z.; Liu, J.; Zhu, T.; Wei, X.; Fan, H.; Wang, B. Adaptation mechanism of salt excluders under saline conditions and its applications. Int. J. Mol. Sci. 2018, 19, 3668. [Google Scholar] [CrossRef]
- Heřmanová, V.; Bárta, J.; Čurn, V. Wild potato species: Characterization and Biological Potential for Potato Breeding. Czech J. Genet. Plant Breed. 2007, 43, 73–81. [Google Scholar] [CrossRef]
- Shaterian, J.; Waterer, D.R.; de Jong, H.; Tanino, K.K. Methodologies and Traits for Evaluating the Salt Tolerance in Diploid Potato Clones. Am. J. Potato Res. 2008, 85, 93–100. [Google Scholar] [CrossRef]
- Jaarsma, R.; de Vries, R.S.M.; de Boer, A.H. Effect of Salt Stress on Growth, Na+ Accumulation and Proline Metabolism in Potato (Solanum tuberosum) Cultivars. PLoS ONE 2013, 8, e60183. [Google Scholar] [CrossRef]
- Ahmed, H.A.A.; Şahin, N.K.; Akdoğan, G.; Yaman, C.; Köm, D.; Uranbey, S. Variability in Salinity Stress Tolerance of Potato (Solanum tuberosum L.) varieties using in vitro screening. Cienc. Agrotecnologia 2020, 44, 1–14. [Google Scholar] [CrossRef]
- Van Straten, G.; Bruning, B.; de Vos, A.C.; González, A.P.; Rozema, J.; van Bodegom, P.M. Estimating Cultivar-specific Salt Tolerance model parameters from Multi-annual Field tests for identification of Salt Tolerant Potato Cultivars. Agric. Water Manag. 2021, 252, 106902. [Google Scholar] [CrossRef]
- Frary, A.; Göl, D.; Keleş, D.; Ökmen, B.; Pinar, H.; Şiǧva, H.T.; Yemenicioǧlu, A.; Doǧanlar, S. Salt tolerance in Solanum pennellii: Antioxidant response and related QTL. BMC Plant Biol. 2010, 10, 1–16. [Google Scholar] [CrossRef]
- Diouf, I.A.; Derivot, L.; Bitton, F.; Pascual, L.; Causse, M. Water deficit and salinity stress reveal many specific QTL for plant growth and fruit quality traits in tomato. Front. Plant Sci. 2018, 9, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Byun, M.; Kwon, H.; Park, S. Chapter 29 Recent Advances in Genetic Engineering of Potato Crops for. Mol. Breed. 2007, 713–737. [Google Scholar]
- Hmida-Sayari, A.; Gargouri-Bouzid, R.; Bidani, A.; Jaoua, L.; Savouré, A.; Jaoua, S. Overexpression of [delta]--pyrroline-5-carboxylate synthetase increases proline production and confers salt tolerance in transgenic potato plants. Plant Sci. 2005, 169, 746–752. [Google Scholar] [CrossRef]
- Behnam, B.; Kikuchi, A.; Celebi-Toprak, F.; Yamanaka, S.; Kasuga, M.; Yamaguchi-Shinozaki, K.; Watanabe, K.N. The Arabidopsis DREB1A gene driven by the stress-inducible rd29A promoter increases salt-stress tolerance in proportion to its copy number in tetrasomic tetraploid potato (Solanum tuberosum). Plant Biotechnol. 2006, 23, 169–177. [Google Scholar] [CrossRef]
- Turhan, H. Salinity Response of Transgenic Potato Genotypes Expressing the Oxalate Oxidase Gene. Turkish J. Agric. For. 2005, 29, 187–195. [Google Scholar] [CrossRef]
- Tian, Z.D.; Zhang, Y.; Liu, J.; Xie, C.H. Novel potato C2H2-type zinc finger protein gene, StZFP1, which responds to biotic and abiotic stress, plays a role in salt tolerance. Plant Biol. 2010, 12, 689–697. [Google Scholar] [CrossRef]
- Krivosheeva, A.B.; Varlamova, T.V.; Yurieva, N.O.; Sobol’kova, G.I.; Kholodova, V.P.; Belyaev, D.V. Potato transformation with the HvNHX3 gene and the improvement of transformant salt tolerance. Russ. J. Plant Physiol. 2014, 61, 792–800. [Google Scholar] [CrossRef]
- Tian, Z.; He, Q.; Wang, H.; Liu, Y.; Zhang, Y.; Shao, F.; Xie, C. The Potato ERF transcription factor StERF3 Negatively Regulates Resistance to Phytophthora infestans and Salt Tolerance in Potato. Plant Cell Physiol. 2014, 56, 992–1005. [Google Scholar] [CrossRef] [PubMed]
- Bayat, F.; Shiran, B.; Belyaev, D.V.; Yur’eva, N.O.; Sobol’kova, G.I.; Alizadeh, H.; Khodambashi, M.; Babakov, A.V. Potato Plants Bearing a Vacuolar Na+/H+ Antiporter HvNHX2 from Barley Are Characterized by Improved Salt Tolerance. Russ. J. Plant Physiol. 2010, 57, 696–706. [Google Scholar] [CrossRef]
- Bouaziz, D.; Pirrello, J.; Ben Amor, H.; Hammami, A.; Charfeddine, M.; Dhieb, A.; Bouzayen, M.; Gargouri-Bouzid, R. Ectopic expression of Dehydration Responsive Element Binding Proteins (StDREB2) confers higher Tolerance to Salt Stress in Potato. Plant Physiol. Biochem. 2012, 60, 98–108. [Google Scholar] [CrossRef]
- Cheng, Y.J.; Kim, M.D.; Deng, X.P.; Kwak, S.S.; Chen, W. Enhanced Salt Stress Tolerance in Transgenic Potato Plants Expressing ibMYB1, A Sweet Potato Transcription Factor. J. Microbiol. Biotechnol. 2013, 23, 1737–1746. [Google Scholar] [CrossRef]
- Liu, M.; Li, Y.; Li, G.; Dong, T.; Liu, S.; Liu, P.; Wang, Q. guo Overexpression of StCYS1 gene Enhances Tolerance to Salt stress in the Transgenic Potato (Solanum tuberosum L.) plant. J. Integr. Agric. 2020, 19, 2239–2246. [Google Scholar] [CrossRef]
- Gangadhar, B.H.; Sajeesh, K.; Venkatesh, J.; Baskar, V.; Abhinandan, K.; Yu, J.W.; Prasad, R.; Mishra, R.K. Enhanced Tolerance of Transgenic Potato Plants Over-expressing Non-specific Lipid Transfer Protein-1 (StnsLTP1) against Multiple Abiotic Stresses. Front. Plant Sci. 2016, 7, 1–12. [Google Scholar] [CrossRef]
- Shi, J.; Wang, J.; Wang, N.; Zhou, H.; Xu, Q.; Yan, G. Overexpression of StGA2ox1 Gene Increases the Tolerance to Abiotic Stress in Transgenic Potato (Solanum tuberosum L.) Plants. Appl. Biochem. Biotechnol. 2019, 187, 1204–1219. [Google Scholar] [CrossRef]
- Jeong, M.J.; Park, S.C.; Byun, M.O. Improvement of Salt Tolerance in Transgenic Potato Plants by Glyceraldehyde-3 Phosphate dehydrogenase Gene Transfer. Mol. Cells 2001, 12, 185–189. [Google Scholar]
- Xu, Q.; He, Q.; Tian, Z. Molecular characterization of StNAC2 in potato and its overexpression confers drought and salt tolerance. Acta Physiol. Plant 2014, 36, 1841–1851. [Google Scholar] [CrossRef]
- Shafi, A.; Pal, A.K.; Sharma, V.; Kalia, S.; Kumar, S.; Ahuja, P.S.; Singh, A.K. Transgenic Potato Plants Overexpressing SOD and APX Exhibit Enhanced Lignification and Starch Biosynthesis with Improved Salt Stress Tolerance. Plant Mol. Biol. Report. 2017, 35, 504–518. [Google Scholar] [CrossRef]
- Zhao, P.; Wang, D.; Wang, R.; Kong, N.; Zhang, C.; Yang, C.; Wu, W.; Ma, H.; Chen, Q. Genome-wide Analysis of the Potato Hsp20 gene family: Identification, Genomic Organization and Expression Profiles in Response to Heat Stress. BMC Genom. 2018, 19, 1–13. [Google Scholar] [CrossRef]
- Ali, A.; Ali, Q.; Iqbal, M.S.; Nasir, I.A.; Wang, X. Salt Tolerance of Potato Genetically Engineered with the Atriplex canescens BADH Gene. Biol. Plant 2020, 64, 271–279. [Google Scholar] [CrossRef]
- Shelake, R.M.; Pramanik, D.; Kim, J.-Y. Exploration of Plant-microbe Interactions for Sustainable Agriculture in CRISPR Era. Microorganisms 2019, 7, 269. [Google Scholar] [CrossRef]
- Shelake, R.M.; Pramanik, D.; Kim, J.-Y. Evolution of Plant Mutagenesis Tools: A Shifting Paradigm from Random to Targeted Genome Editing. Plant Biotechnol. Rep. 2019, 13, 423–445. [Google Scholar] [CrossRef]
- Pramanik, D.; Shelake, R.M.; Kim, M.J.; Kim, J.-Y. CRISPR-mediated Engineering across the Central Dogma in Plant Biology for Basic Research and Crop Improvement. Mol. Plant 2021, 14, 127–150. [Google Scholar] [CrossRef]
- Zhou, J.; Deng, K.; Cheng, Y.; Zhong, Z.; Tian, L.; Tang, X.; Tang, A.; Zheng, X.; Zhang, T.; Qi, Y.; et al. CRISPR-Cas9 Based Genome Editing Reveals New Insights into Micro RNA Function and Regulation in Rice. Front. Plant Sci. 2017, 13, 1598. [Google Scholar] [CrossRef]
- Pramanik, D.; Shelake, R.M.; Park, J.; Kim, M.J.; Hwang, I.; Park, Y.; Kim, J.-Y. CRISPR/Cas9-mediated Generation of Pathogen-Resistant Tomato against Tomato Yellow Leaf Curl Virus and Powdery Mildew. Int. J. Mol. Sci. 2021, 22, 1878. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Zhang, S.; Wang, W.; Xiong, X.; Meng, F.; Cui, X. Efficient targeted mutagenesis in potato by the CRISPR/Cas9 system. Plant Cell Rep. 2015, 34, 1473–1476. [Google Scholar] [CrossRef]
- Andersson, M.; Turesson, H.; Olsson, N.; Fält, A.S.; Ohlsson, P.; Gonzalez, M.N.; Samuelsson, M.; Hofvander, P. Genome Editing in Potato Via CRISPR/Cas9 Ribonucleoprotein Delivery. Physiol. Plant 2018, 164, 378–384. [Google Scholar] [CrossRef] [PubMed]
- Makhotenko, A.V.; Khromov, A.V.; Snigir, E.A.; Makarova, S.S.; Makarov, V.V.; Suprunova, T.P.; Kalinina, N.O.; Taliansky, M.E. Functional Analysis of Coilin in Virus Resistance and Stress Tolerance of Potato Solanum tuberosum using CRISPR-Cas9 editing. Dokl. Biochem. Biophys. 2019, 484, 88–91, Pleiades Publishing. [Google Scholar] [CrossRef] [PubMed]
- Veillet, F.; Perrot, L.; Chauvin, L.; Kermarrec, M.P.; Guyon-Debast, A.; Chauvin, J.E.; Nogué, F.; Mazier, M. Transgene-free Genome Editing in Tomato and Potato Plants using Agrobacterium-mediated Delivery of a CRISPR/Cas9 Cytidine Base Editor. Int. J. Mol. Sci. 2019, 20, 402. [Google Scholar] [CrossRef]
- Veillet, F.; Kermarrec, M.P.; Chauvin, L.; Chauvin, J.E.; Nogué, F. CRISPR-induced indels and base editing using the Staphylococ-cus aureus Cas9 in potato. PLoS ONE 2020, 15, 17e0235942. [Google Scholar] [CrossRef]
- Bánfalvi, Z.; Csákvári, E.; Villányi, V.; Kondrák, M. Generation of Transgene-free PDS Mutants in Potato by Agrobacterium-mediated transformation. BMC Biotechnol. 2020, 20, 1–10. [Google Scholar] [CrossRef]
- González, M.N.; Massa, G.A.; Andersson, M.; Turesson, H.; Olsson, N.; Fält, A.S.; Storani, L.; Décima Oneto, C.A.; Hofvander, P.; Feingold, S.E. Reduced Enzymatic Browning in Potato Tubers by Specific Editing of a polyphenol oxidase gene via Ribonucleoprotein Complexes Delivery of the CRISPR/Cas9 System. Front. Plant Sci. 2020, 9, 1649. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Source | Results | Description | Reference |
|---|---|---|---|---|
| Mannitol 1-phosphate dehydrogenase (mtlD) gene | Escherichia coli | Increased the mannitol accumulation | Enhanced tolerance at 100 mM NaCl of the transgenic lines both in vitro and in hydroponic culture | [72] |
| AtHKT1 | Arabidopsis thaliana | Decrease Na+ content and improving K+/Na+ ratio in plant leaves | Tolerant to salinity stress by reducing Na+ accumulation in leaves and promoting K+/Na+ homeostasis | [77] |
| StDWF4 | Solanum tuberosum L cv. Zihuabai | Over-expression of StDWF4 gene | Accumulation of osmolytes enhanced by BRs, resulting in the tolerance to salinity | [92] |
| Pyrroline-5-carboxylate synthetase (P5CS) | Arabidopsis thaliana | Enhanced Proline accumulation | Enhanced proline accumulation with improved tolerance to salinity | [162] |
| Dehydration-responsive element-binding protein 1A (DREB1A) | Arabidopsis thaliana | Increased expression of DREB1A gene driven by the rd29A promoter | Two transgenic line out of 120 highly tolerant to salinity. Correlation of DREB1A copy number and mean tolerance level. | [163] |
| Oxalate oxidase gene | Hordeum vulgare root | Enhanced oxalate oxidase enzyme activity | Higher salinity tolerance due to increased activity of oxalate oxidase enzyme | [164] |
| Zinc finger protein (StZFP1) gene | Solanum tuberosum | Increased expression of StZFP1 gene driven by the rd29A promoter | Increased root number, root initiation and root elongation at 200 mM NaCl concentration | [165] |
| NHX antiporter HvNHX3 gene | Hordeum vulgare cv. Elo | Larger biomass and height of transformed plants | Improved salt tolerance at 100 mM NaCl concentration | [166] |
| Antiporter gene HvNHX2 | Hordeum vulgare | Higher potassium was found in roots of transgenic plants | Salt-tolerant transgenic plants had longer roots, higher dry weight, and suppressed cell expansion as compared to wild-type plants. | [168] |
| StDREB2 | Solanum tuberosum cv. Nicola | Over-expression of StDREB2 gene | Enhanced tolerance at 200 mM NaCl via the regulation of ABA hormone signaling and through a mechanism allowing proline synthesis | [169] |
| IbMYB1 | Ipomoea batatas | Increased expression of IbMYB1 gene driven by the SWPA2 promoter | Affects secondary metabolism, which leads to improved tolerance ability in transgenic potatoes at 400 mM NaCl concentration | [170] |
| StERF3 | Solanum tuberosum | Silencing of the StERF3 gene elevate salt tolerance | Increased tolerance at 150 mM NaCl and increased tolerance to late blight | [171] |
| StCYS1 | Solanum tuberosum L cv. Netherland 15 | Over-expression of StCYS1 gene | Increased accumulation of proline and chlorophyll, higher H2O2 scavenging capability and cell membrane integrity under salt stress condition | [172] |
| StnsLTP1 | Solanum tuberosum | Over-expression of StnsLTP1 gene | Tolerance to multiple abiotic stresses through enhanced activation of antioxidative defense mechanisms | [173] |
| StGA2ox1 | Solanum tuberosum | Over-expression of StGA2ox1 gene | Tolerance to multiple abiotic stresses through the accumulation of proline | [174] |
| Glyceraldehyde-3-phosphate dehydrogenase (GDP) | Pleurotus sajor-caju | Increased expression of GDP gene driven by the CaMV35S promoter | Improved salt tolerance and survived even after ten days at 2 M NaCl concentration | [175] |
| StNAC2 | Solanum tuberosum | Overexpression of StNAC2 | Transgenic line tolerant to both salt and drought stress | [175] |
| CuZn-superoxide dismutase (PaSOD) and ascorbate peroxide (RaAPX) gene | Potentilla atrosanguinea and Rheum australe | Increased the expression of SOD and APX enzymes | Physiological, anatomical, and molecular adjustments in the H2O2 regulated lignin biosynthesis signaling pathways resulting in salt stress tolerance | [176] |
| StHsp20 genes | Solanum tuberosum | Up-regulated expression of StHsp20 genes | Transgenic lines tolerant to multiple abiotic stress viz., salt, drought and heat stress | [177] |
| Betaine aldehyde dehydrogenase (BADH) gene | Atriplex canescens | Overexpression of BADH under triple CaMV 35S promoters | Tolerant to salinity and tolerance related to number of promoters in transgenic lines | [178] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chourasia, K.N.; Lal, M.K.; Tiwari, R.K.; Dev, D.; Kardile, H.B.; Patil, V.U.; Kumar, A.; Vanishree, G.; Kumar, D.; Bhardwaj, V.; et al. Salinity Stress in Potato: Understanding Physiological, Biochemical and Molecular Responses. Life 2021, 11, 545. https://doi.org/10.3390/life11060545
Chourasia KN, Lal MK, Tiwari RK, Dev D, Kardile HB, Patil VU, Kumar A, Vanishree G, Kumar D, Bhardwaj V, et al. Salinity Stress in Potato: Understanding Physiological, Biochemical and Molecular Responses. Life. 2021; 11(6):545. https://doi.org/10.3390/life11060545
Chicago/Turabian StyleChourasia, Kumar Nishant, Milan Kumar Lal, Rahul Kumar Tiwari, Devanshu Dev, Hemant Balasaheb Kardile, Virupaksh U. Patil, Amarjeet Kumar, Girimalla Vanishree, Dharmendra Kumar, Vinay Bhardwaj, and et al. 2021. "Salinity Stress in Potato: Understanding Physiological, Biochemical and Molecular Responses" Life 11, no. 6: 545. https://doi.org/10.3390/life11060545
APA StyleChourasia, K. N., Lal, M. K., Tiwari, R. K., Dev, D., Kardile, H. B., Patil, V. U., Kumar, A., Vanishree, G., Kumar, D., Bhardwaj, V., Meena, J. K., Mangal, V., Shelake, R. M., Kim, J.-Y., & Pramanik, D. (2021). Salinity Stress in Potato: Understanding Physiological, Biochemical and Molecular Responses. Life, 11(6), 545. https://doi.org/10.3390/life11060545