
Clinical Relevance of Secreted Small Noncoding RNAs in an Embryo Implantation Potential Prediction at Morula and Blastocyst Development Stages

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Clinical Characteristics of Couples
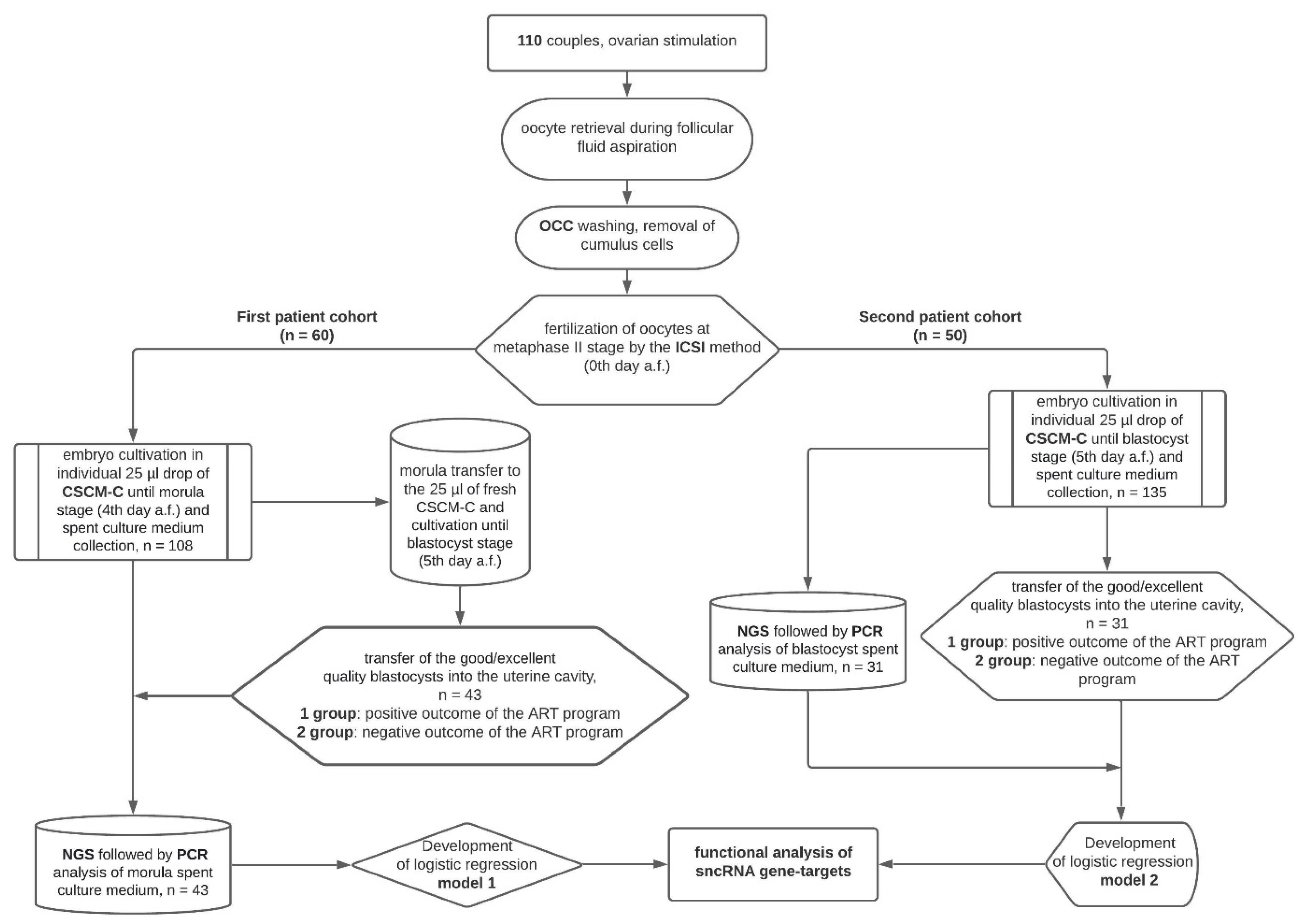
2.2. Experimental Design
2.3. Extraction of RNA from Spent Culture Medium
2.4. cDNA Library Preparation and RNA Deep Sequencing
2.5. Quantitative Real-Time RT-PCR
2.6. Statistical Analysis of the Obtained Data
3. Results
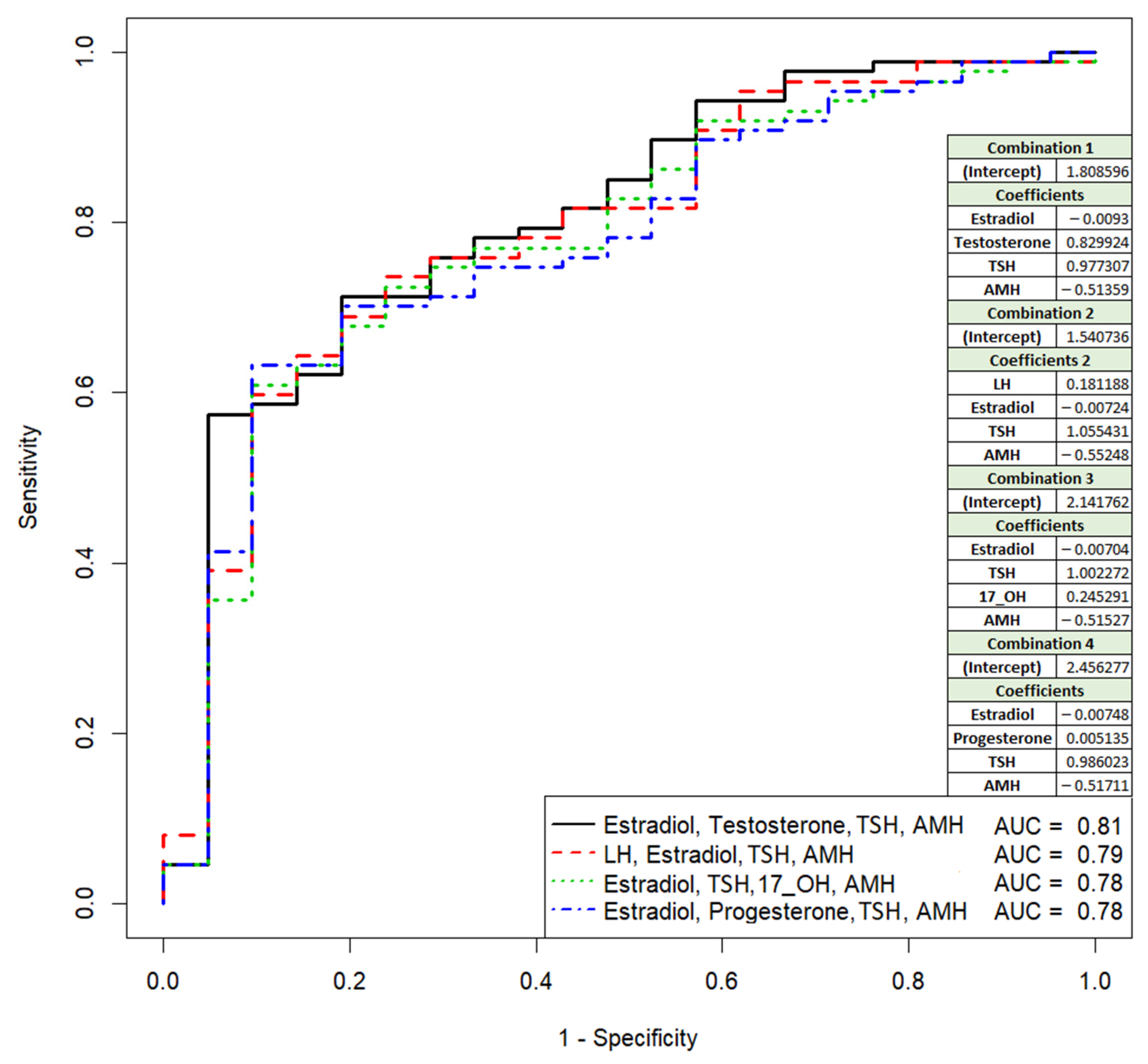
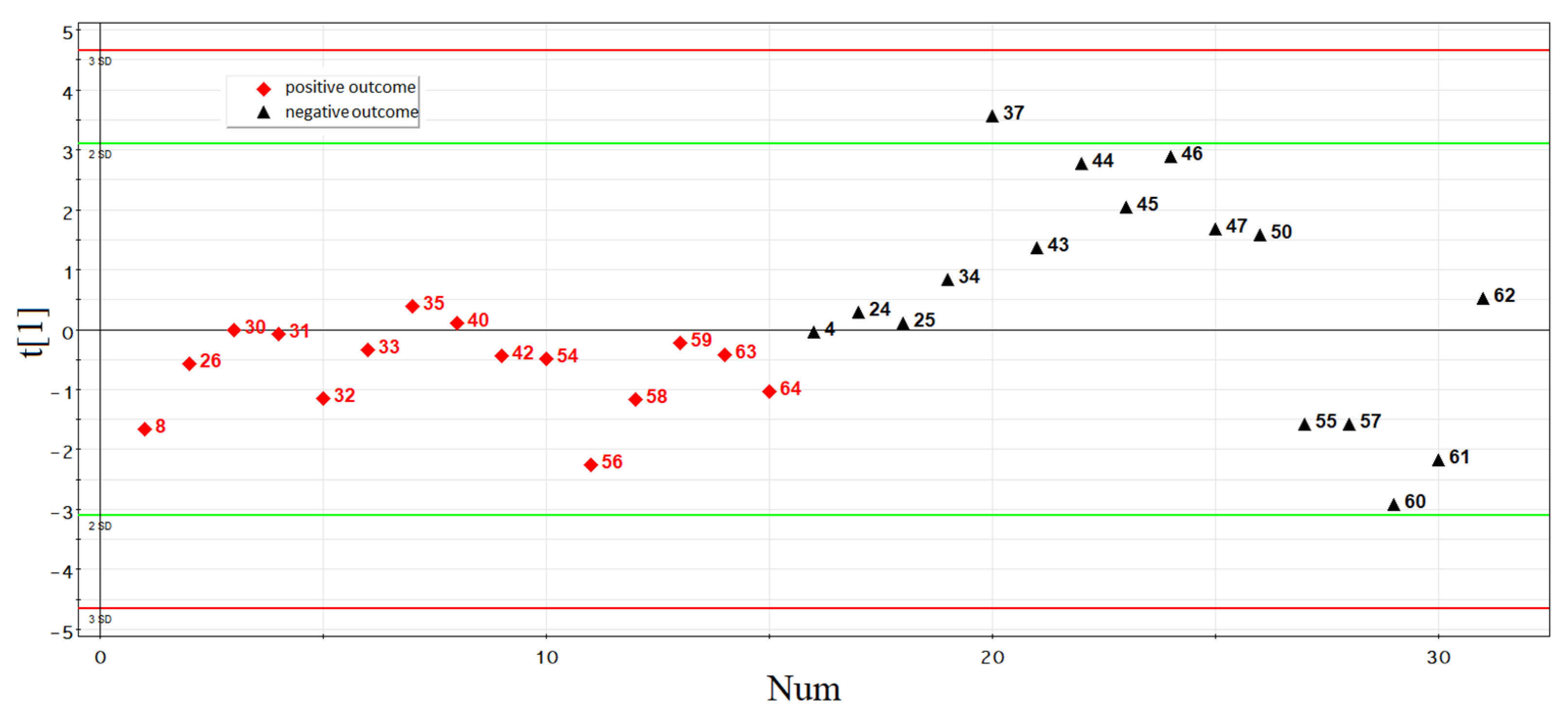
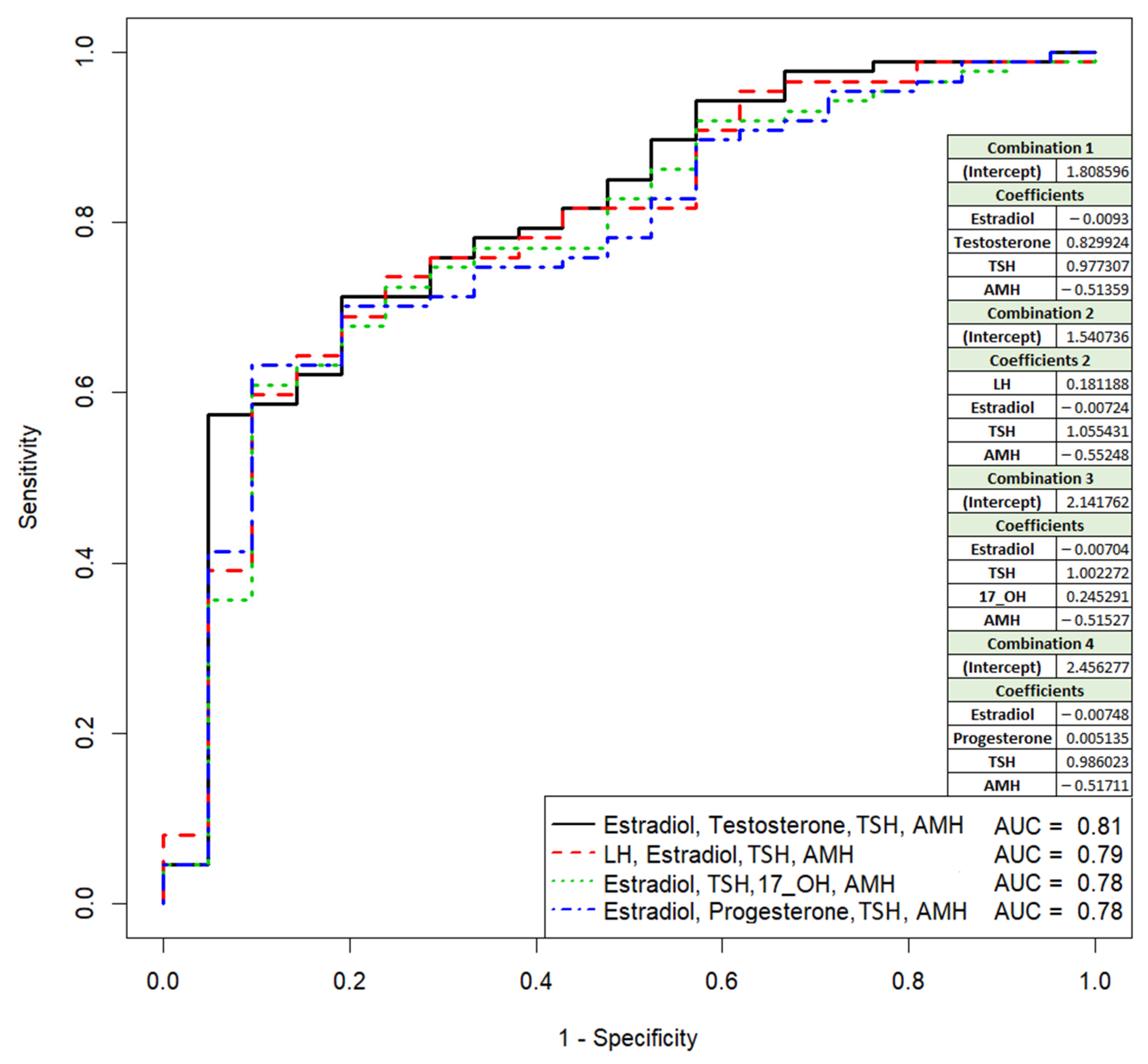
3.1. Comparative Analysis of Clinical Data and Parameters of the IVF/ICSI Protocol in Couples, Depending on the Outcome of the ART Program
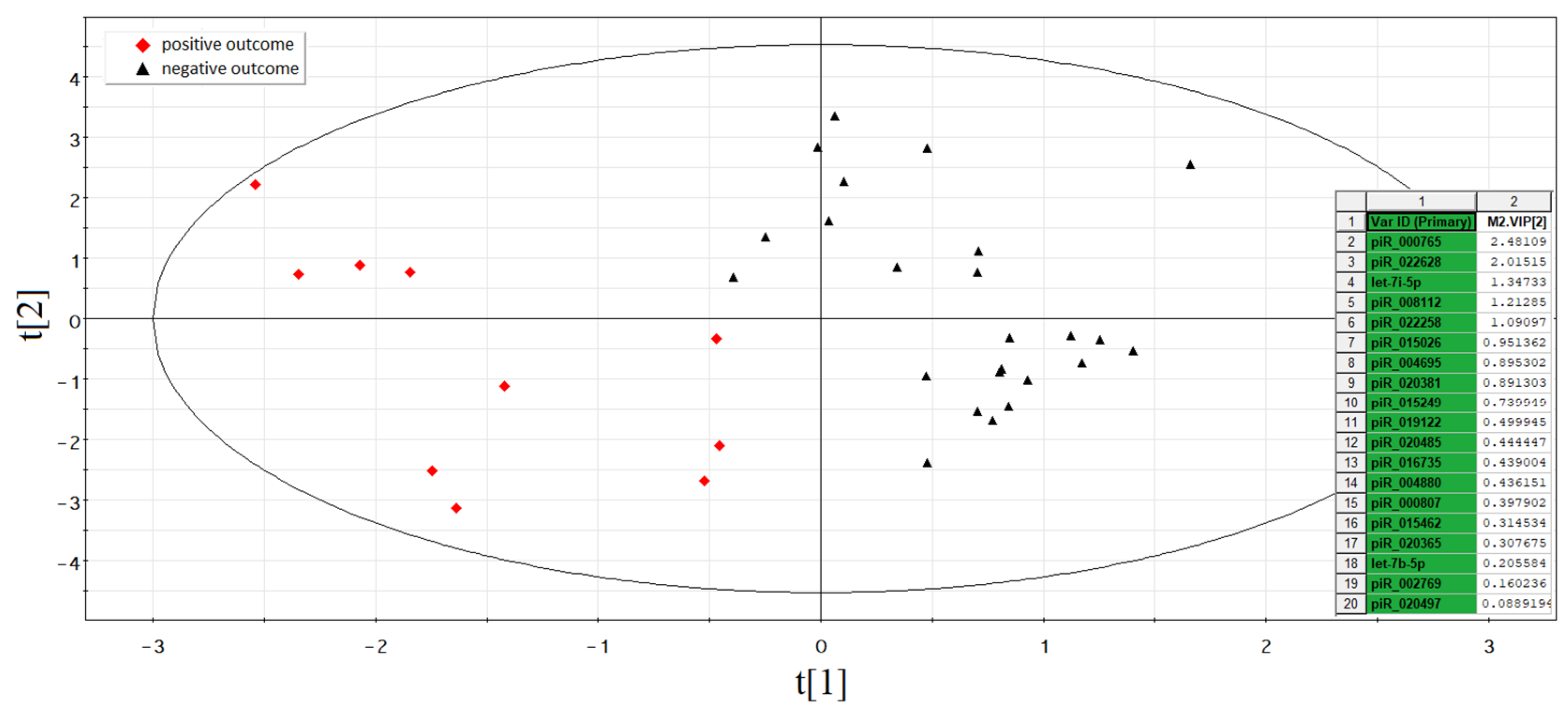
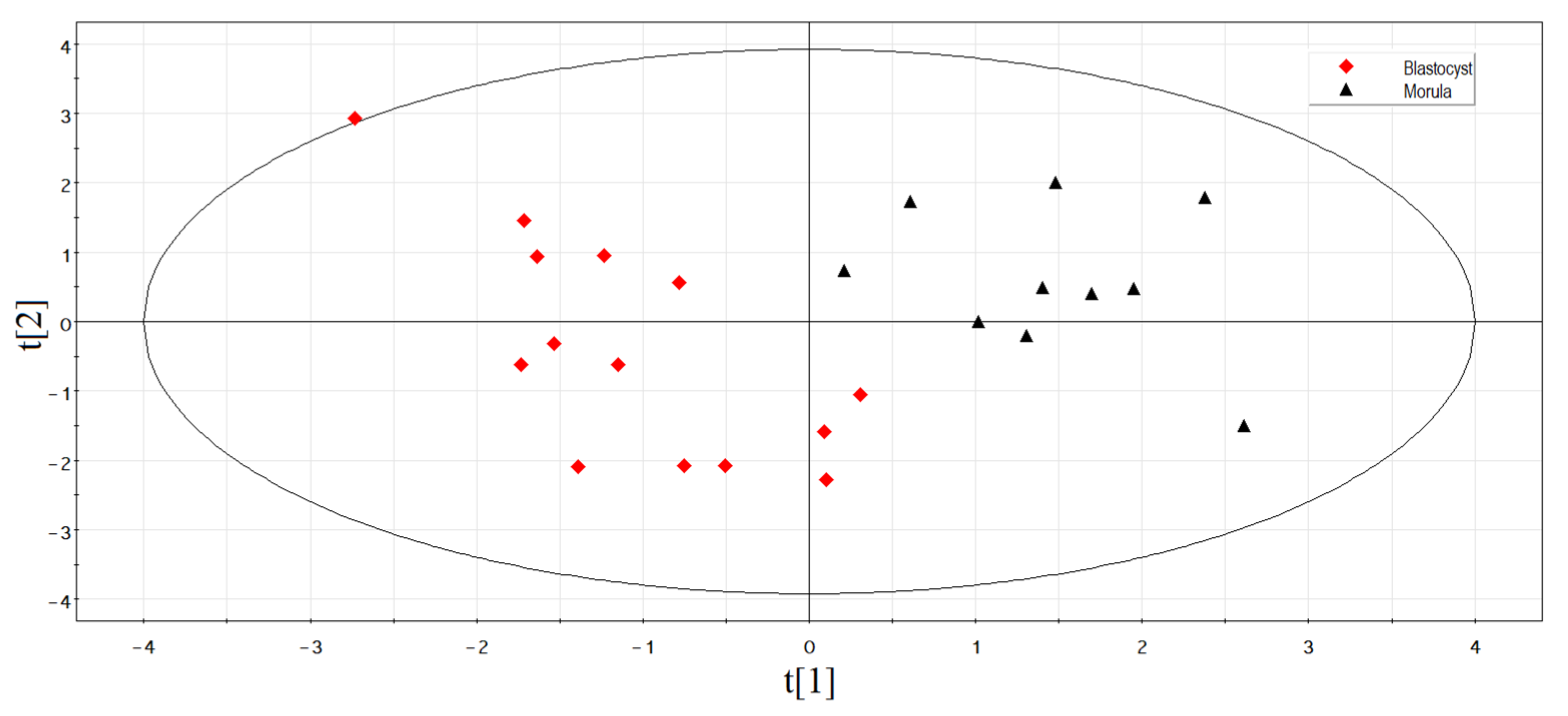
3.2. Identification of sncRNAs Associated with Implantation Potential of Embryo at Morula Stage
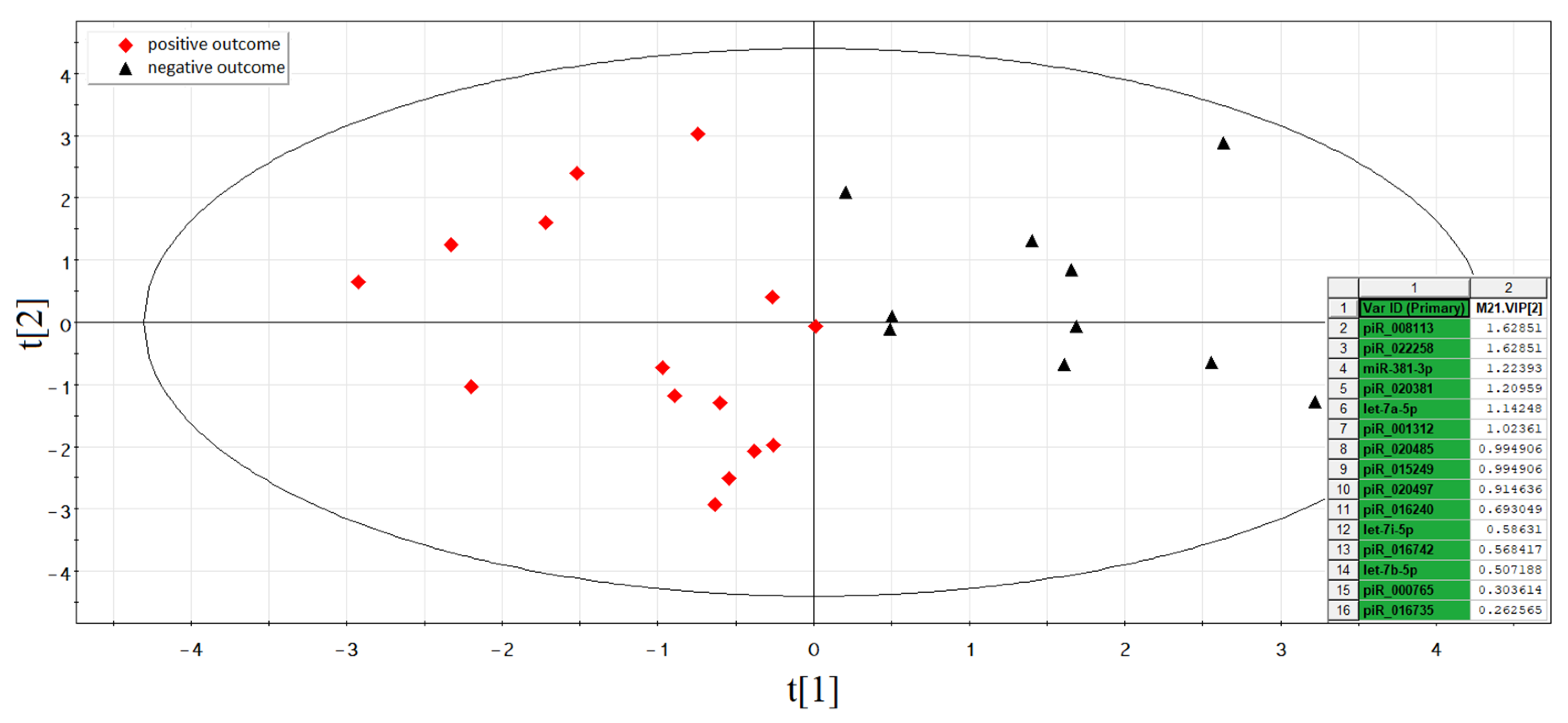
3.2.1. Deep Sequencing of the Spent Culture Medium
3.2.2. Quantitative RT-PCR Analysis in Real Time
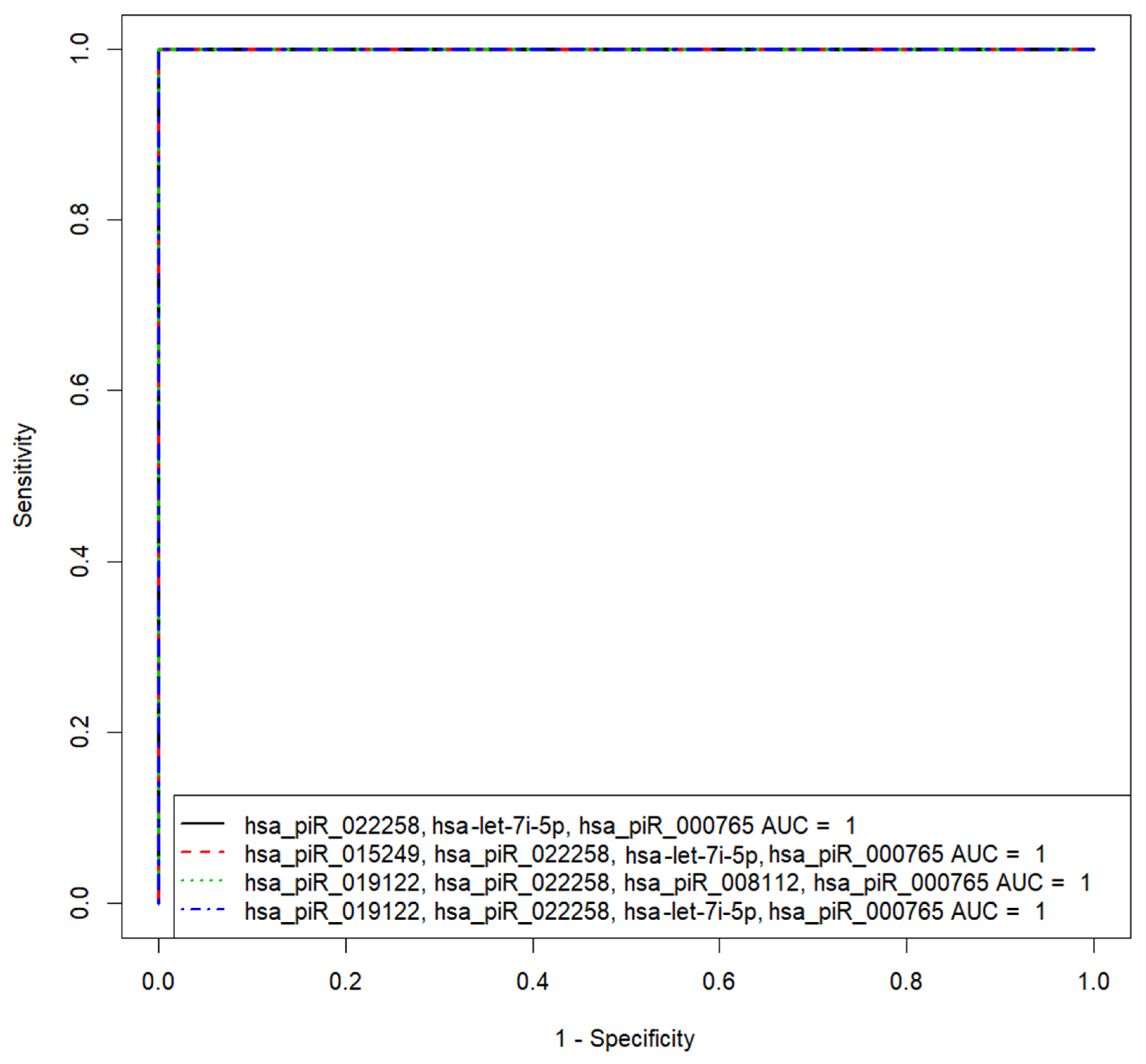
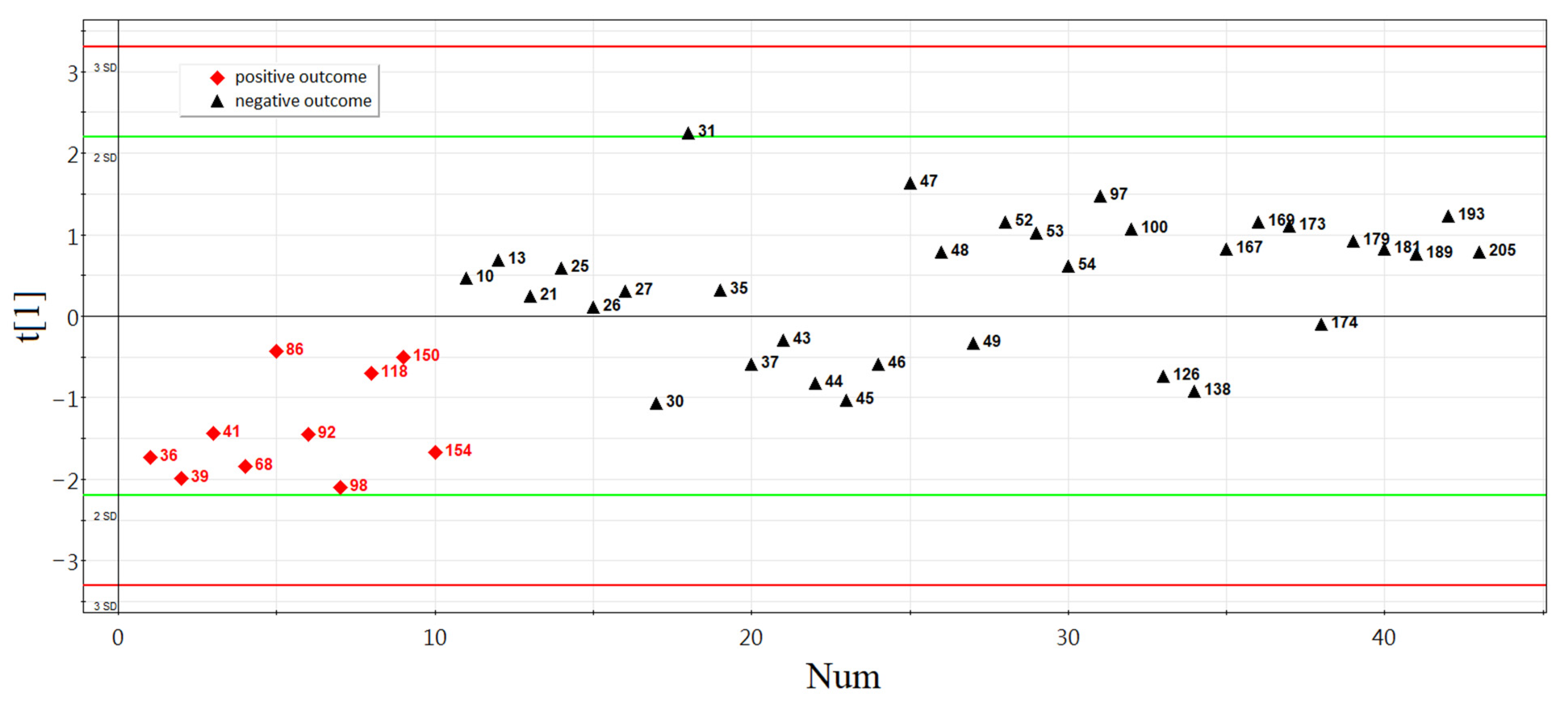
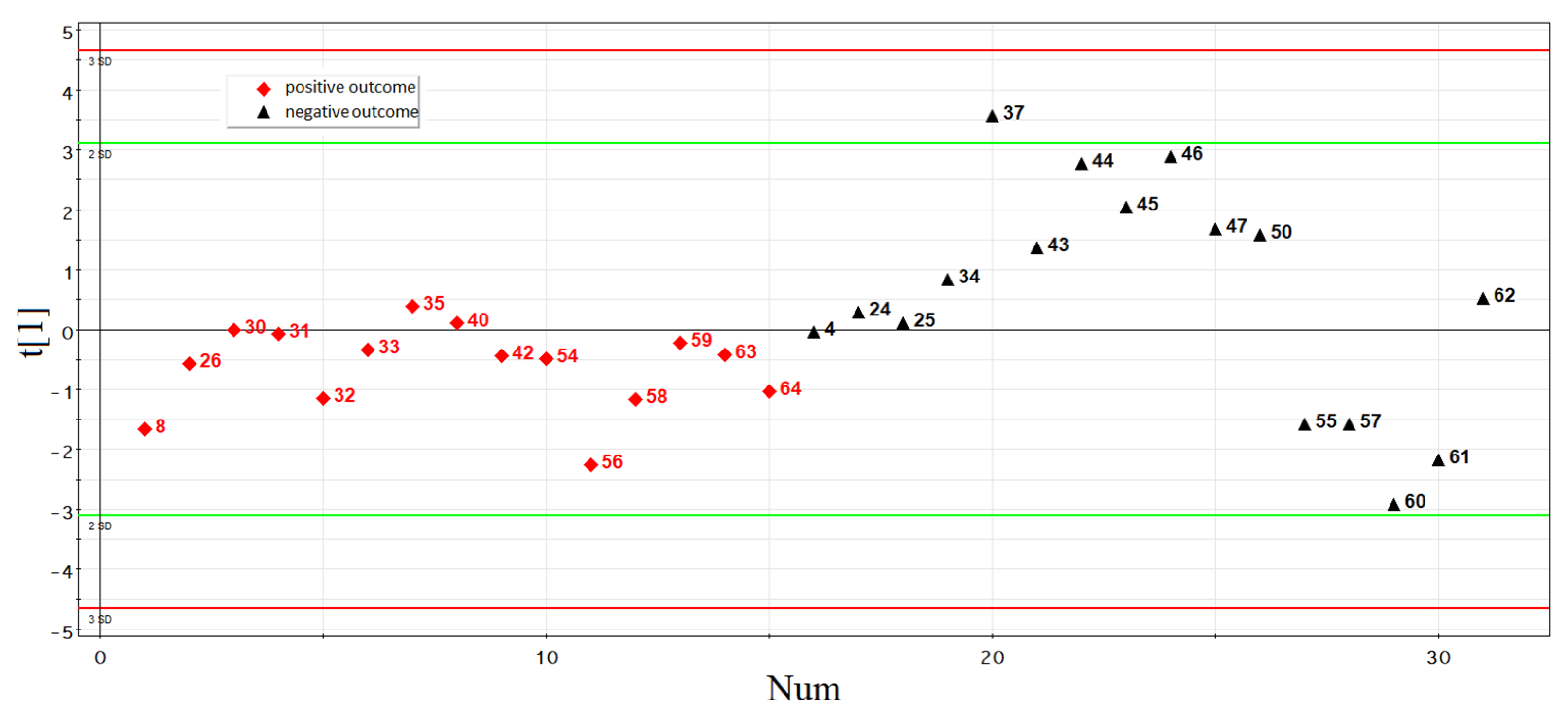
3.2.3. Development of Logistic Regression Model 1
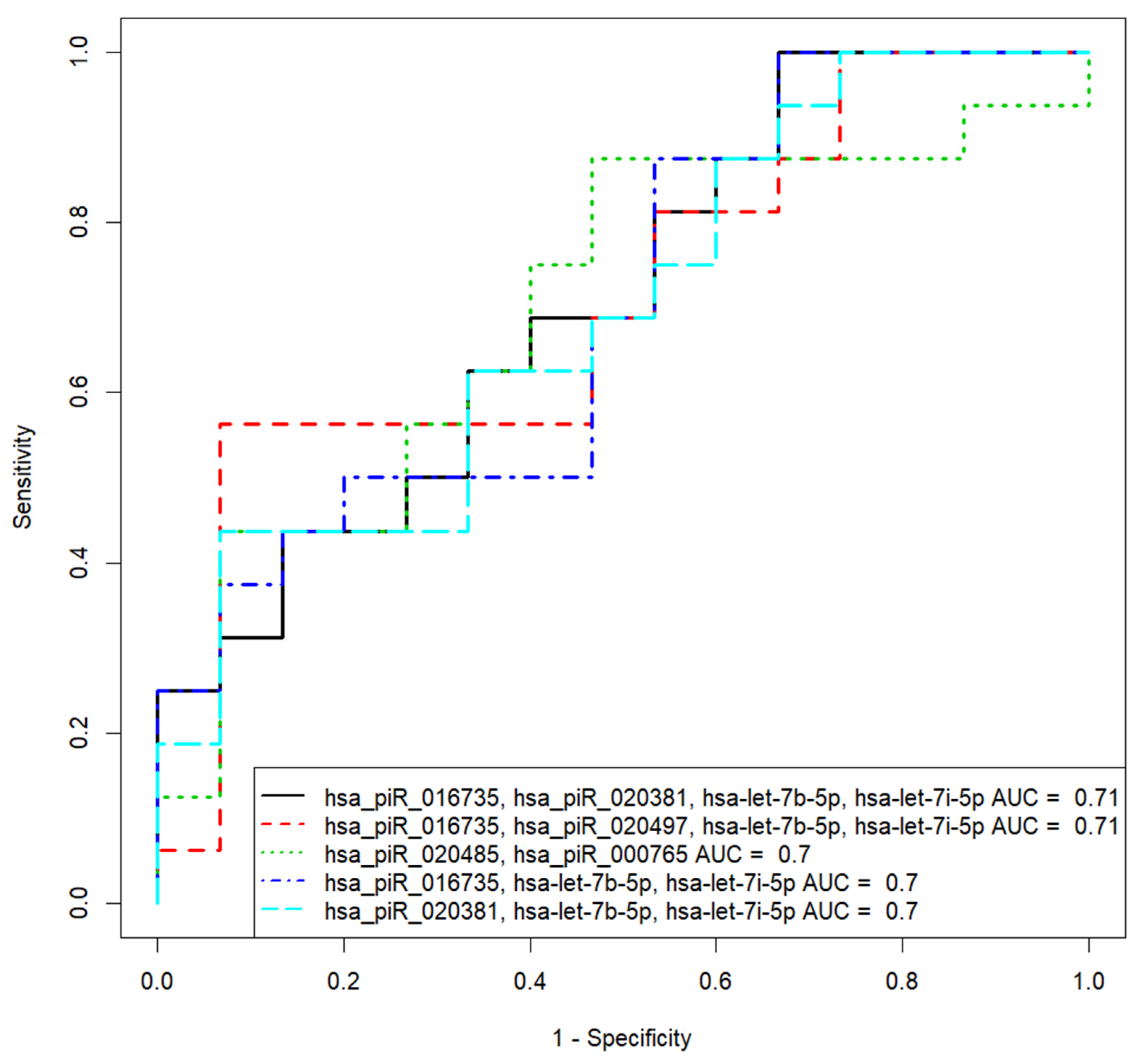
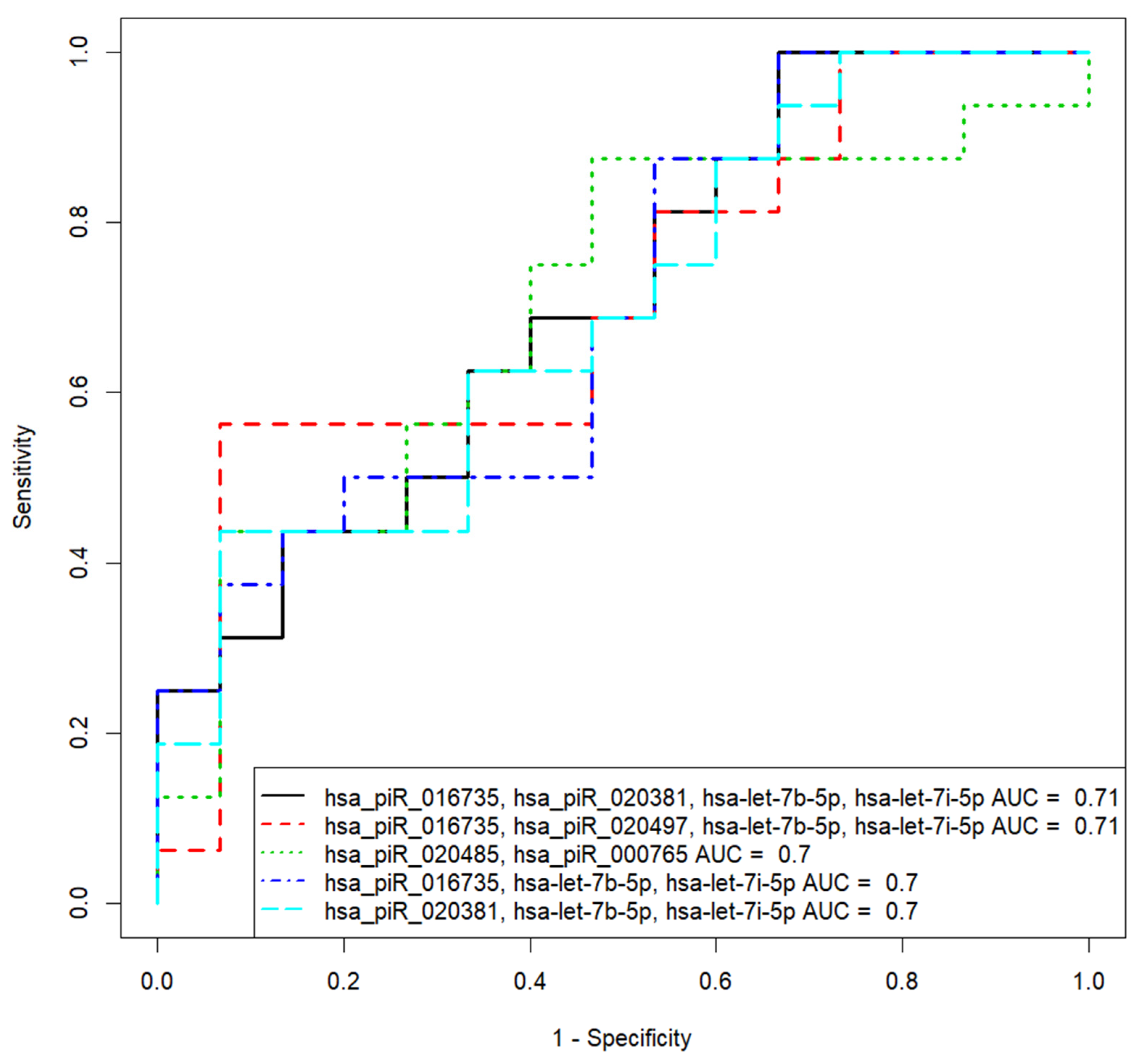
3.3. Verification of the Predictive Value of sncRNA Markers Associated with Implantation Potential of Morula at the Blastocyst Stage
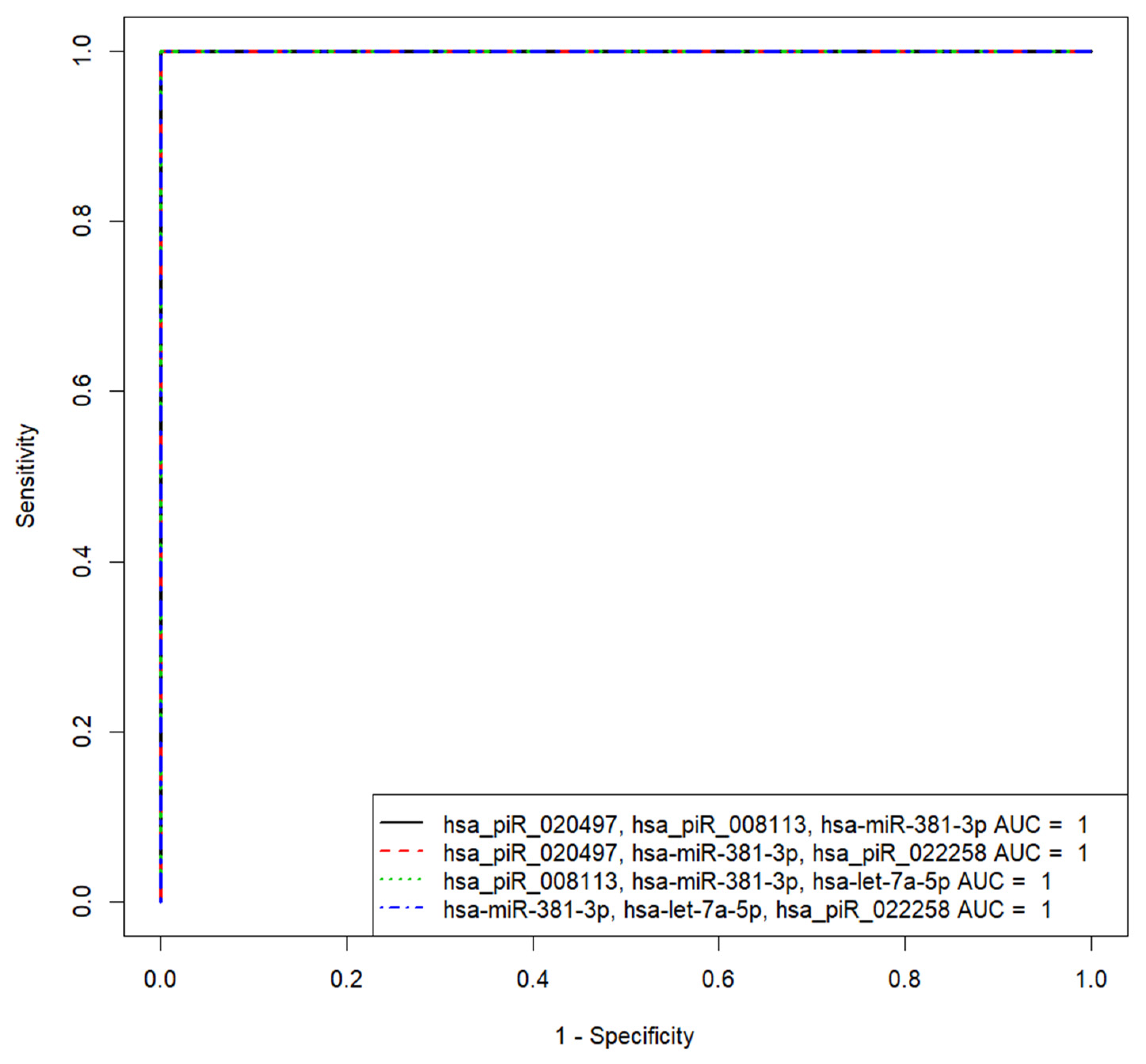
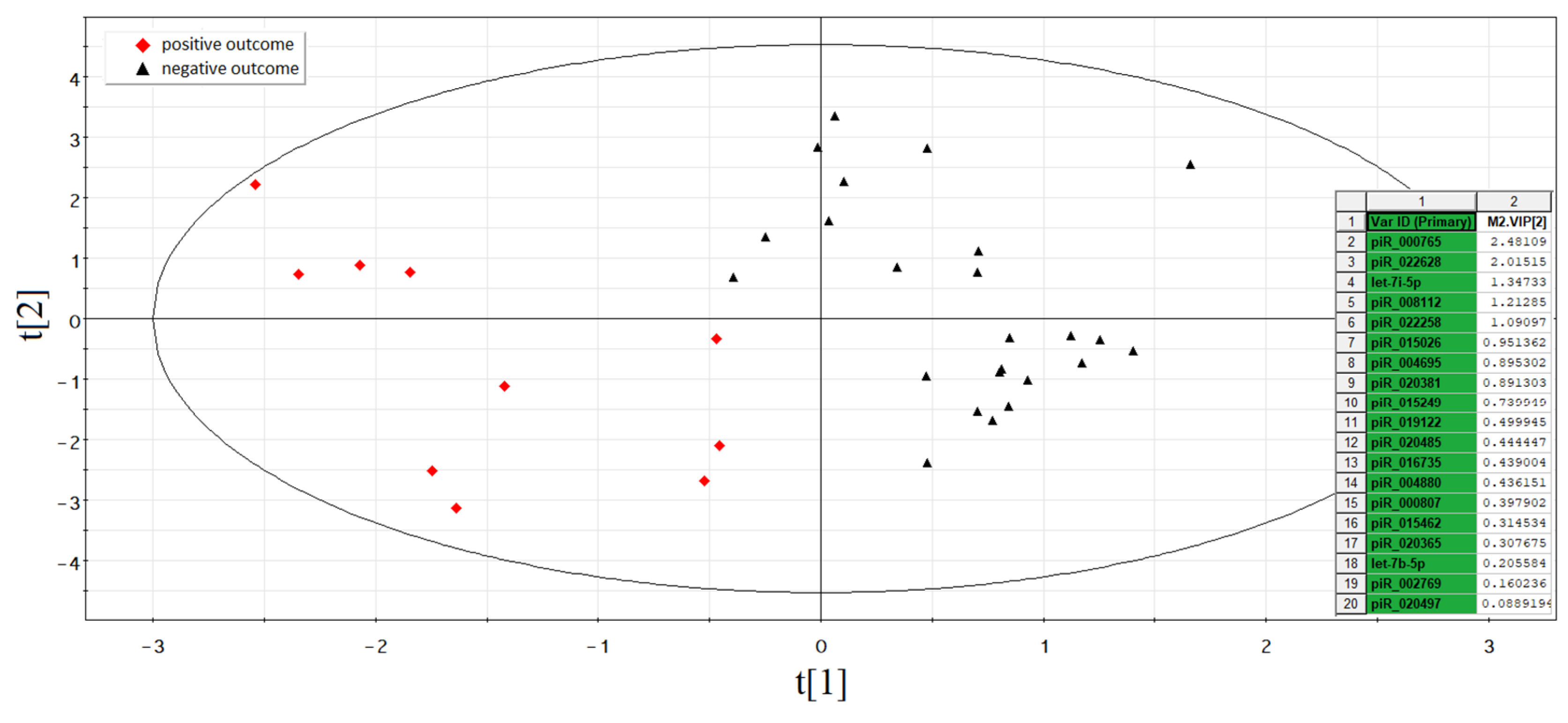
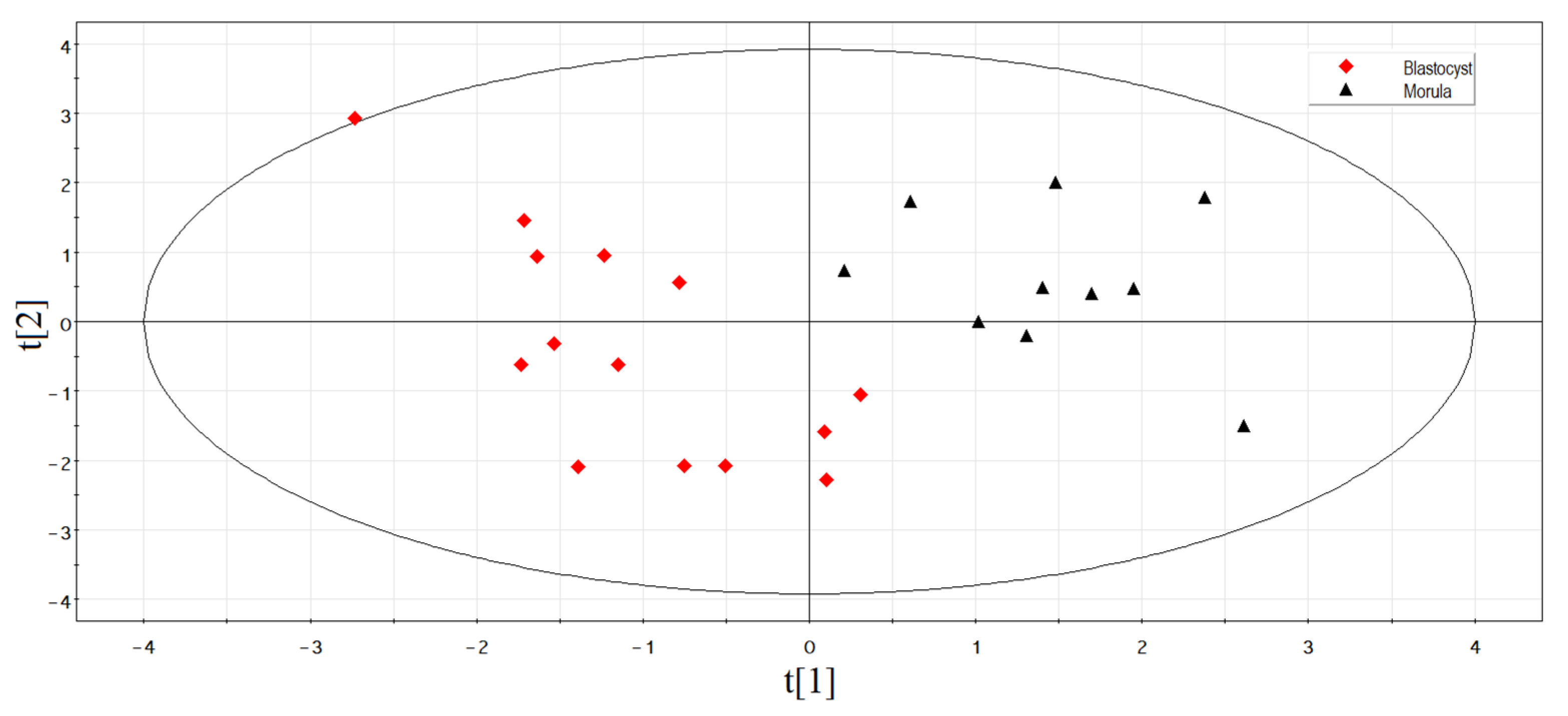
3.4. Identification of sncRNAs Associated with Implantation Potential of Embryo at Blastocyst Stage
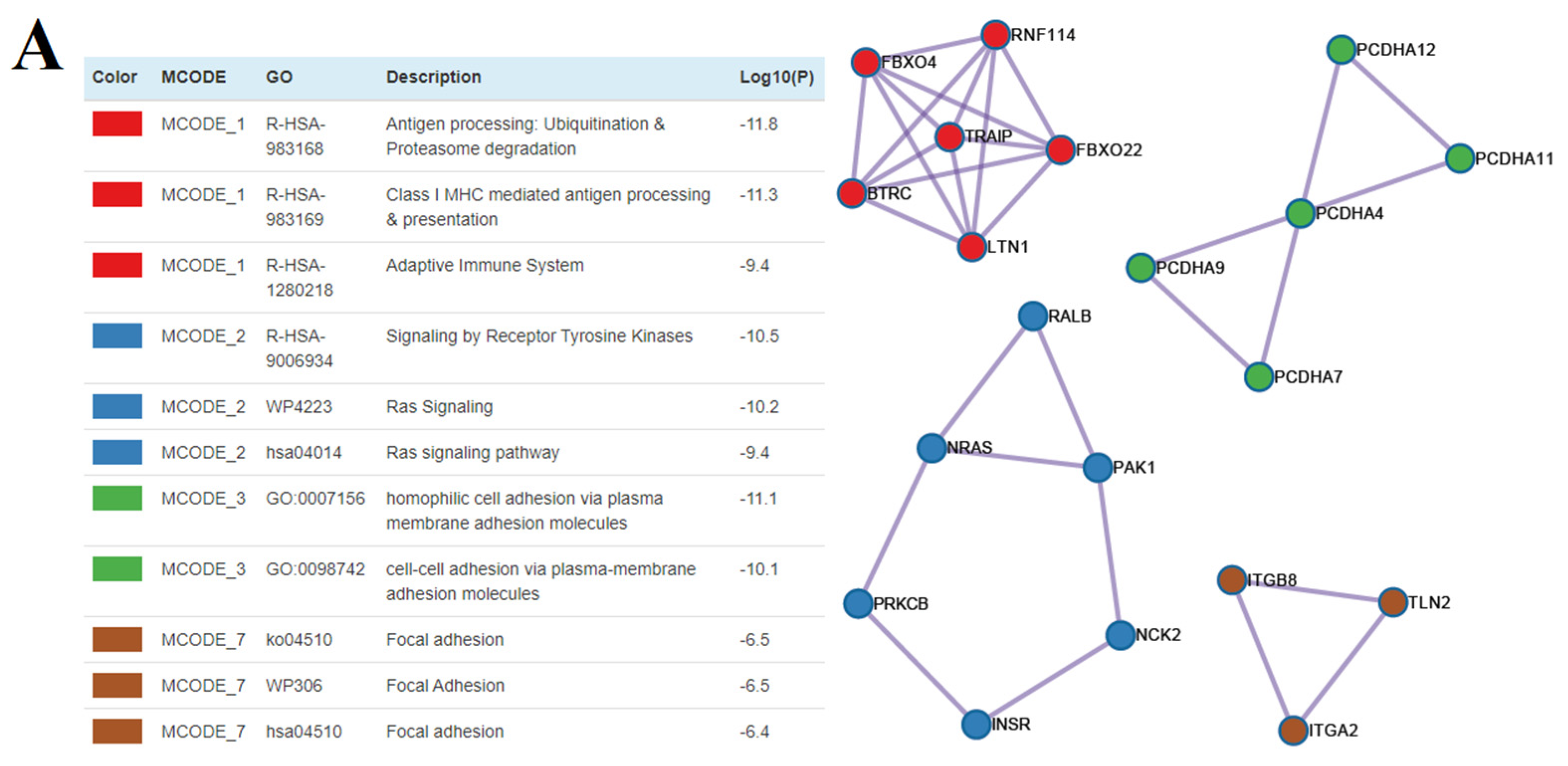
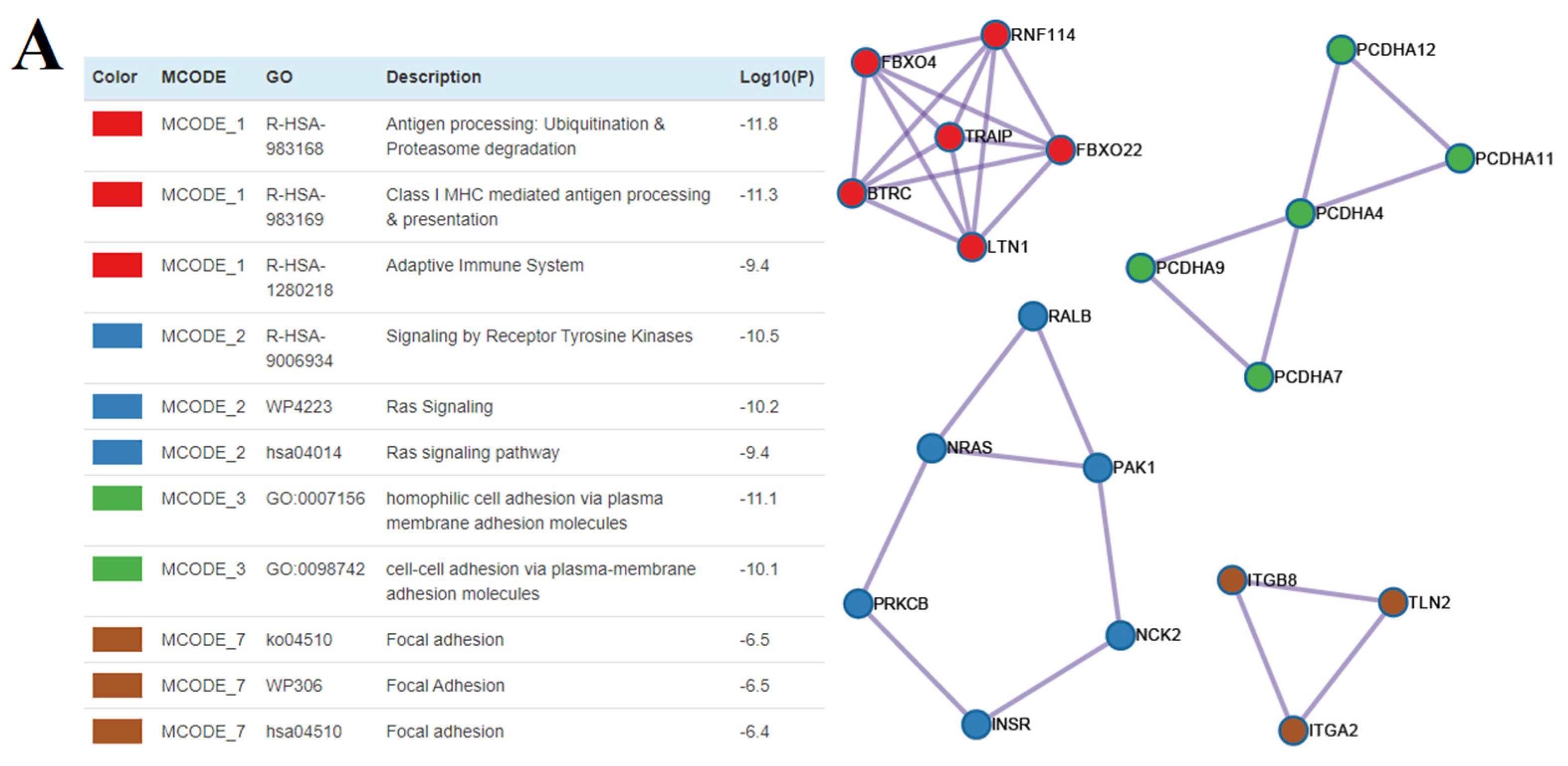
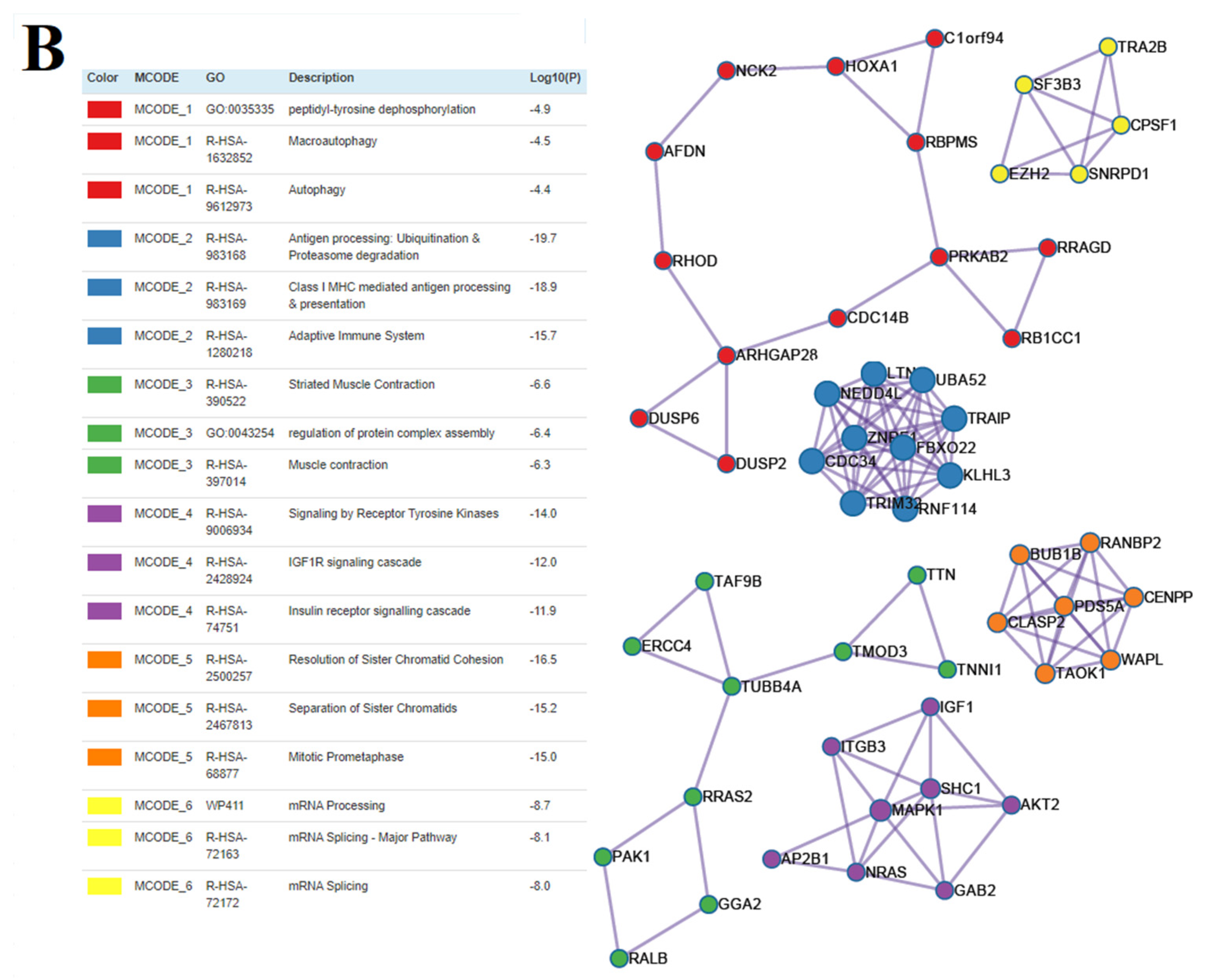
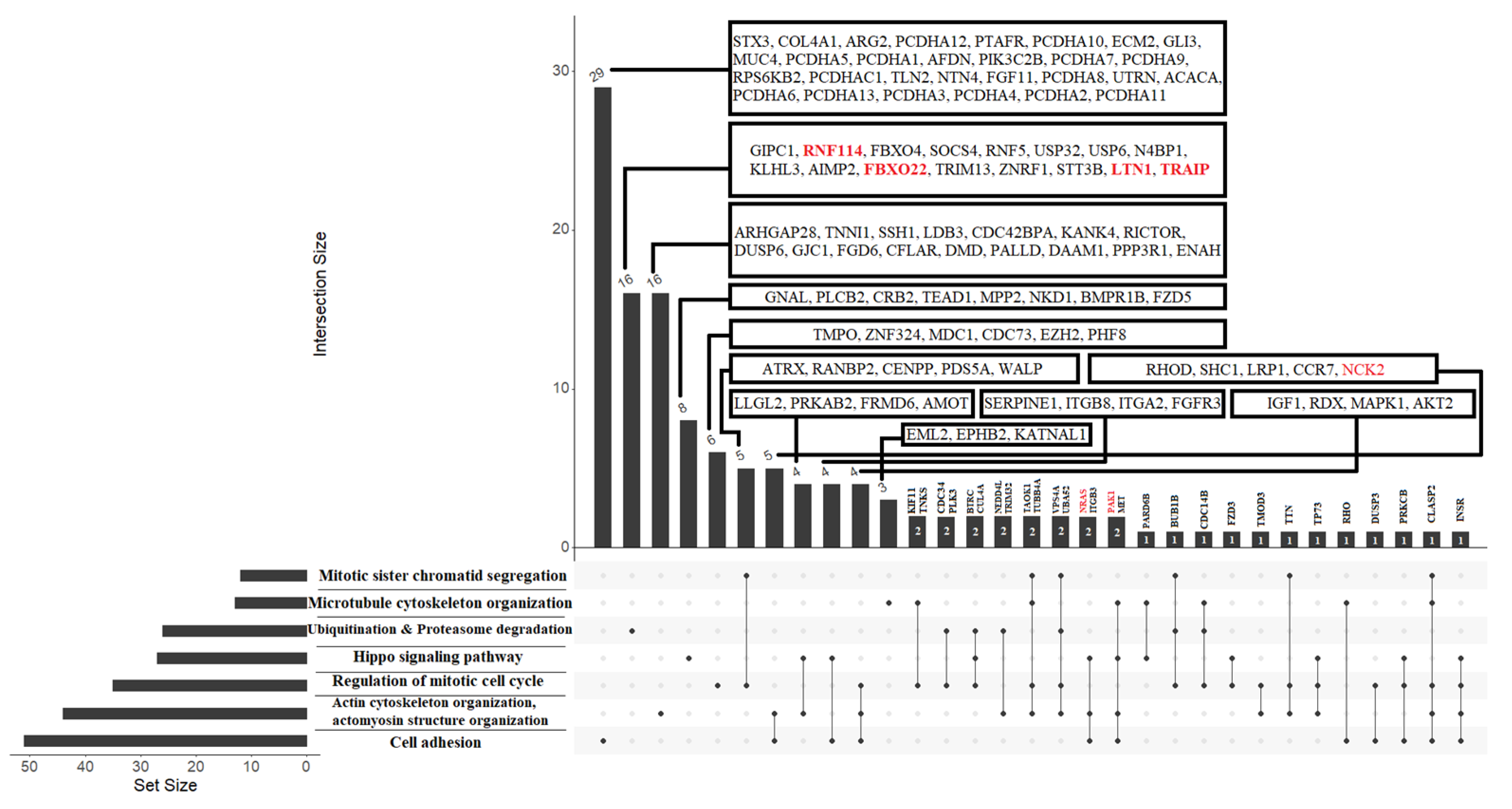
3.5. The Functional Significance of sncRNA Molecules Associated with the Implantation Potential of an Embryo at the Morula and Blastocyst Stages
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Huang, J.Y.J.; Rosenwaks, Z. Assisted reproductive techniques. Methods Mol. Biol. 2014, 1154, 171–231. [Google Scholar]
- Howie, R.; Kay, V. Controlled ovarian stimulation for in-vitro fertilization. Br. J. Hosp. Med. 2018, 79, 194–199. [Google Scholar] [CrossRef]
- Palermo, G.D.; Cohen, J.; Alikani, M.; Adler, A.; Rosenwaks, Z. Intracytoplasmic sperm injection: A novel treatment for all forms of male factor infertility. Fertil. Steril. 1995, 63, 1231–1240. [Google Scholar] [CrossRef]
- Cobo, A.; Castellò, D.; Vallejo, B.; Albert, C.; de los Santos, J.M.; Remohí, J. Outcome of cryotransfer of embryos developed from vitrified oocytes: Double vitrification has no impact on delivery rates. Fertil. Steril. 2013, 99, 1623–1630. [Google Scholar] [CrossRef] [PubMed]
- Meseguer, M.; Herrero, J.; Tejera, A.; Hilligsøe, K.M.; Ramsing, N.B.; Remohí, J. The use of morphokinetics as a predictor of embryo implantation. Hum. Reprod. 2011, 26, 2658–2671. [Google Scholar] [CrossRef] [Green Version]
- Wong, C.C.; Loewke, K.E.; Bossert, N.L.; Behr, B.; De Jonge, C.J.; Baer, T.M.; Reijo Pera, R.A. Non-invasive imaging of human embryos before embryonic genome activation predicts development to the blastocyst stage. Nat. Biotechnol. 2010, 28, 1115–1121. [Google Scholar] [CrossRef]
- Herrero, J.; Meseguer, M. Selection of high potential embryos using time-lapse imaging: The era of morphokinetics. Fertil. Steril. 2013, 99, 1030–1034. [Google Scholar] [CrossRef]
- Scott, K.L.; Hong, K.H.; Scott, R.T.J. Selecting the optimal time to perform biopsy for preimplantation genetic testing. Fertil. Steril. 2013, 100, 608–614. [Google Scholar] [CrossRef]
- Schoolcraft, W.B.; Katz-Jaffe, M.G. Comprehensive chromosome screening of trophectoderm with vitrification facilitates elective single-embryo transfer for infertile women with advanced maternal age. Fertil. Steril. 2013, 100, 615–619. [Google Scholar] [CrossRef]
- Gleicher, N.; Kushnir, V.A.; Barad, D.H. Worldwide decline of IVF birth rates and its probable causes. Hum. Reprod. Open 2019, 2019, hoz017. [Google Scholar] [CrossRef] [PubMed]
- Capalbo, A.; Rienzi, L.; Cimadomo, D.; Maggiulli, R.; Elliott, T.; Wright, G.; Nagy, Z.P.; Ubaldi, F.M. Correlation between standard blastocyst morphology, euploidy and implantation: An observational study in two centers involving 956 screened blastocysts. Hum. Reprod. 2014, 29, 1173–1181. [Google Scholar] [CrossRef] [Green Version]
- Bori, L.; Paya, E.; Alegre, L.; Viloria, T.A.; Remohi, J.A.; Naranjo, V.; Meseguer, M. Novel and conventional embryo parameters as input data for artificial neural networks: An artificial intelligence model applied for prediction of the implantation potential. Fertil. Steril. 2020, 114, 1232–1241. [Google Scholar] [CrossRef] [PubMed]
- Haas, J.; Meriano, J.; Bassil, R.; Barzilay, E.; Zilberberg, E.; Casper, R.F. Developmental potential of slow-developing embryos: Day-5 morulae compared with day-5 cavitating morulae. Fertil. Steril. 2019, 111, 105–111. [Google Scholar] [CrossRef] [Green Version]
- Leese, H.J. Quiet please, do not disturb: A hypothesis of embryo metabolism and viability. Bioessays 2002, 24, 845–849. [Google Scholar] [CrossRef] [PubMed]
- Baumann, C.G.; Morris, D.G.; Sreenan, J.M.; Leese, H.J. The quiet embryo hypothesis: Molecular characteristics favoring viability. Mol. Reprod. Dev. 2007, 74, 1345–1353. [Google Scholar] [CrossRef]
- D’Souza, F.; Uppangala, S.; Asampille, G.; Salian, S.R.; Kalthur, G.; Talevi, R.; Atreya, H.S.; Adiga, S.K. Spent embryo culture medium metabolites are related to the in vitro attachment ability of blastocysts. Sci. Rep. 2018, 8, 17025. [Google Scholar] [CrossRef] [Green Version]
- Cortezzi, S.S.; Cabral, E.C.; Trevisan, M.G.; Ferreira, C.R.; Setti, A.S.; Braga, D.P.d.A.F.; Figueira, R.d.C.S.; Iaconelli, A.J.; Eberlin, M.N.; Borges, E.J. Prediction of embryo implantation potential by mass spectrometry fingerprinting of the culture medium. Reproduction 2013, 145, 453–462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vergouw, C.G.; Kieslinger, D.C.; Kostelijk, E.H.; Botros, L.L.; Schats, R.; Hompes, P.G.; Sakkas, D.; Lambalk, C.B. Day 3 embryo selection by metabolomic profiling of culture medium with near-infrared spectroscopy as an adjunct to morphology: A randomized controlled trial. Hum. Reprod. 2012, 27, 2304–2311. [Google Scholar] [CrossRef] [Green Version]
- Dominguez, F.; Meseguer, M.; Aparicio-Ruiz, B.; Piqueras, P.; Quiñonero, A.; Simón, C. New strategy for diagnosing embryo implantation potential by combining proteomics and time-lapse technologies. Fertil. Steril. 2015, 104, 908–914. [Google Scholar] [CrossRef]
- Nguyen, H.P.T.; Simpson, R.J.; Salamonsen, L.A.; Greening, D.W. Extracellular Vesicles in the Intrauterine Environment: Challenges and Potential Functions. Biol. Reprod. 2016, 95, 109. [Google Scholar] [CrossRef]
- Capalbo, A.; Ubaldi, F.M.; Cimadomo, D.; Noli, L.; Khalaf, Y.; Farcomeni, A.; Ilic, D.; Rienzi, L. MicroRNAs in spent blastocyst culture medium are derived from trophectoderm cells and can be explored for human embryo reproductive competence assessment. Fertil. Steril. 2016, 105, 223–225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Homer, H.; Rice, G.E.; Salomon, C. Review: Embryo- and endometrium-derived exosomes and their potential role in assisted reproductive treatments-liquid biopsies for endometrial receptivity. Placenta 2017, 54, 89–94. [Google Scholar] [CrossRef] [Green Version]
- Gross, N.; Kropp, J.; Khatib, H. MicroRNA Signaling in Embryo Development. Biology 2017, 6, 34. [Google Scholar] [CrossRef] [PubMed]
- Suh, N.; Blelloch, R. Small RNAs in early mammalian development: From gametes to gastrulation. Development 2011, 138, 1653–1661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Timofeeva, A.V.; Chagovets, V.V.; Drapkina, Y.S.; Makarova, N.P.; Kalinina, E.A.; Sukhikh, G.T. Cell-Free, Embryo-Specific sncRNA as a Molecular Biological Bridge between Patient Fertility and IVF Efficiency. Int. J. Mol. Sci. 2019, 20, 2912. [Google Scholar] [CrossRef] [Green Version]
- Timofeeva, A.; Drapkina, Y.; Fedorov, I.; Chagovets, V.; Makarova, N.; Shamina, M.; Kalinina, E.; Sukhikh, G. Small Noncoding RNA Signatures for Determining the Developmental Potential of an Embryo at the Morula Stage. Int. J. Mol. Sci. 2020, 21, 9399. [Google Scholar] [CrossRef]
- Tao, J.; Tamis, R.; Fink, K.; Williams, B.; Nelson-White, T.; Craig, R. The neglected morula/compact stage embryo transfer. Hum. Reprod. 2002, 17, 1513–1518. [Google Scholar] [CrossRef] [Green Version]
- Gardner, D.K. In vitro culture of human blastocyst. In Towards Reproductive Certainty: Infertiliy and Genetics Beyond; CRC Press: Boca Raton, FL, USA, 1999; pp. 378–388. [Google Scholar]
- Gardner, D.K.; Lane, M.; Stevens, J.; Schlenker, T.; Schoolcraft, W.B. Blastocyst score affects implantation and pregnancy outcome: Towards a single blastocyst transfer. Fertil. Steril. 2000, 73, 1155–1158. [Google Scholar] [CrossRef]
- Langmead, B.; Trapnell, C.; Pop, M.; Salzberg, S.L. Ultrafast and memory-efficient alignment of short DNA sequences to the human genome. Genome Biol. 2009, 10, R25. [Google Scholar] [CrossRef] [Green Version]
- Team, R.C. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013; Available online: https://www.r-project.org (accessed on 10 March 2021).
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Team, Rs. RStudio: Integrated Development for R. RStudio. Available online: http://www.rstudio.com/ (accessed on 23 March 2021).
- Panay, N.; Anderson, R.A.; Nappi, R.E.; Vincent, A.J.; Vujovic, S.; Webber, L.; Wolfman, W. Premature ovarian insufficiency: An International Menopause Society White Paper. Climacteric 2020, 23, 426–446. [Google Scholar] [CrossRef]
- Yang, Y.; Zhou, C.; Wang, Y.; Liu, W.; Liu, C.; Wang, L.; Liu, Y.; Shang, Y.; Li, M.; Zhou, S.; et al. The E3 ubiquitin ligase RNF114 and TAB1 degradation are required for maternal-to-zygotic transition. EMBO Rep. 2017, 18, 205–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsukamoto, S.; Kuma, A.; Murakami, M.; Kishi, C.; Yamamoto, A.; Mizushima, N. Autophagy is essential for preimplantation development of mouse embryos. Science 2008, 321, 117–120. [Google Scholar] [CrossRef] [PubMed]
- Lim, H.Y.G.; Plachta, N. Cytoskeletal control of early mammalian development. Nat. Rev. Mol. Cell Biol. 2021, 22, 548–562. [Google Scholar] [CrossRef] [PubMed]
- Mihajlović, A.I.; Bruce, A.W. The first cell-fate decision of mouse preimplantation embryo development: Integrating cell position and polarity. Open Biol. 2017, 7, 170210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simon, D.N.; Wilson, K.L. The nucleoskeleton as a genome-associated dynamic “network of networks”. Nat. Rev. Mol. Cell Biol. 2011, 12, 695–708. [Google Scholar] [CrossRef]
- Nakaya, Y.; Sukowati, E.W.; Sheng, G. Epiblast integrity requires CLASP and Dystroglycan-mediated microtubule anchoring to the basal cortex. J. Cell Biol. 2013, 202, 637–651. [Google Scholar] [CrossRef] [Green Version]
- Patergnani, S.; Marchi, S.; Rimessi, A.; Bonora, M.; Giorgi, C.; Mehta, K.D.; Pinton, P. PRKCB/protein kinase C, beta and the mitochondrial axis as key regulators of autophagy. Autophagy 2013, 9, 1367–1385. [Google Scholar] [CrossRef] [Green Version]
- Xie, K.-M.; Hou, X.-F.; Li, M.-Q.; Li, D.-J. NME1 at the human maternal-fetal interface downregulates titin expression and invasiveness of trophoblast cells via MAPK pathway in early pregnancy. Reproduction 2010, 139, 799–808. [Google Scholar] [CrossRef] [Green Version]
- Nussinov, R.; Zhang, M.; Maloney, R.; Jang, H. Ras isoform-specific expression, chromatin accessibility, and signaling. Biophys. Rev. 2021, 13, 489–505. [Google Scholar] [CrossRef]
- Jo, Y.-J.; Jang, W.-I.; Kim, N.-H.; Namgoong, S. Tropomodulin-3 is essential in asymmetric division during mouse oocyte maturation. Sci. Rep. 2016, 6, 29204. [Google Scholar] [CrossRef] [Green Version]
- Buffone, M.G.; Schindler, K.; Schultz, R.M. Overexpression of CDC14B causes mitotic arrest and inhibits zygotic genome activation in mouse preimplantation embryos. Cell Cycle 2009, 8, 3904–3913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suo, L.; Zhou, Y.X.; Jia, L.L.; Wu, H.B.; Zheng, J.; Lyu, Q.F.; Sun, L.H.; Sun, H.; Kuang, Y.P. Transcriptome profiling of human oocytes experiencing recurrent total fertilization failure. Sci. Rep. 2018, 8, 17890. [Google Scholar] [CrossRef] [PubMed]
- Humphries, J.D.; Chastney, M.R.; Askari, J.A.; Humphries, M.J. Signal transduction via integrin adhesion complexes. Curr. Opin. Cell Biol. 2019, 56, 14–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, Y.-J.; Forbes, K.; Carver, J.; Aplin, J.D. The role of the osteopontin-integrin αvβ3 interaction at implantation: Functional analysis using three different in vitro models. Hum. Reprod. 2014, 29, 739–749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lessey, B.A.; Castelbaum, A.J.; Buck, C.A.; Lei, Y.; Yowell, C.W.; Sun, J. Further characterization of endometrial integrins during the menstrual cycle and in pregnancy. Fertil. Steril. 1994, 62, 497–506. [Google Scholar] [CrossRef]
- Yeo, D.; He, H.; Baldwin, G.S.; Nikfarjam, M. The role of p21-activated kinases in pancreatic cancer. Pancreas 2015, 44, 363–369. [Google Scholar] [CrossRef] [PubMed]
- Bladt, F.; Aippersbach, E.; Gelkop, S.; Strasser, G.A.; Nash, P.; Tafuri, A.; Gertler, F.B.; Pawson, T. The murine Nck SH2/SH3 adaptors are important for the development of mesoderm-derived embryonic structures and for regulating the cellular actin network. Mol. Cell. Biol. 2003, 23, 4586–4597. [Google Scholar] [CrossRef] [Green Version]
- Jacquet, K.; Banerjee, S.L.; Chartier, F.J.M.; Elowe, S.; Bisson, N. Proteomic Analysis of NCK1/2 Adaptors Uncovers Paralog-specific Interactions That Reveal a New Role for NCK2 in Cell Abscission During Cytokinesis. Mol. Cell. Proteom. 2018, 17, 1979–1990. [Google Scholar] [CrossRef] [Green Version]
- Wawro, M.E.; Sobierajska, K.; Ciszewski, W.M.; Wagner, W.; Frontczak, M.; Wieczorek, K.; Niewiarowska, J. Tubulin beta 3 and 4 are involved in the generation of early fibrotic stages. Cell Signal. 2017, 38, 26–38. [Google Scholar] [CrossRef] [PubMed]
- Ott, C.; Nachmias, D.; Adar, S.; Jarnik, M.; Sherman, S.; Birnbaum, R.Y.; Lippincott-Schwartz, J.; Elia, N. VPS4 is a dynamic component of the centrosome that regulates centrosome localization of γ-tubulin, centriolar satellite stability and ciliogenesis. Sci. Rep. 2018, 8, 3353. [Google Scholar] [CrossRef]
- Plouffe, S.W.; Meng, Z.; Lin, K.C.; Lin, B.; Hong, A.W.; Chun, J.V.; Guan, K.-L. Characterization of Hippo Pathway Components by Gene Inactivation. Mol. Cell 2016, 64, 993–1008. [Google Scholar] [CrossRef] [Green Version]
- Feng, C.; Yang, F.; Wang, J. FBXO4 inhibits lung cancer cell survival by targeting Mcl-1 for degradation. Cancer Gene Ther. 2017, 24, 342–347. [Google Scholar] [CrossRef]
- Lian, Z.; Lee, E.K.; Bass, A.J.; Wong, K.K.; Klein-Szanto, A.J.; Rustgi, A.K.; Diehl, J.A. FBXO4 loss facilitates carcinogen induced papilloma development in mice. Cancer Biol. Ther. 2015, 16, 750–755. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.-H.; Choi, M.-Y.; Cui, Y.-H.; Kaushik, N.; Uddin, N.; Yoo, K.-C.; Kim, M.-J.; Lee, S.-J. Regulation of FBXO4-mediated ICAM-1 protein stability in metastatic breast cancer. Oncotarget 2017, 8, 83100–83113. [Google Scholar] [CrossRef] [Green Version]
- Herington, J.L.; Bi, J.; Martin, J.D.; Bany, B.M. Beta-catenin (CTNNB1) in the mouse uterus during decidualization and the potential role of two pathways in regulating its degradation. J. Histochem. Cytochem. Off. J. Histochem. Soc. 2007, 55, 963–974. [Google Scholar] [CrossRef] [PubMed]
- Mao, J.; O’Gorman, C.; Sutovsky, M.; Zigo, M.; Wells, K.D.; Sutovsky, P. Ubiquitin A-52 residue ribosomal protein fusion product 1 (Uba52) is essential for preimplantation embryo development. Biol. Open 2018, 7, bio035717. [Google Scholar] [CrossRef] [Green Version]
- Ding, Y.; Zhang, Y.; Xu, C.; Tao, Q.-H.; Chen, Y.-G. HECT domain-containing E3 ubiquitin ligase NEDD4L negatively regulates Wnt signaling by targeting dishevelled for proteasomal degradation. J. Biol. Chem. 2013, 288, 8289–8298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michael, W.M.; Newport, J. Coupling of mitosis to the completion of S phase through Cdc34-mediated degradation of Wee1. Science 1998, 282, 1886–1889. [Google Scholar] [CrossRef]
- Johmura, Y.; Harris, A.S.; Ohta, T.; Nakanishi, M. FBXO22, an epigenetic multiplayer coordinating senescence, hormone signaling, and metastasis. Cancer Sci. 2020, 111, 2718–2725. [Google Scholar] [CrossRef]
- Ghosh, A.; Shcherbik, N. Cooperativity between the Ribosome-Associated Chaperone Ssb/RAC and the Ubiquitin Ligase Ltn1 in Ubiquitination of Nascent Polypeptides. Int. J. Mol. Sci. 2020, 21, 6815. [Google Scholar] [CrossRef] [PubMed]
- Chapard, C.; Meraldi, P.; Gleich, T.; Bachmann, D.; Hohl, D.; Huber, M. TRAIP is a regulator of the spindle assembly checkpoint. J. Cell Sci. 2014, 127, 5149–5156. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| sncRNA 1 | Accession Number 1 | Nucleotide Sequence of Sense Primer for PCR, 5′-3′ | PCR Primers Annealing Temperature, °C |
|---|---|---|---|
| hsa_piR_001312 | DQ571813 | attggtggttcagtggtagaattctcgcctg | 46.2 |
| hsa_piR_002769 | DQ573726 | tcataagtgggagctaaatgatgagaac | 46.2 |
| hsa_piR_008113 | DQ581032 | tgaggtagtaggttgtatagttttagggtc | 46.2 |
| hsa_piR_016240 | DQ592148 | tgttaaccgaaaggttggtggttcgagcc | 48.9 |
| hsa_piR_016742 | DQ593049 | ccggctagctcagtcggtagagcatgaga | 48.9 |
| hsa_piR_019122 | DQ596252 | gacagagaaaacaaggtggtgaactatgccc | 46.2 |
| hsa_piR_020365 | DQ597975 | ggccgtgatcgtatagtggttagtactctg | 46.2 |
| hsa_piR_000765 | DQ570956 | agcattggtggttcagtggtagaattctcgc | 49 |
| hsa_piR_000807 | DQ571005 | agcctgatgatgtcctcctccagttgccgc | 53 |
| hsa_piR_004695 | DQ576442 | tcctggaccagcctgatgatgtcctcctcc | 57 |
| hsa_piR_004880 | DQ576715 | tccttgtcctggaccagcctgatgatgtcct | 45 |
| hsa_piR_008112 | DQ581031 | tgaggtagtagattgtatagttgtggggtagt | 46.2 |
| hsa_piR_015026 | DQ590548 | tggttcagtggtagaattctcgcctcc | 45 |
| hsa_piR_015249 | DQ590830 | tgtagtcgtggccgagtggttaaggc | 46.2 |
| hsa_piR_015462 | DQ591122 | tgtcctgggccagcctgatgatgtcctcctc | 45 |
| hsa_piR_016735 | DQ593039 | ccgcctgggaataccgggtgctgtaggctta | 50 |
| hsa_piR_017716 | DQ594453 | ttccctggtggtctagtggttaggattcggc | 45 |
| hsa_piR_020381 | DQ597997 | ggcgggagtaactatgactctcttaaggta | 53 |
| hsa_piR_020485 | DQ598159 | ggggatgtagctcagtggtagagcgcatgct | 53 |
| hsa_piR_020497 | DQ598177 | tgtagctcagtggtagagcgcgtgct | 45 |
| hsa_piR_022258 | DQ600471 | tactacctgattggtcgggtgtgagc | 48.9 |
| hsa_piR_022628 | DQ600952 | tagagcatgagactcttaatctcagggtcgtg | 48.9 |
| hsa_piR_020388 | DQ598008 | ggctcgttggtctaggggtatgattctcgg | 45 |
| hsa-miR-381-3p | MIMAT0000736 | tatacaagggcaagctctctgt | 52.7 |
| hsa-let-7a-5p | MIMAT0000062 | Hs_let-7a_2 miScript Primer Assay, MS00031220 | 55 |
| hsa-let-7b-5p | MIMAT0000063 | Hs_let-7b_1 miScript Primer Assay, MS00003122 | 55 |
| hsa-let-7i-5p | MIMAT0000415 | Hs_let-7i_1 miScript Primer Assay, MS00003157 | 55 |
| Clinical Data and Parameters of the IVF/ICSI Protocol | Negative Outcome (n = 85) | Positive Outcome (n = 25) | Mann–Whitney Test, p 1 |
|---|---|---|---|
| Me (Q1;Q3) 1 | |||
| Age of Female | 32.5 (30;35) | 31 (30;32.75) | 0.112915 |
| Menarche, age | 13 (13;14) | 13 (13;14) | 0.941626 |
| Duration of menstrual bleeding, days | 5 (4;5) | 4 (4;5) | 0.195459 |
| The length of the menstrual cycle, days | 28 (28;30) | 28 (28;29.75) | 0.256809 |
| Prolactin (120.0–500.0, mIU/L) | 301.2 (198.55;352.75) | 284.35 (194.25;337.57) | 0.473241 |
| 17-OH-progesterone (0.3–3.0, nmole/L) | 1.58 (1.1;1.96) | 1.3 (1.22;1.65) | 0.538915 |
| Dehydroepiandrosterone sulphate (0.9–11.7, µmole/L) | 5.2 (3.1;7.6) | 3.45 (2.45;6.8) | 0.283792 |
| Cortisol (200.0–550.0, nmole/L) | 315 (245.05;380.5) | 306.15 (259.55;373.3) | 0.901587 |
| Testosterone (0.5–2.5, nmole/L) | 1.61 (1.12;2) | 1.15 (0.74;1.67) | 0.017549 |
| Anti-müllerian hormone (1.0–10.6, ng/mL) | 2.21 (1.52;3.1) | 3.05 (2.35;4.33) | 0.007737 |
| Thyroid-stimulating hormone (0.4–3.5, mIU/L) | 1.77 (1.15;2.49) | 1.27 (0.98;2.14) | 0.182973 |
| Free thyroxine (10.0–25.0, pmole/L) | 15.1 (12.22;18) | 14.75 (12.06;18.3) | 0.950685 |
| Luteinizing hormone (2.3–15.0, IU/L) | 5.9 (4.75;7.08) | 4.8 (4.15;5.74) | 0.036986 |
| Progesterone, luteal phase of the menstrual cycle (16.0–95.0, nmole/L) | 27.6 (19.02;43) | 31.58 (24.05;47) | 0.251808 |
| Follicle-stimulating hormone (2.0–10.0, IU/L) | 7.55 (6.53;8.95) | 6.41 (6.18;8.3) | 0.094989 |
| Estradiol (150.0–450.0, pmole/L) | 154.5 (100.05;206.01) | 165.75 (145.25;230) | 0.32861 |
| Gonadotropin Dosage, IU | 1725 (1293.75;1950) | 1425 (1143.75;1762.5) | 0.103318 |
| Stimulation Duration, days | 9 (8;10) | 9 (8.25;10) | 0.406869 |
| Number of follicles on the day of administration of the trigger of the final oocyte maturation | 8.5 (5;11) | 8.5 (5.25;10) | 0.986806 |
| Number of OCCs | 7 (4.25;12) | 8 (4.25;10.75) | 0.820127 |
| Number of Metaphase II (MII) Oocytes | 6 (3;9.75) | 6 (4;7.75) | 0.842485 |
| Age of Male | 34 (32;38) | 33 (31;34.75) | 0.112239 |
| Sperm Concentration, Million per Milliliter | 60 (31.25;107) | 61.5 (41;91) | 0.944129 |
| Morphologically Normal Spermatozoa, % | 2 (2;3) | 2 (1.25;3) | 0.582895 |
| Sperm with Progressive Motility, % | 56 (44;67.75) | 60 (37.5;67) | 0.904815 |
| Number of two pronuclei embryos | 5 (3;8) | 5.5 (3.25;7.5) | 0.765231 |
| Blastocyst number | 1 (1;3) | 1 (1;2.75) | 0.606454 |
| Duration of infertility | 4 (2;6) | 3 (2;5) | 0.302846 |
| Number of IVF attempts in history | 0 (0;1) | 0 (0;0.75) | 0.189145 |
| sncRNA | Spent Culture Medium from Morula without Potential for Implantation 1 | Spent Culture Medium from Morula with Potential for Implantation 1 | Culture Medium without Embryo 1 | Mann–Whitney Test (p-Value) | |||||
|---|---|---|---|---|---|---|---|---|---|
| #179 | #167 | #193 | #173 | #150 | #118 | #154 | #207 | ||
| hsa_piR_022628 | 7 | 21 | 63 | 21 | 77 | 119 | 133 | 14 | 0.00484937 |
| hsa_piR_015249 | 75 | 160 | 135 | 200 | 240 | 245 | 310 | 210 | 0.009863152 |
| hsa_piR_016735 | 23 | 107 | 171 | 124 | 205 | 232 | 265 | 204 | 0.011349613 |
| hsa_piR_004880 | 139 | 277 | 572 | 291 | 616 | 651 | 839 | 549 | 0.012957723 |
| hsa_piR_004695 | 139 | 280 | 576 | 295 | 622 | 651 | 844 | 551 | 0.013187947 |
| hsa_piR_015462 | 278 | 560 | 1152 | 590 | 1244 | 1302 | 1688 | 1102 | 0.013187947 |
| hsa_piR_000807 | 278 | 560 | 1152 | 590 | 1244 | 1300 | 1684 | 1102 | 0.01323007 |
| hsa_piR_022258 | 0 | 0 | 0 | 0 | 10 | 68 | 68 | 0 | 0.014900377 |
| hsa_piR_008112 | 2 | 0 | 8 | 2 | 9 | 7 | 9 | 0 | 0.026879189 |
| hsa_piR_019122 | 4 | 5 | 24 | 13 | 28 | 23 | 54 | 35 | 0.030210112 |
| hsa-let-7i-5p | 39 | 2 | 0 | 25 | 151 | 23 | 101 | 18 | 0.036138954 |
| hsa_piR_020365 | 9 | 15 | 12 | 27 | 19 | 53 | 40 | 9 | 0.036254562 |
| has-let-7b-5p | 37 | 2 | 7 | 33 | 160 | 29 | 95 | 17 | 0.037297033 |
| hsa_piR_015026 | 20 | 60 | 65 | 55 | 90 | 65 | 140 | 15 | 0.039937759 |
| hsa_piR_020381 | 1122 | 6942 | 5946 | 7146 | 9432 | 7413 | 14,019 | 5532 | 0.042809887 |
| hsa_piR_000765 | 540 | 432 | 300 | 565 | 1146 | 1248 | 4014 | 240 | 0.043505506 |
| hsa_piR_020497 | 170 | 122 | 189 | 140 | 246 | 165 | 223 | 296 | 0.045522073 |
| hsa_piR_002769 | 0 | 0 | 0 | 0 | 0 | 82 | 164 | 0 | 0.04660816 |
| hsa_piR_020388 | 80 | 41 | 64 | 53 | 86 | 100 | 54 | 83 | 0.115425375 |
| hsa_piR_020485 | 483 | 210 | 546 | 357 | 364 | 308 | 413 | 532 | 0.349984126 |
| Biomarker | AUC | Sp, % | Se, % | Cutoff | i | K |
|---|---|---|---|---|---|---|
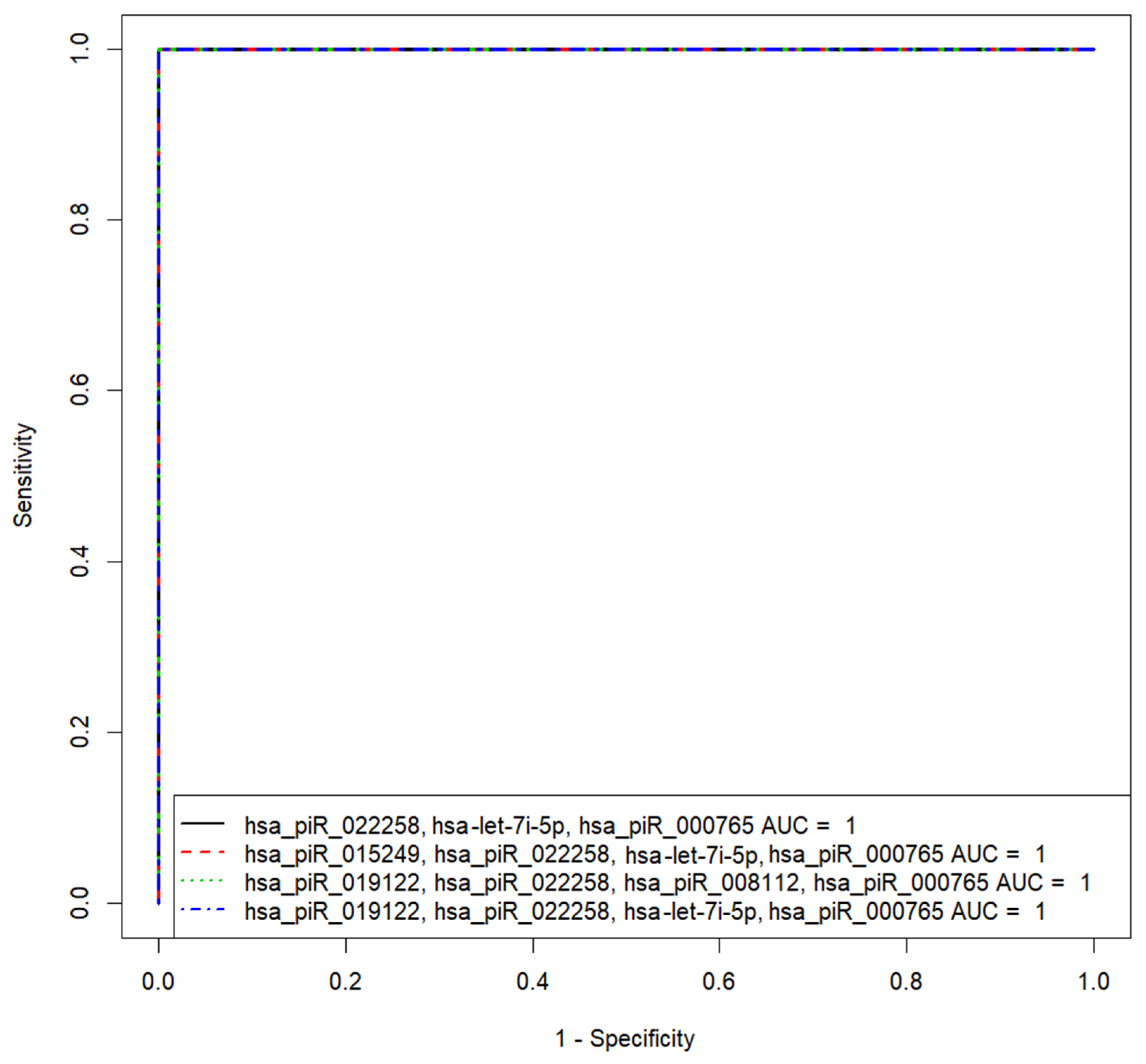
| “hsa_piR_022258, hsa-let-7i-5p, hsa_piR_000765” | 1 | 100 | 100 | 0.5 | 67.9 | −1248.05 −1127.34 −834.25 |
| “hsa_piR_015249, hsa_piR_022258, hsa-let-7i-5p, hsa_piR_000765” | 1 | 100 | 100 | 0.5 | −15.05 | 63.06 −211.33 −178.26 −146.78 |
| “hsa_piR_019122, hsa_piR_022258, hsa_piR_008112, hsa_piR_000765” | 1 | 100 | 100 | 0.5 | −280.49 | −26.33 −191.08 295.17 −107.57 |
| “hsa_piR_019122, hsa_piR_022258, hsa-let-7i-5p, hsa_piR_000765” | 1 | 100 | 100 | 0.5 | 12.02 | 31.37 −154.67 −92.71 −99.38 |
| sncRNA | sncRNA Read Count | Fold Changes in the sncRNA Expression Level | |||||
|---|---|---|---|---|---|---|---|
| #4 | #34 | #24 | #8 | #4/#8 | #34/#8 | #24/#8 | |
| hsa-let-7a-5p | 3835 | 443 | 1541 | 7783 | 0.493 | 0.057 | 0.198 |
| hsa-let-7b-5p | 20737 | 254 | 1834 | 26297 | 0.789 | 0.010 | 0.070 |
| hsa-let-7i-5p | 3465 | 90 | 1055 | 4072 | 0.851 | 0.022 | 0.259 |
| hsa-miR-381-3p | 120 | 1 | 83 | 475 | 0.253 | 0.002 | 0.175 |
| hsa_piR_008113 | 173 | 153 | 93 | 652 | 0.265 | 0.235 | 0.143 |
| hsa_piR_001312 | 486 | 486 | 549 | 2151 | 0.226 | 0.226 | 0.255 |
| hsa_piR_000765 | 8850 | 7248 | 2004 | 15162 | 0.584 | 0.478 | 0.132 |
| hsa_piR_016735 | 26 | 118 | 81 | 347 | 0.075 | 0.340 | 0.233 |
| hsa_piR_015249 | 175 | 625 | 580 | 280 | 0.625 | 2.232 | 2.071 |
| hsa_piR_020497 | 163 | 596 | 562 | 181 | 0.901 | 3.293 | 3.105 |
| hsa_piR_016240 | 54 | 126 | 108 | 1 | 54.000 | 126.000 | 108.000 |
| hsa_piR_016742 | 65 | 120 | 150 | 30 | 2.167 | 4.000 | 5.000 |
| hsa_piR_020485 | 2429 | 1841 | 1876 | 952 | 2.551 | 1.934 | 1.971 |
| hsa_piR_020388 | 448 | 499 | 546 | 386 | 1.161 | 1.293 | 1.415 |
| Biomarker | AUC | Sp, % | Se, % | Cutoff | i | K |
|---|---|---|---|---|---|---|
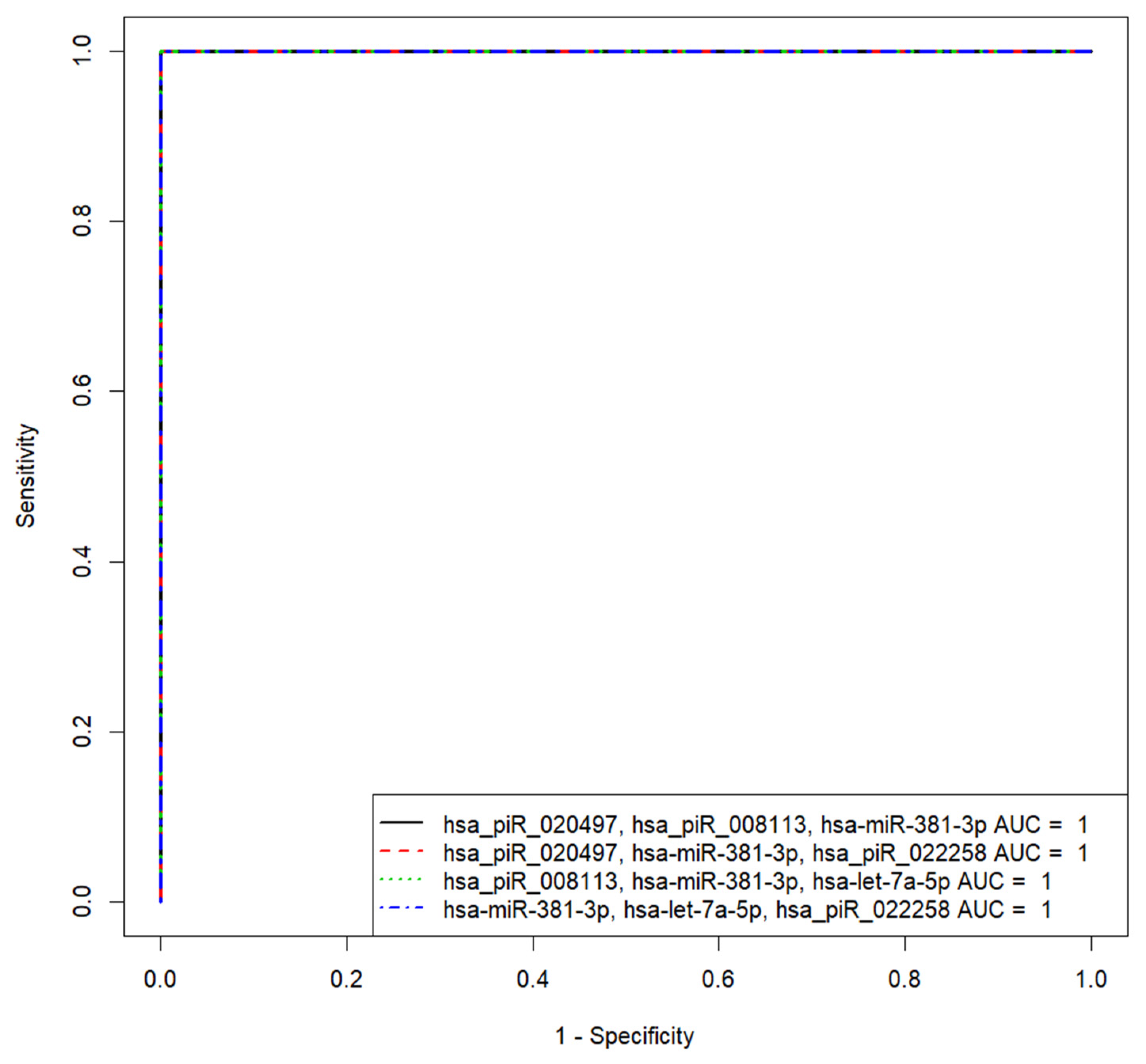
| «hsa_piR_020497, hsa_piR_008113, hsa-miR-381-3p» | 1 | 100 | 100 | 0.5 | 1860.371618 | −1999.768215, 3345.749357, −98.52044368 |
| «hsa_piR_020497, hsa-miR-381-3p, hsa_piR_022258» | 1 | 100 | 100 | 0.5 | 1860.371618 | −1999.768215, −98.52044368, −3345.749357 |
| «hsa_piR_008113, hsa-miR-381-3p, hsa-let-7a-5p» | 1 | 100 | 100 | 0.5 | −6233.293319 | 6761.631563, −516.0692614, −369.5848655 |
| «hsa-miR-381-3p, hsa-let-7a-5p, hsa_piR_022258» | 1 | 100 | 100 | 0.5 | −6233.293319 | −516.0692614, −369.5848655, −6761.631563 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Timofeeva, A.V.; Fedorov, I.S.; Shamina, M.A.; Chagovets, V.V.; Makarova, N.P.; Kalinina, E.A.; Nazarenko, T.A.; Sukhikh, G.T. Clinical Relevance of Secreted Small Noncoding RNAs in an Embryo Implantation Potential Prediction at Morula and Blastocyst Development Stages. Life 2021, 11, 1328. https://doi.org/10.3390/life11121328
Timofeeva AV, Fedorov IS, Shamina MA, Chagovets VV, Makarova NP, Kalinina EA, Nazarenko TA, Sukhikh GT. Clinical Relevance of Secreted Small Noncoding RNAs in an Embryo Implantation Potential Prediction at Morula and Blastocyst Development Stages. Life. 2021; 11(12):1328. https://doi.org/10.3390/life11121328
Chicago/Turabian StyleTimofeeva, Angelika V., Ivan S. Fedorov, Maria A. Shamina, Vitaliy V. Chagovets, Nataliya P. Makarova, Elena A. Kalinina, Tatiana A. Nazarenko, and Gennady T. Sukhikh. 2021. "Clinical Relevance of Secreted Small Noncoding RNAs in an Embryo Implantation Potential Prediction at Morula and Blastocyst Development Stages" Life 11, no. 12: 1328. https://doi.org/10.3390/life11121328
APA StyleTimofeeva, A. V., Fedorov, I. S., Shamina, M. A., Chagovets, V. V., Makarova, N. P., Kalinina, E. A., Nazarenko, T. A., & Sukhikh, G. T. (2021). Clinical Relevance of Secreted Small Noncoding RNAs in an Embryo Implantation Potential Prediction at Morula and Blastocyst Development Stages. Life, 11(12), 1328. https://doi.org/10.3390/life11121328