
Comparative Genome Analysis of Streptococcus suis Serotype 9 Isolates from China, The Netherland, and the U.K.

 and
and 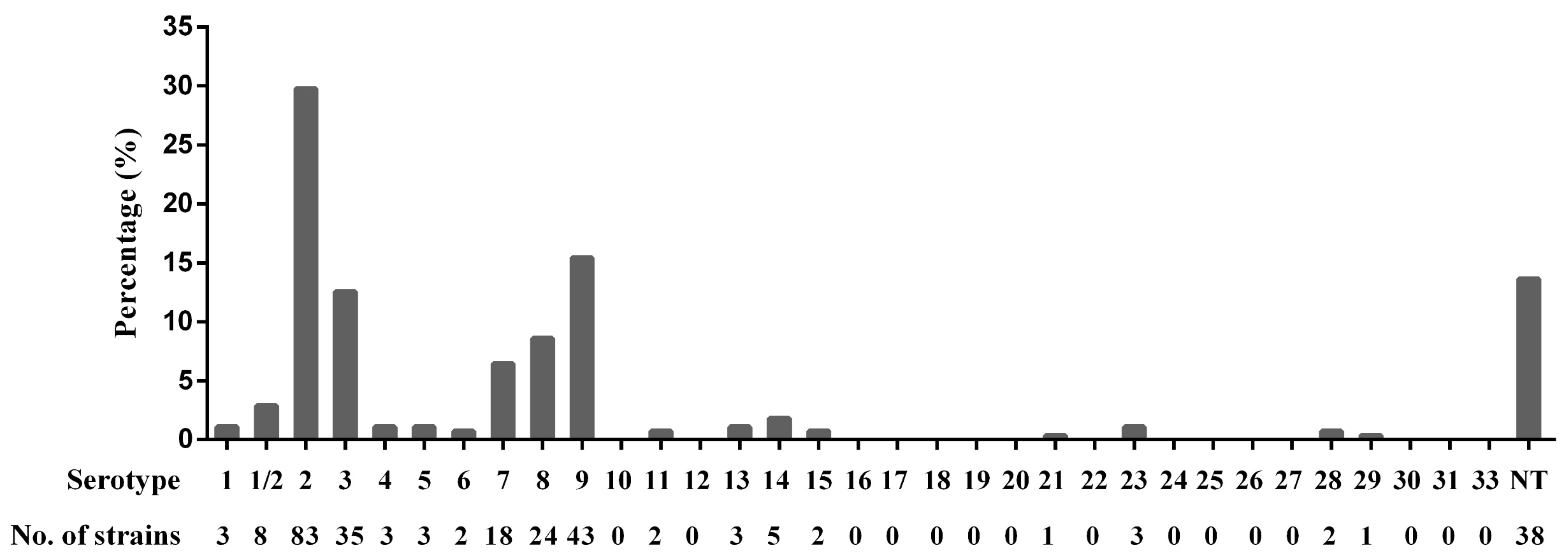{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Isolation and Identification
2.2. Serotyping of S. suis
2.3. Genome Sequencing Analysis
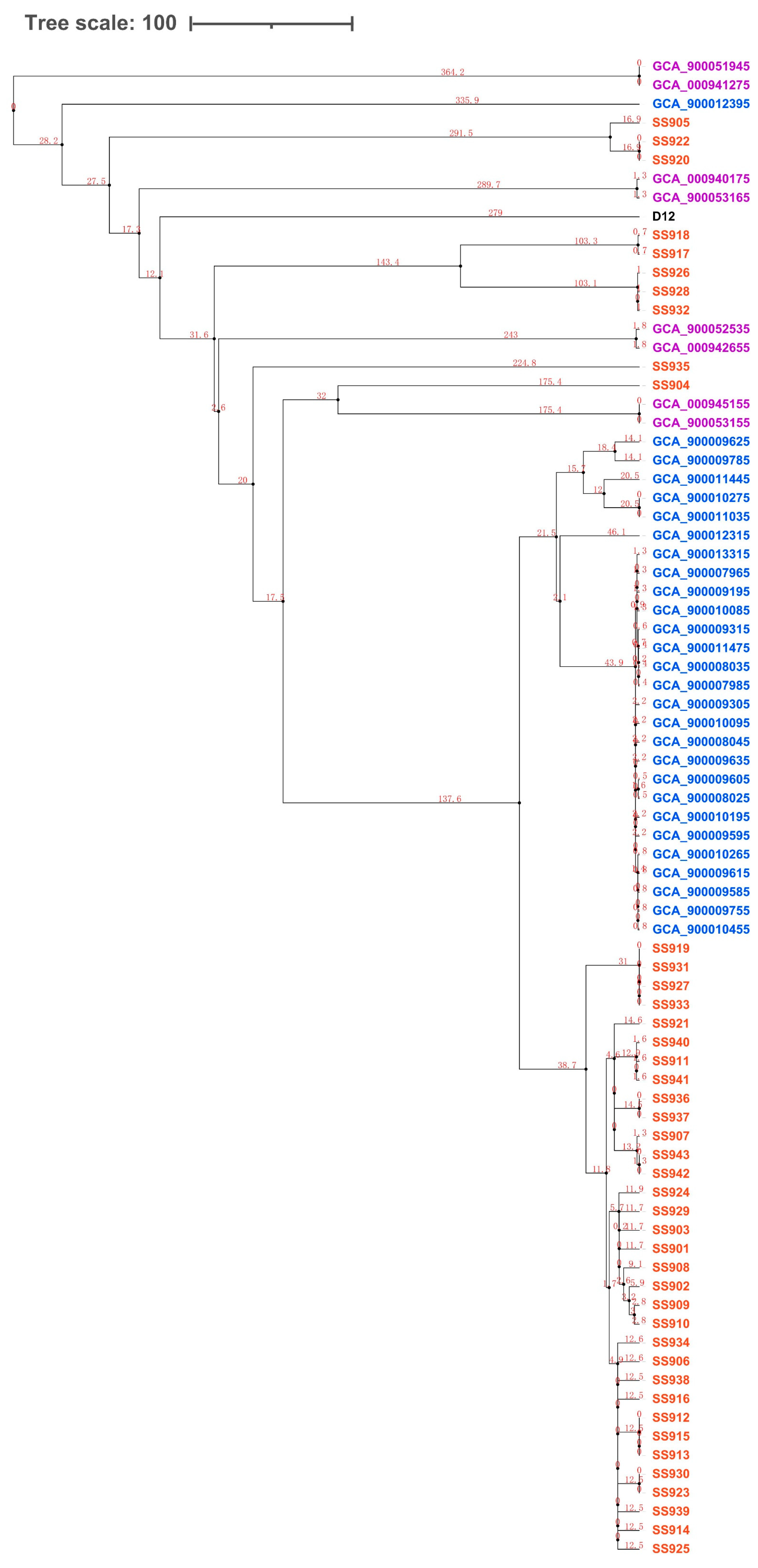
2.4. Phylogenetic Analysis
2.5. Comparative Analysis of Virulence Potential
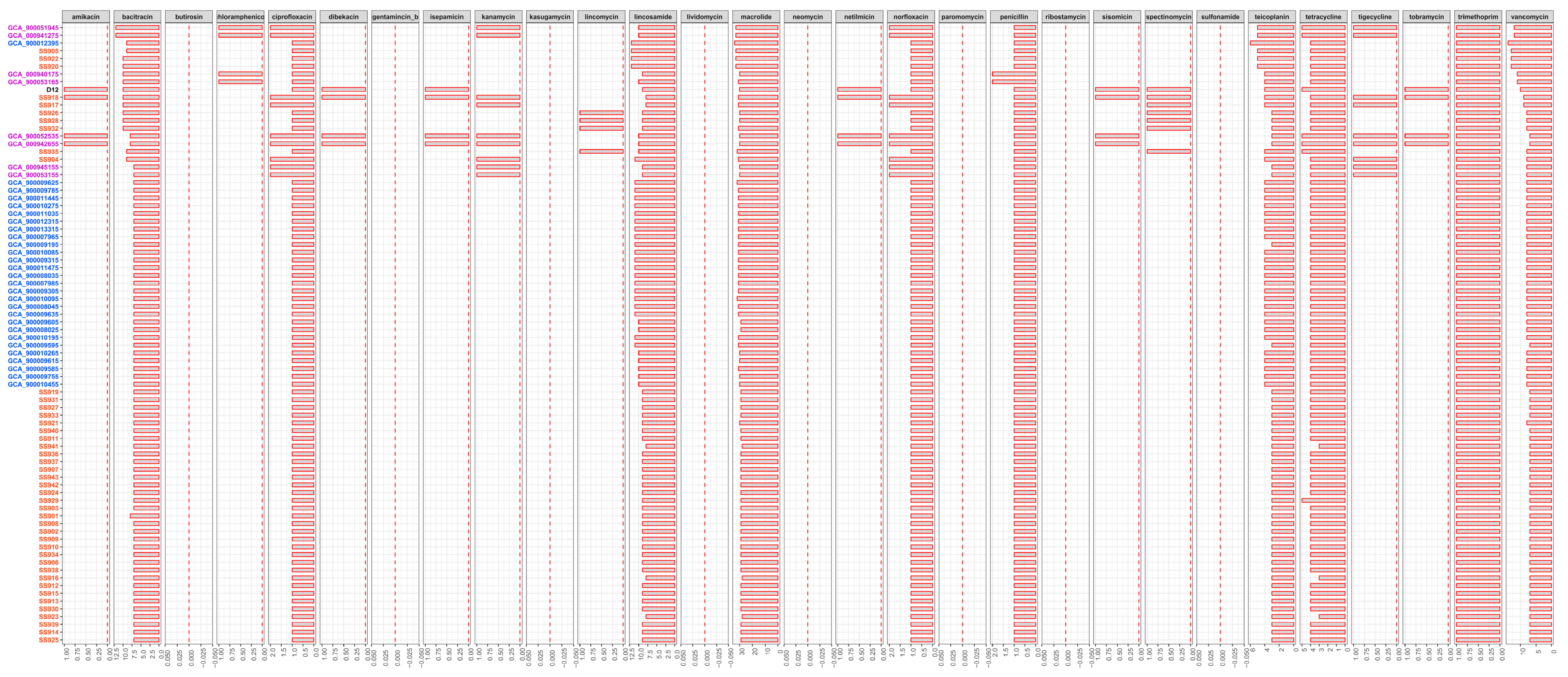
2.6. Comparative Analysis of Drug Resistance Potential
2.7. Nucleotide Sequence Accession Number
3. Results
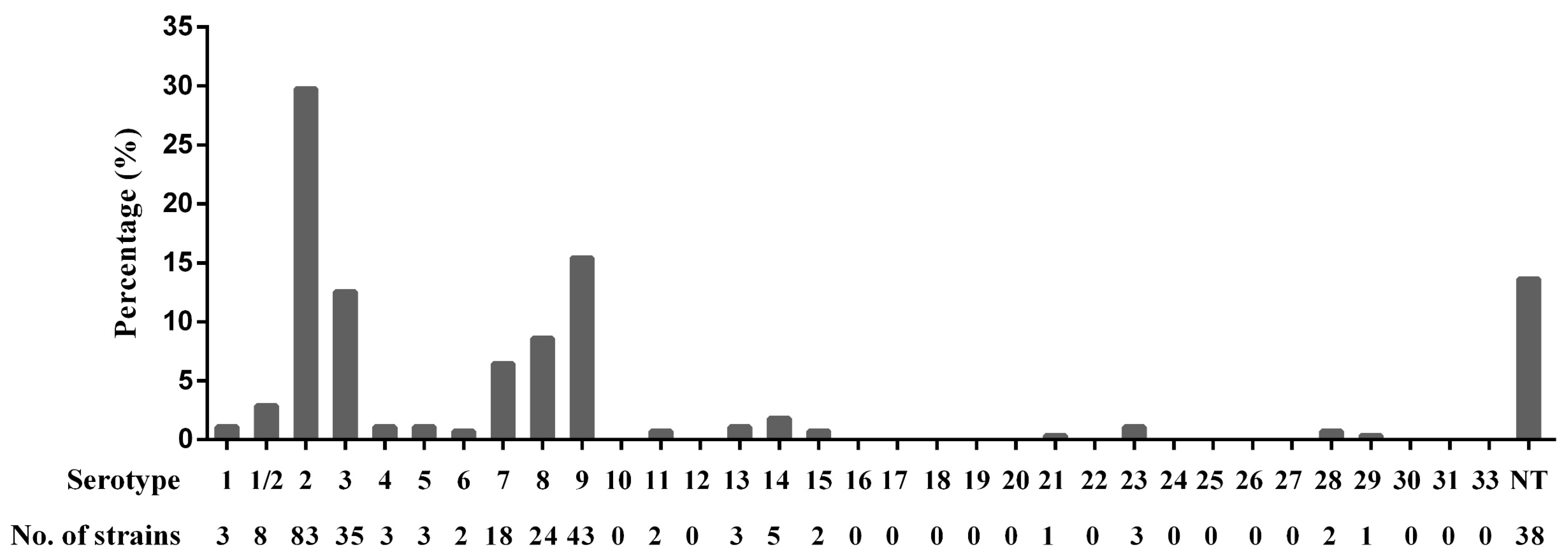
3.1. Serotypes of S. suis Isolates
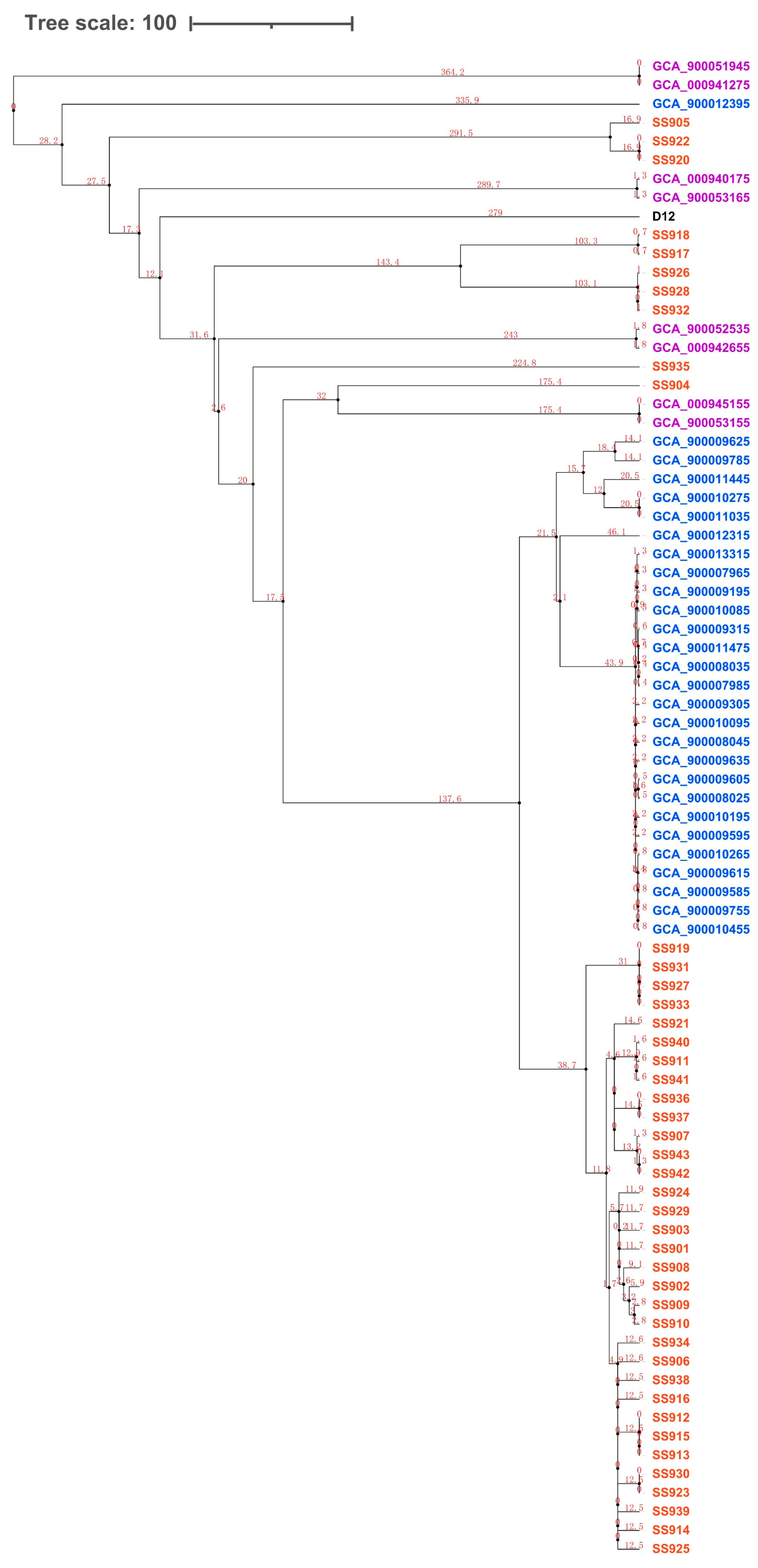
3.2. Evolution Analysis
3.3. Distribution of Virulence-Associated Genes among Serotype 9 Strains
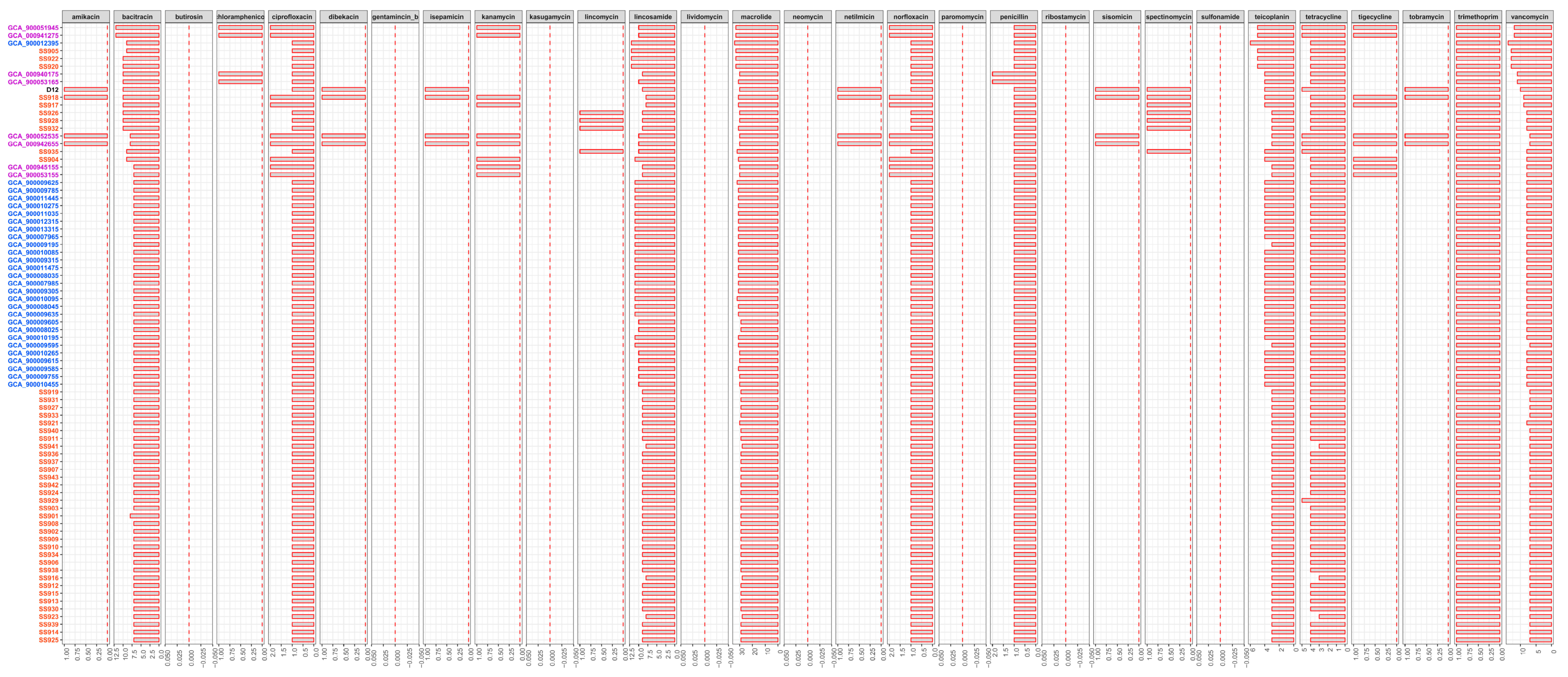
3.4. Difference in Drug Resistance Potential among Tested Strains
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ye, C.; Zheng, H.; Zhang, J.; Jing, H.; Wang, L.; Xiong, Y.; Wang, W.; Zhou, Z.; Sun, Q.; Luo, X.; et al. Clinical, Experimental, and Genomic Differences between Intermediately Pathogenic, Highly Pathogenic, and Epidemic Streptococcus suis. J. Infect. Dis. 2009, 199, 97–107. [Google Scholar] [CrossRef] [Green Version]
- Goyette-Desjardins, G.; Auger, J.P.; Xu, J.; Segura, M.; Gottschalk, M. Streptococcus suis, an important pig pathogen and emerging zoonotic agent-an update on the worldwide distribution based on serotyping and sequence typing. Emerg. Microbes Infect. 2014, 3, e45. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Huang, J.; Yu, J.; Xu, Z.; Liu, L.; Song, Y.; Sun, X.; Zhang, A.; Jin, M. Hp1330 contributes to Streptococcus suis virulence by inducing toll-like receptor 2- and erk1/2-dependent pro-inflammatory responses and influencing in vivo s. Suis loads. Front. Immunol. 2017, 8, 869. [Google Scholar] [CrossRef]
- Zhao, Y.; Liu, G.; Li, S.; Wang, M.; Song, J.; Wang, J.; Tang, J.; Li, M.; Hu, F. Role of a Type IV–Like Secretion System of Streptococcus suis 2 in the Development of Streptococcal Toxic Shock Syndrome. J. Infect. Dis. 2011, 204, 274–281. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Lin, L.; Fu, L.; Han, L.; Zhang, A. Neutrophil extracellular Taps play an important role in clearance of Streptococcus suis in vivo. Microbiol. Immunol. 2016, 60, 228–233. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.-I.; Jeon, A.B.; Jung, B.-Y.; Byun, J.-W.; Gottschalk, M.; Kim, A.; Kim, J.W.; Kim, H.-Y. Capsular serotypes, virulence-associated genes and antimicrobial susceptibility of Streptococcus suis isolates from pigs in Korea. J. Vet. Med. Sci. 2017, 79, 780–787. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Yang, Y.; Yan, S.; Liu, J.; Xu, Z.; Yu, J.; Song, Y.; Zhang, A.; Jin, M. A novel pro-inflammatory protein of Streptococcus suis 2 induces the toll-like receptor 2-dependent expression of pro-inflammatory cytokines in raw 264.7 macrophages via activation of erk1/2 pathway. Front. Microbiol. 2015, 6, 178. [Google Scholar] [CrossRef] [Green Version]
- Dong, W.; Zhu, Y.; Ma, Y.; Ma, J.; Zhang, Y.; Yuan, L.; Pan, Z.; Wu, Z.; Yao, H. Multilocus sequence typing and virulence genotyping of Streptococcus suis serotype 9 isolates revealed high genetic and virulence diversity. FEMS Microbiol. Lett. 2017, 364, 364. [Google Scholar] [CrossRef]
- Dekker, N.; Bouma, A.; Daemen, I.; Vernooij, H.; Van Leengoed, L.; Wagenaar, J.A.; Stegeman, A. Effect of Simultaneous Exposure of Pigs to Streptococcus suis Serotypes 2 and 9 on Their Colonization and Transmission, and on Mortality. Pathogens 2017, 6, 46. [Google Scholar] [CrossRef] [Green Version]
- Zheng, H.; Du, P.; Qiu, X.; Kerdsin, A.; Roy, D.; Bai, X.; Xu, J.; Vela, A.I.; Gottschalk, M. Genomic comparisons of Streptococcus suis serotype 9 strains recovered from diseased pigs in Spain and Canada. Vet. Res. 2018, 49, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okwumabua, O.; O’Connor, M.; Shull, E. A polymerase chain reaction (pcr) assay specific for Streptococcus suis based on the gene encoding the glutamate dehydrogenase. FEMS Microbiol. Lett. 2003, 218, 79–84. [Google Scholar] [CrossRef] [PubMed]
- Wei, Z.; Li, R.; Zhang, A.; He, H.; Hua, Y.; Xia, J.; Cai, X.; Chen, H.; Jin, M. Characterization of Streptococcus suis isolates from the diseased pigs in China between 2003 and 2007. Vet. Microbiol. 2009, 137, 196–201. [Google Scholar] [CrossRef]
- Pan, F.; Zhang, H.; Dong, X.; Ye, W.; He, P.; Zhang, S.; Zhu, J.X.; Zhong, N. Comparative genomic analysis of multidrug-resistant Streptococcus pneumoniae isolates. Infect. Drug Resist. 2018, 11, 659–670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fittipaldi, N.; Segura, M.; Grenier, D.; Gottschalk, M. Virulence factors involved in the pathogenesis of the infection caused by the swine pathogen and zoonotic agent Streptococcus suis. Future Microbiol. 2012, 7, 259–279. [Google Scholar] [CrossRef]
- Wang, H.; Wang, J.; Yu, P.; Ge, P.; Jiang, Y.; Xu, R.; Chen, R.; Liu, X. Identification of antibiotic resistance genes in the multidrug-resistant Acinetobacter baumannii strain, MDR-SHH02, using whole-genome sequencing. Int. J. Mol. Med. 2016, 39, 364–372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Segura, M.; Fittipaldi, N.; Calzas, C.; Gottschalk, M. Critical Streptococcus suis Virulence Factors: Are They All Really Critical? Trends Microbiol. 2017, 25, 585–599. [Google Scholar] [CrossRef]
- Wang, K.; Zhang, W.; Li, X.; Lu, C.; Chen, J.; Fan, W.; Huang, B. Characterization of Streptococcus suis Isolates from Slaughter Swine. Curr. Microbiol. 2013, 66, 344–349. [Google Scholar] [CrossRef]
- Qiao, M.; Ying, G.-G.; Singer, A.; Zhu, Y.-G. Review of antibiotic resistance in China and its environment. Environ. Int. 2018, 110, 160–172. [Google Scholar] [CrossRef] [Green Version]
- Pazda, M.; Kumirska, J.; Stepnowski, P.; Mulkiewicz, E. Antibiotic resistance genes identified in wastewater treatment plant systems—A review. Sci. Total Environ. 2019, 697, 134023. [Google Scholar] [CrossRef]
- Zhang, C.; Zhang, P.; Wang, Y.; Fu, L.; Liu, L.; Xu, D.; Hou, Y.; Li, Y.; Fu, M.; Wang, X.; et al. Capsular serotypes, antimicrobial susceptibility, and the presence of transferable oxazolidinone resistance genes in Streptococcus suis isolated from healthy pigs in China. Vet. Microbiol. 2020, 247, 108750. [Google Scholar] [CrossRef]
- Soares, T.C.S.; Paes, A.C.; Megid, J.; Ribolla, P.; Paduan, K.D.S.; Gottschalk, M. Antimicrobial susceptibility of Streptococcus suis isolated from clinically healthy swine in Brazil. Can. J. Vet. Res. Rev. Can. Rech. Vet. 2014, 78, 145–149. [Google Scholar]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, H.; Huang, J.; Hu, X.; Hu, M.; Zhang, Q.; Jin, M. Comparative Genome Analysis of Streptococcus suis Serotype 9 Isolates from China, The Netherland, and the U.K. Life 2021, 11, 1324. https://doi.org/10.3390/life11121324
Yang H, Huang J, Hu X, Hu M, Zhang Q, Jin M. Comparative Genome Analysis of Streptococcus suis Serotype 9 Isolates from China, The Netherland, and the U.K. Life. 2021; 11(12):1324. https://doi.org/10.3390/life11121324
Chicago/Turabian StyleYang, Huanhuan, Jingjing Huang, Xiaotong Hu, Min Hu, Qiang Zhang, and Meilin Jin. 2021. "Comparative Genome Analysis of Streptococcus suis Serotype 9 Isolates from China, The Netherland, and the U.K." Life 11, no. 12: 1324. https://doi.org/10.3390/life11121324
APA StyleYang, H., Huang, J., Hu, X., Hu, M., Zhang, Q., & Jin, M. (2021). Comparative Genome Analysis of Streptococcus suis Serotype 9 Isolates from China, The Netherland, and the U.K. Life, 11(12), 1324. https://doi.org/10.3390/life11121324