The Therapeutic Effectiveness of Full Spectrum Hemp Oil Using a Chronic Neuropathic Pain Model

 ,
,
Abstract
1. Introduction
2. Materials and Methods
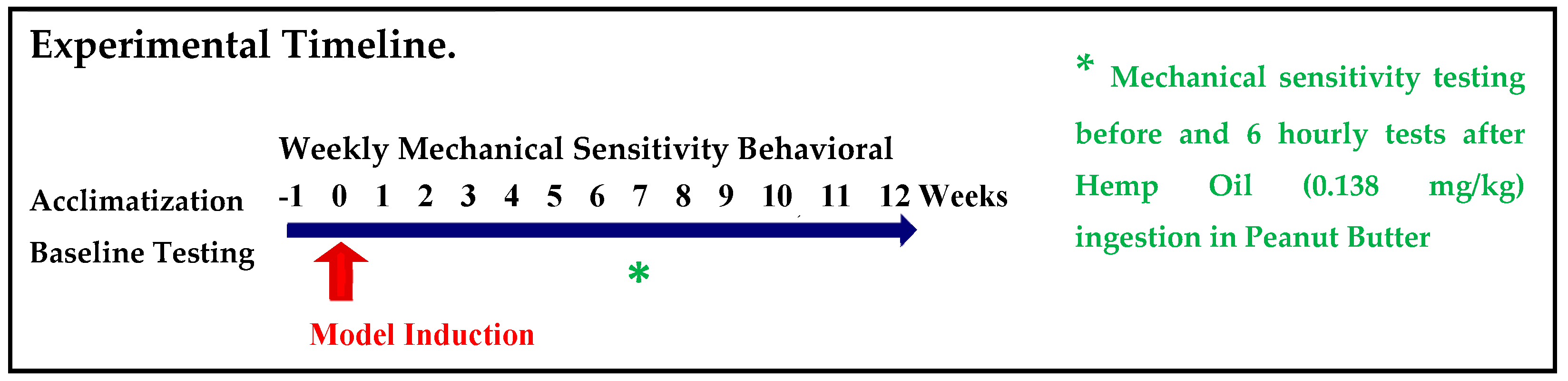
2.1. Experimental Plan
2.2. Animals
2.3. Surgical Procedures for Induction of the Model
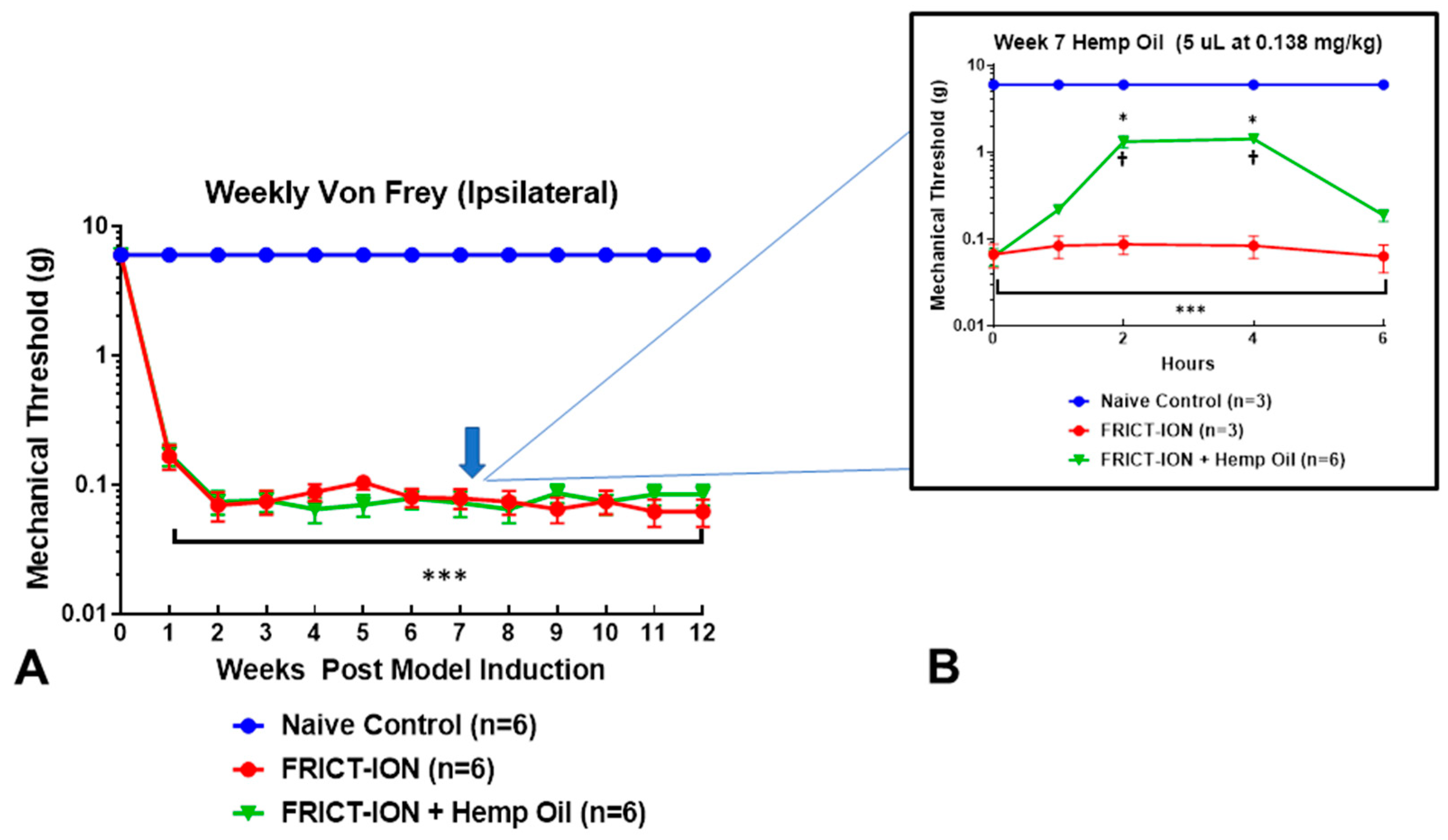
2.4. Assessment of Mechanical Allodynia
2.5. Hemp Oil Dosing
2.6. Hemp Cultivation and Oil Extraction
2.7. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Russo, E.B. Taming THC: Potential cannabis synergy and phytocannabinoid-terpenoid entourage effects. Br. J. Pharmacol. 2011, 163, 1344–1364. [Google Scholar] [CrossRef]
- Lewis, M.A.; Russo, E.B.; Smith, K.M. Pharmacological Foundations of Cannabis Chemovars. Planta Med. 2018, 84, 225–233. [Google Scholar] [CrossRef]
- Andre, C.M.; Hausman, J.-F.; Guerriero, G. Cannabis sativa: The Plant of the Thousand and One Molecules. Front. Plant Sci. 2016, 7, 19. [Google Scholar] [CrossRef]
- Fischedick, J.E.S. Cannabinoids and Terpenes as Chemotaxonomic Markers in Cannabis. Nat. Prod. Chem. Res. 2015, 3. [Google Scholar] [CrossRef]
- National Academies of Sciences, Engineering, and Medicine. The Health Effects of Cannabis and Cannabinoids: The Current State of Evidence and Recommendations for Research; National Academies Press: Washington, DC, USA, 2017. [Google Scholar] [CrossRef]
- Stith, S.S.; Vigil, J.M. Federal barriers to Cannabis research ALTHOUGH THE MAJORITY of the general Digital identifi ers for fungal species No surprise that comb jellies poop. Science 2016, 352, 1182. [Google Scholar]
- Kirk, J.M.; Doty, P.; De Wit, H. Effects of expectancies on subjective responses to oral Δ-tetrahydrocannabinol. Pharmacol. Biochem. Behav. 1998. [Google Scholar] [CrossRef]
- Metrik, J.; Rohsenow, D.J.; Monti, P.M.; McGeary, J.; Cook, T.A.; de Wit, H.; Kahler, C.W. Effectiveness of a Marijuana Expectancy Manipulation: Piloting the Balanced-Placebo Design for Marijuana. Exp. Clin. Psychopharmacol. 2009. [Google Scholar] [CrossRef]
- Bair, M.J.; Robinson, R.L.; Katon, W.; Kroenke, K. Depression and Pain Comorbidity: A Literature Review. Arch. Intern. Med. 2003. [Google Scholar] [CrossRef]
- Torta, R.; Ieraci, V.; Zizzi, F. A Review of the Emotional Aspects of Neuropathic Pain: From Comorbidity to Co-Pathogenesis. Pain Ther. 2017. [Google Scholar] [CrossRef]
- Montera, M.; Westlund, K. Minimally Invasive Oral Surgery Induction of the FRICT-ION Chronic Neuropathic Pain Model. Bio-Protocol 2020. [Google Scholar] [CrossRef]
- Hannaman, M.R.; Fitts, D.A.; Doss, R.M.; Weinstein, D.E.; Bryant, J.L. The refined biomimetic NeuroDigm GELTM model of neuropathic pain in a mature rat. F1000Research 2017. [Google Scholar] [CrossRef]
- Dutta, S.; Sengupta, P. Men and mice: Relating their ages. Life Sci. 2016. [Google Scholar] [CrossRef]
- Vicario, N.; Parenti, R.; Aricò, G.; Turnaturi, R.; Scoto, G.M.; Chiechio, S.; Parenti, C. Repeated activation of delta opioid receptors counteracts nerve injury-induced TNF-α up-regulation in the sciatic nerve of rats with neuropathic pain: A possible correlation with delta opioid receptors-mediated antiallodinic effect. Mol. Pain 2016. [Google Scholar] [CrossRef]
- Vicario, N.; Pasquinucci, L.; Spitale, F.M.; Chiechio, S.; Turnaturi, R.; Caraci, F.; Parenti, C. Simultaneous Activation of Mu and Delta Opioid Receptors Reduces Allodynia and Astrocytic Connexin 43 in an Animal Model of Neuropathic Pain. Mol. Neurobiol. 2019. [Google Scholar] [CrossRef]
- Lowery, J.J.; Raymond, T.J.; Giuvelis, D.; Bidlack, J.M.; Polt, R.; Bilsky, E.J. In vivo characterization of MMP-2200, a mixed δ/μ opioid agonist, in mice. J. Pharmacol. Exp. Ther. 2011. [Google Scholar] [CrossRef]
- Santello, M.; Nevian, T. Dysfunction of cortical dendritic integration in neuropathic pain reversed by serotoninergic neuromodulation. Neuron 2015. [Google Scholar] [CrossRef]
- Santello, M.; Bisco, A.; Nevian, N.E.; Lacivita, E.; Leopoldo, M.; Nevian, T. The brain-penetrant 5-HT7 receptor agonist LP-211 reduces the sensory and affective components of neuropathic pain. Neurobiol. Dis. 2017. [Google Scholar] [CrossRef]
- Sogabe, S.; Yagasaki, Y.; Onozawa, K.; Kawakami, Y. Mesocortical dopamine system modulates mechanical nociceptive responses recorded in the rat prefrontal cortex. BMC Neurosci. 2013. [Google Scholar] [CrossRef]
- Santana, N.; Mengod, G.; Artigas, F. Quantitative analysis of the expression of dopamine D1 and D2 receptors in pyramidal and GABAergic neurons of the rat prefrontal cortex. Cereb Cortex 2009. [Google Scholar] [CrossRef]
- Kindred, J.H.; Li, K.; Ketelhut, N.B.; Proessl, F.; Fling, B.W.; Honce, J.M.; Shaffer, W.R.; Rudroff, T. Cannabis use in people with Parkinson’s disease and Multiple Sclerosis: A web-based investigation. Complement. Ther. Med. 2017, 33, 99–104. [Google Scholar] [CrossRef]
- Sun, T.; Wang, J.; Lia, X.; Lia, Y.; Feng, D.; Shi, W.; Zhao, M.; Wang, J.; Wua, Y. Gastrodin relieved complete Freund’s adjuvant-induced spontaneous pain by inhibiting inflammatory response. Int. Immunopharmacol. 2016. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.F.; Guan, S.Y.; Luo, L.; Li, Y.J.; Yang, L.; Zhou, X.X.; Liu, G. Tetrahydroxystilbene glucoside relieves the chronic inflammatory pain by inhibiting neuronal apoptosis, microglia activation, and GluN2B overexpression in anterior cingulate cortex. Mol. Pain. 2018. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, M. Ethical guidelines for investigations of experimental pain in conscious animals. Pain 1983. [Google Scholar] [CrossRef]
- Lyons, D.N.; Zhang, L.; Pandya, J.D.; Danaher, R.J.; Ma, F.; Miller, C.S.; Westlund, K.N. Combination Drug Therapy of Pioglitazone and D-cycloserine Attenuates Chronic Orofacial Neuropathic Pain and Anxiety by Improving Mitochondrial Function Following Trigeminal Nerve Injury. Clin. J. Pain 2018. [Google Scholar] [CrossRef]
- Chaplan, S.R.; Bach, F.W.; Pogrel, J.W.; Chung, J.M.; Yaksh, T.L. Quantitative assessment of tactile allodynia in the rat paw. J. Neurosci. Methods 1994. [Google Scholar] [CrossRef]
- Nielsen, S.; Sabioni, P.; Trigo, J.M.; Ware, M.A.; Betz-Stablein, B.D.; Murnion, B.; Le Foll, B. Opioid-sparing effect of cannabinoids: A systematic review and meta-analysis. Neuropsychopharmacology 2017. [Google Scholar] [CrossRef]
- Hill, K.P. Medical marijuana for treatment of chronic pain and other medical and psychiatric problems: A clinical review. JAMA J. Am. Med. Assoc. 2015. [Google Scholar] [CrossRef]
- Rabgay, K.; Waranuch, N.; Chaiyakunapruk, N.; Sawangjit, R.; Ingkaninan, K.; Dilokthornsakul, P. The effects of cannabis, cannabinoids, and their administration routes on pain control efficacy and safety: A systematic review and network meta-analysis. J. Am. Pharm. Assoc. 2020. [Google Scholar] [CrossRef]
- Woodhams, S.G.; Chapman, V.; Finn, D.P.; Hohmann, A.G.; Neugebauer, V. The cannabinoid system and pain. Neuropharmacology 2017. [Google Scholar] [CrossRef]
- Kamal, B.S.; Kamal, F.; Lantela, D.E. Cannabis and the Anxiety of Fragmentation—A Systems Approach for Finding an Anxiolytic Cannabis Chemotype. Front. Neurosci. 2018. [Google Scholar] [CrossRef]
- Poli, P.; Crestani, F.; Salvadori, C.; Valenti, I.; Sannino, C. Medical cannabis in patients with chronic pain: Effect on pain relief, pain disability, and psychological aspects. A prospective non randomized single arm clinical trial. Clin. Ter. 2018. [Google Scholar] [CrossRef]
- Stith, S.S.; Vigil, J.M.; Adams, I.M.; Reeve, A.P. Effects of Legal Access to Cannabis on Scheduled II–V Drug Prescriptions. J. Am. Med. Dir. Assoc. 2018. [Google Scholar] [CrossRef] [PubMed]
- Stith, S.S.; Vigil, J.M.; Brockelman, F.; Keeling, K.; Hall, B. The Association between Cannabis Product Characteristics and Symptom Relief. Sci. Rep. 2019, 9, 1–8. [Google Scholar] [CrossRef]
- Piper, B.J.; DeKeuster, R.M.; Beals, M.L.; Cobb, C.M.; Burchman, C.A.; Perkinson, L.; Abess, A.T. Substitution of medical cannabis for pharmaceutical agents for pain, anxiety, and sleep. J. Psychopharmacol. 2017, 31, 569–575. [Google Scholar] [CrossRef]
- Dai, H.; Richter, K.P. A National Survey of Marijuana Use Among US Adults with Medical Conditions, 2016–2017. JAMA J. Am. Med. Assoc. 2019, 2, e1911936. [Google Scholar] [CrossRef]
- Degenhardt, L.; Dierker, L.; Chiu, W.T.; Medina-Mora, M.E.; Neumark, Y.; Sampson, N.; De Girolamo, G. Evaluating the drug use “gateway” theory using cross-national data: Consistency and associations of the order of initiation of drug use among participants in the WHO World Mental Health Surveys. Drug Alcohol Depend. 2010, 108, 84–97. [Google Scholar] [CrossRef]
- Vigil, J.M.; Stith, S.S.; Adams, I.M.; Reeve, A.P. Associations between medical cannabis and prescription opioid use in chronic pain patients: A preliminary cohort study. PLoS ONE 2017, 12, e0187795. [Google Scholar] [CrossRef]
- Li, X.; Vigil, J.M.; Stith, S.S.; Brockelman, F.; Keeling, K.; Hall, B. The effectiveness of self-directed medical cannabis treatment for pain. Complement. Ther. Med. 2019. [Google Scholar] [CrossRef]
- Stith, S.S.; Vigil, J.M.; Brockelman, F.; Keeling, K.; Hall, B. Patient-reported symptom relief following medical cannabis consumption. Front. Pharmacol. 2018, 9, 916. [Google Scholar] [CrossRef]
- Ma, F.; Zhang, L.; Lyons, D.; Westlund, K.N. Orofacial neuropathic pain mouse model induced by Trigeminal Inflammatory Compression (TIC) of the infraorbital nerve. Mol. Brain 2012. [Google Scholar] [CrossRef]
- Vos, B.P.; Strassman, A.M.; Maciewicz, R.J. Behavioral evidence of trigeminal neuropathic pain following chronic constriction injury to the rat’s infraorbital nerve. J. Neurosci. 1994. [Google Scholar] [CrossRef]
- Genaro, K.; Fabris, D.; Arantes, A.L.F.; Zuardi, A.W.; Crippa, J.A.S.; Prado, W.A. Cannabidiol is a potential therapeutic for the affective-motivational dimension of incision pain in rats. Front. Pharmacol. 2017. [Google Scholar] [CrossRef]
- Gewandter, J.S.; Dworkin, R.H.; Turk, D.C.; Farrar, J.T.; Fillingim, R.B.; Gilron, I.; Robinson, J.P. Research design considerations for chronic pain prevention clinical trials: IMMPACT recommendations. Pain 2015. [Google Scholar] [CrossRef]
- Cooper, S.A.; Desjardins, P.J.; Turk, D.C.; Dworkin, R.H.; Katz, N.P.; Kehlet, H.; Croll, S. Research design considerations for single-dose analgesic clinical trials in acute pain: IMMPACT recommendations. Pain 2016. [Google Scholar] [CrossRef]
- Haviv, Y.; Zadik, Y.; Sharav, Y.; Benoliel, R. Painful Traumatic Trigeminal Neuropathy: An Open Study on the Pharmacotherapeutic Response to Stepped Treatment. J. Oral Facial Pain Headache 2014. [Google Scholar] [CrossRef]
- Baad-Hansen, L.; Benoliel, R. Neuropathic orofacial pain: Facts and fiction. Cephalalgia 2017. [Google Scholar] [CrossRef]
- Tham, M.; Yilmaz, O.; Alaverdashvili, M.; Kelly, M.E.M.; Denovan-Wright, E.M.; Laprairie, R.B. Allosteric and orthosteric pharmacology of cannabidiol and cannabidiol-dimethylheptyl at the type 1 and type 2 cannabinoid receptors. Br. J. Pharmacol. 2019. [Google Scholar] [CrossRef]
- Thomas, A.; Baillie, G.L.; Phillips, A.M.; Razdan, R.K.; Ross, R.A.; Pertwee, R.G. Cannabidiol displays unexpectedly high potency as an antagonist of CB 1 and CB 2 receptor agonists in vitro. Br. J. Pharmacol. 2007, 150, 613–623. [Google Scholar] [CrossRef]
- Laprairie, R.B.; Bagher, A.M.; Kelly, M.E.M.; Denovan-Wright, E.M. Cannabidiol is a negative allosteric modulator of the cannabinoid CB1 receptor. Br. J. Pharmacol. 2015. [Google Scholar] [CrossRef]
- McPartland, J.M.; Duncan, M.; Di Marzo, V.; Pertwee, R.G. Are cannabidiol and Δ9-tetrahydrocannabivarin negative modulators of the endocannabinoid system? A systematic review. Br. J. Pharmacol. 2015, 172, 737–753. [Google Scholar] [CrossRef]
- Pertwee, R.G. The diverse CB1 and CB2 receptor pharmacology of three plant cannabinoids: delta9-tetrahydrocannabinol, cannabidiol and delta9-tetrahydrocannabivarin. Br. J. Pharmacol. 2008, 153, 199–215. [Google Scholar] [CrossRef] [PubMed]
- De Gregorio, D.; McLaughlin, R.J.; Posa, L.; Ochoa-Sanchez, R.; Enns, J.; Lopez-Canul, M.; Gobbi, G. Cannabidiol modulates serotonergic transmission and reverses both allodynia and anxiety-like behavior in a model of neuropathic pain. Pain 2019, 160, 136. [Google Scholar] [CrossRef] [PubMed]
- Russo, E.B.; Burnett, A.; Hall, B.; Parker, K.K. Agonistic properties of cannabidiol at 5-HT1a receptors. Neurochem. Res. 2005. [Google Scholar] [CrossRef] [PubMed]
- Iannotti, F.A.; Hill, C.L.; Leo, A.; Alhusaini, A.; Soubrane, C.; Mazzarella, E.; Stephens, G.J. Nonpsychotropic plant cannabinoids, Cannabidivarin (CBDV) and Cannabidiol (CBD), activate and desensitize Transient Receptor Potential Vanilloid 1 (TRPV1) channels in vitro: Potential for the treatment of neuronal hyperexcitability. ACS Chem. Neurosci. 2014, 5, 1131–1141. [Google Scholar] [CrossRef]
- Bisogno, T.; Hanuš, L.; De Petrocellis, L.; Tchilibon, S.; Ponde, D.E.; Brandi, I.; Di Marzo, V. Molecular targets for cannabidiol and its synthetic analogues: Effect on vanilloid VR1 receptors and on the cellular uptake and enzymatic hydrolysis of anandamide. Br. J. Pharmacol. 2001. [Google Scholar] [CrossRef]
- Mendis GD, C.; Berecki, G.; Morrisroe, E.; Pachernegg, S.; Li, M.; Varney, M.; Petrou, S. Discovering the pharmacodynamics of conolidine and cannabidiol using a cultured neuronal network based workflow. Sci. Rep. 2019. [Google Scholar] [CrossRef]
- Bakas, T.; van Nieuwenhuijzen, P.S.; Devenish, S.O.; McGregor, I.S.; Arnold, J.C.; Chebib, M. The direct actions of cannabidiol and 2-arachidonoyl glycerol at GABAA receptors. Pharmacol. Res. 2017. [Google Scholar] [CrossRef]
- Ahrens, J.; Demir, R.; Leuwer, M.; De La Roche, J.; Krampfl, K.; Foadi, N.; Haeseler, G. The nonpsychotropic cannabinoid cannabidiol modulates and directly activates alpha-1 and alpha-1-beta glycine receptor function. Pharmacology 2009. [Google Scholar] [CrossRef]
- Rodríguez-Muñoz, M.; Onetti, Y.; Cortés-Montero, E.; Garzón, J.; Sánchez-Blázquez, P. Cannabidiol enhances morphine antinociception, diminishes NMDA-mediated seizures and reduces stroke damage via the sigma 1 receptor. Mol. Brain 2018. [Google Scholar] [CrossRef]
- Ward, S.J.; McAllister, S.D.; Kawamura, R.; Murase, R.; Neelakantan, H.; Walker, E.A. Cannabidiol inhibits paclitaxel-induced neuropathic pain through 5-HT 1A receptors without diminishing nervous system function or chemotherapy efficacy. Br. J. Pharmacol. 2014. [Google Scholar] [CrossRef]
- Costa, B.; Giagnoni, G.; Franke, C.; Trovato, A.E.; Colleoni, M. Vanilloid TRPV1 receptor mediates the antihyperalgesic effect of the nonpsychoactive cannabinoid, cannabidiol, in a rat model of acute inflammation. Br. J. Pharmacol. 2004. [Google Scholar] [CrossRef] [PubMed]
- Dunlap, K.; Luebke, J.I.; Turner, T.J. Exocytotic Ca2+ channels in mammalian central neurons. Trends Neurosci. 1995. [Google Scholar] [CrossRef]
- Berecki, G.; Motin, L.; Haythornthwaite, A.; Vink, S.; Bansal, P.; Drinkwater, R.; Christie, M.J. Analgesic ω-conotoxins CVIE and CVIF selectively and voltage-dependently block recombinant and native N-type calcium channels. Mol. Pharmacol. 2010. [Google Scholar] [CrossRef] [PubMed]
- Xiong, W.; Cui, T.; Cheng, K.; Yang, F.; Chen, S.R.; Willenbring, D.; Zhang, L. Cannabinoids suppress inflammatory and neuropathic pain by targeting α3 glycine receptors. J. Exp. Med. 2012. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.F.; Linher-Melville, K.; Niazmand, M.J.; Sharma, M.; Shahid, A.; Zhu, K.L.; Singh, G. An evaluation of the anti-hyperalgesic effects of cannabidiolic acid-methyl ester in a preclinical model of peripheral neuropathic pain. Br. J. Pharmacol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Nadal, X.; del Río, C.; Casano, S.; Palomares, B.; Ferreiro-Vera, C.; Navarrete, C.; Morello, G. Tetrahydrocannabinolic acid is a potent PPARγ agonist with neuroprotective activity. Br. J. Pharmacol. 2017. [Google Scholar] [CrossRef]
- Harris, H.M.; Rousseau, M.A.; Wanas, A.S.; Radwan, M.M.; Caldwell, S.; Sufka, K.J.; ElSohly, M.A. Role of Cannabinoids and Terpenes in Cannabis-Mediated Analgesia in Rats. Cannabis Cannabinoid Res. 2019. [Google Scholar] [CrossRef]
- Gallily, R.; Yekhtin, Z.; Hanuš, L.O. The Anti-Inflammatory Properties of Terpenoids from Cannabis. Cannabis Cannabinoid Res. 2018. [Google Scholar] [CrossRef]
- Lorenzetti, B.B.; Souza, G.E.P.; Sarti, S.J.; Santos Filho, D.; Ferreira, S.H. Myrcene mimics the peripheral analgesic activity of lemongrass tea. J. Ethnopharmacol. 1991. [Google Scholar] [CrossRef]
- Mohamed Hanaa, A.R.; Sallam, Y.I.; El-Leithy, A.S.; Aly, S.E. Lemongrass (Cymbopogon citratus) essential oil as affected by drying methods. Ann. Agric. Sci. 2012. [Google Scholar] [CrossRef]
- Gramosa, M.; Silveira, E. Volatile constituents of Copalifera langsdorfffi from the Brazilian northeast. J. Essent Oil Res. 2005, 17, 130–132. [Google Scholar] [CrossRef]
- Da Trindade, R.; da Silva, J.K.; Setzer, W.N. Copaifera of the neotropics: A review of the phytochemistry and pharmacology. Int. J. Mol. Sci. 2018, 19, 1511. [Google Scholar] [CrossRef]
- Klauke, A.L.; Racz, I.; Pradier, B.; Markert, A.; Zimmer, A.M.; Gertsch, J.; Zimmer, A. The cannabinoid CB2 receptor-selective phytocannabinoid beta-caryophyllene exerts analgesic effects in mouse models of inflammatory and neuropathic pain. Eur. Neuropsychopharmacol. 2014. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.S.; Lee, H.J.; Jeon, Y.D.; Han, Y.H.; Kee, J.Y.; Kim, H.J.; Kim, S.J. Alpha-Pinene Exhibits Anti-Inflammatory Activity Through the Suppression of MAPKs and the NF-κB Pathway in Mouse Peritoneal Macrophages. Am. J. Chin. Med. 2015, 43, 731–742. [Google Scholar] [CrossRef] [PubMed]
- Quintão, N.L.M.; Da Silva, G.F.; Antonialli, C.S.; Rocha, L.W.; Filho, V.C.; Cicció, J.F. Chemical composition and evaluation of the anti-hypernociceptive effect of the essential oil extracted from the leaves of Ugni myricoides on inflammatory and neuropathic models of pain in mice. Planta Med. 2010. [Google Scholar] [CrossRef]
- Gatchel, R.J.; Peng, Y.B.; Peters, M.L.; Fuchs, P.N.; Turk, D.C. The Biopsychosocial Approach to Chronic Pain: Scientific Advances and Future Directions. Psychol. Bull. 2007. [Google Scholar] [CrossRef]
- McCaffrey, M.; Pasero, C. Pain: Clinical Manual, 2nd ed.; Mosby Inc.: St. Louis, MO, USA, 1999. [Google Scholar]
- Vigil, J.M. No pain, no social gains: A social-signaling perspective of human pain behaviors. World J. Anesthesiol. 2014, 3, 18. [Google Scholar] [CrossRef]
- Vigil, J.M. A socio-relational framework of sex differences in the expression of emotion. Behav. Brain Sci. 2009. [Google Scholar] [CrossRef]
- Vigil, J.M.; Coulombe, P. Biological sex and social setting affects pain intensity and observational coding of other people’s pain behaviors. Pain 2011. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Phytochemical | % Total Weight | mg/g |
|---|---|---|
| Cannabichromene (CBC) | 0.120 | 1.200 |
| Cannabigerolic Acid (CBGA) | 0.281 | 2.810 |
| Cannabigerol (CBG) | 0.046 | 0.460 |
| Tetrahydrocannabivarin (THCV) | 0.022 | 0.220 |
| Delta-8-Tetrahydrocannabinol (Δ8THC) | 0.000 | 0.000 |
| Cannabidivarin (CBDV) | 0.000 | 0.000 |
| Cannabinol (CBN) | 0.000 | 0.000 |
| Cannabidiolic Acid (CBDA) | 14.464 | 144.640 |
| Cannabidiol (CBD) | 0.506 | 5.060 |
| Delta-9-Tetrahydrocannabinol (Δ9THC) | 0.057 | 0.570 |
| Tetrahydrocannabinolic acid (THCA) | 0.661 | 6.610 |
| Total | 16.157 | 161.570 |
| Phytochemical | % Total Weight | mg/g |
|---|---|---|
| Alpha-Humulene | 0.036 | 0.360 |
| Alpha-Pinene | 0.048 | 0.480 |
| Beta-Myrcene | 0.256 | 2.560 |
| Alpha-Bisabolol | 0.040 | 0.400 |
| Beta-Caryophyllene | 0.121 | 1.210 |
| Limonene | 0.022 | 0.220 |
| Guaiol | 0.043 | 0.430 |
| Farnesene | 0.080 | 0.800 |
| Total | 0.646 | 6.460 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vigil, J.M.; Montera, M.A.; Pentkowski, N.S.; Diviant, J.P.; Orozco, J.; Ortiz, A.L.; Rael, L.J.; Westlund, K.N. The Therapeutic Effectiveness of Full Spectrum Hemp Oil Using a Chronic Neuropathic Pain Model. Life 2020, 10, 69. https://doi.org/10.3390/life10050069
Vigil JM, Montera MA, Pentkowski NS, Diviant JP, Orozco J, Ortiz AL, Rael LJ, Westlund KN. The Therapeutic Effectiveness of Full Spectrum Hemp Oil Using a Chronic Neuropathic Pain Model. Life. 2020; 10(5):69. https://doi.org/10.3390/life10050069
Chicago/Turabian StyleVigil, Jacob M., Marena A. Montera, Nathan S. Pentkowski, Jegason P. Diviant, Joaquin Orozco, Anthony L. Ortiz, Lawrence J. Rael, and Karin N. Westlund. 2020. "The Therapeutic Effectiveness of Full Spectrum Hemp Oil Using a Chronic Neuropathic Pain Model" Life 10, no. 5: 69. https://doi.org/10.3390/life10050069
APA StyleVigil, J. M., Montera, M. A., Pentkowski, N. S., Diviant, J. P., Orozco, J., Ortiz, A. L., Rael, L. J., & Westlund, K. N. (2020). The Therapeutic Effectiveness of Full Spectrum Hemp Oil Using a Chronic Neuropathic Pain Model. Life, 10(5), 69. https://doi.org/10.3390/life10050069