Biosignatures Associated with Freshwater Microbialites

 ,
,  , ,
, ,
Abstract
1. Introduction
2. Methods
2.1. Site Description
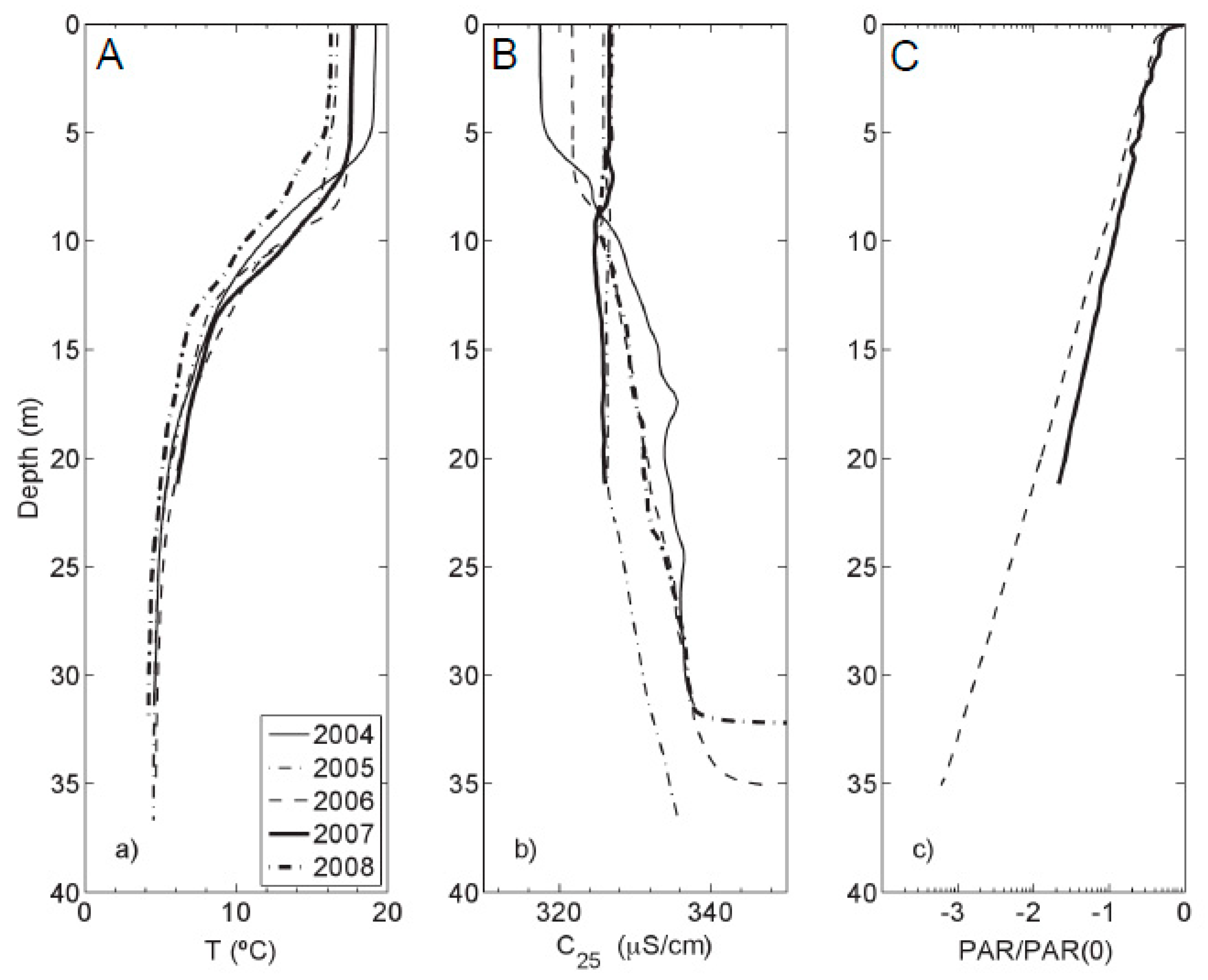
2.2. Physical Properties
2.3. Sample Collection and Analysis
2.3.1. Water Samples
2.3.2. Phospholipid Fatty Acid Extraction
2.3.3. Stable Isotope Analysis
2.3.4. Scanning Electron Microscopy and Confocal Laser Scanning Microscopy
2.3.5. DNA Extraction, Purity and Concentration Measurements
2.3.6. Quantitative PCR Measurements and Standard Curve Construction
2.3.7. Statistical Analyses
3. Results
3.1. Physical Properties of Kelly Lake
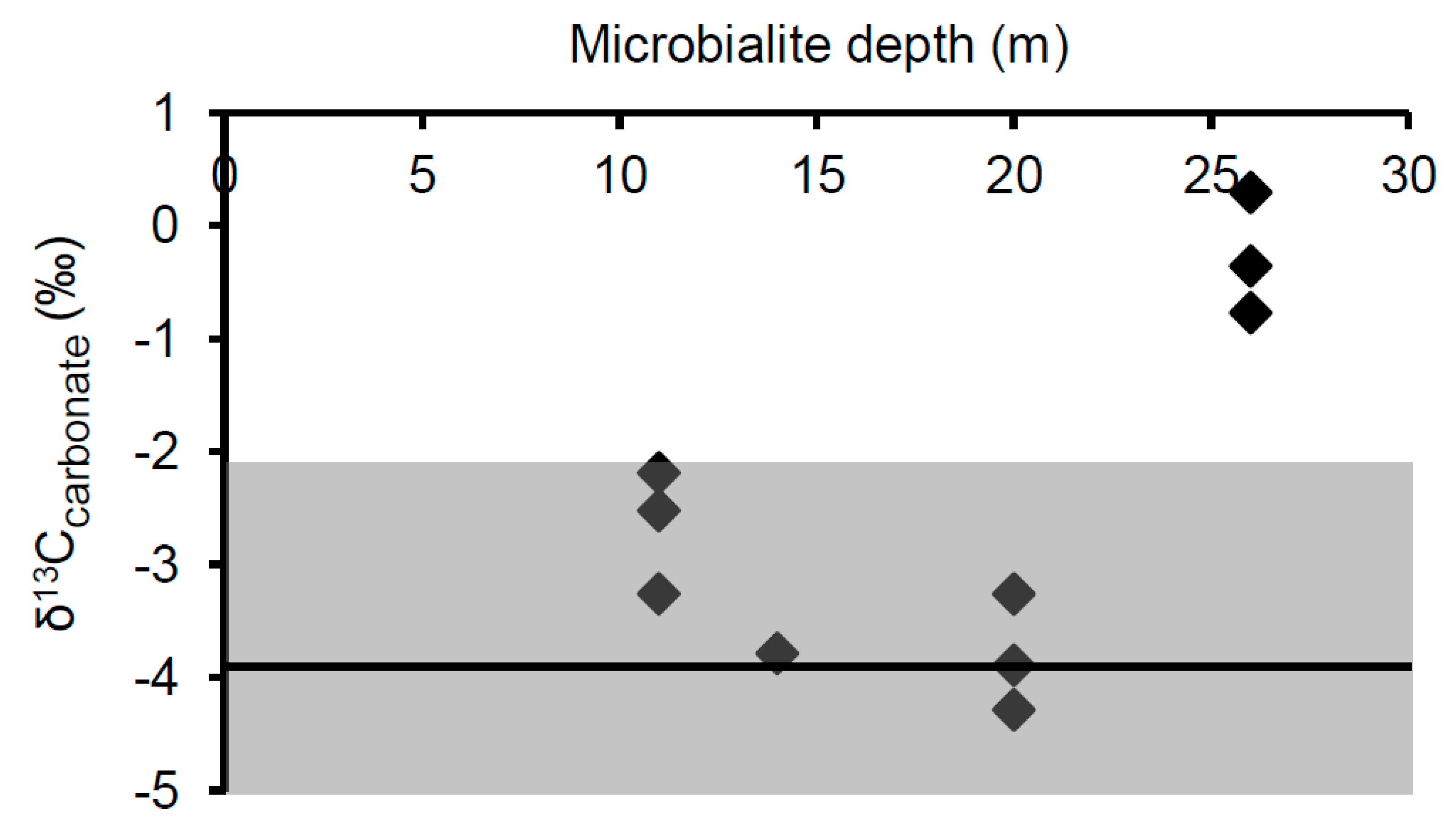
3.2. Isotopic Composition of DIC and Microbialite Carbonate
3.3. Isotopic Composition of Microbialite Bulk Organic Material
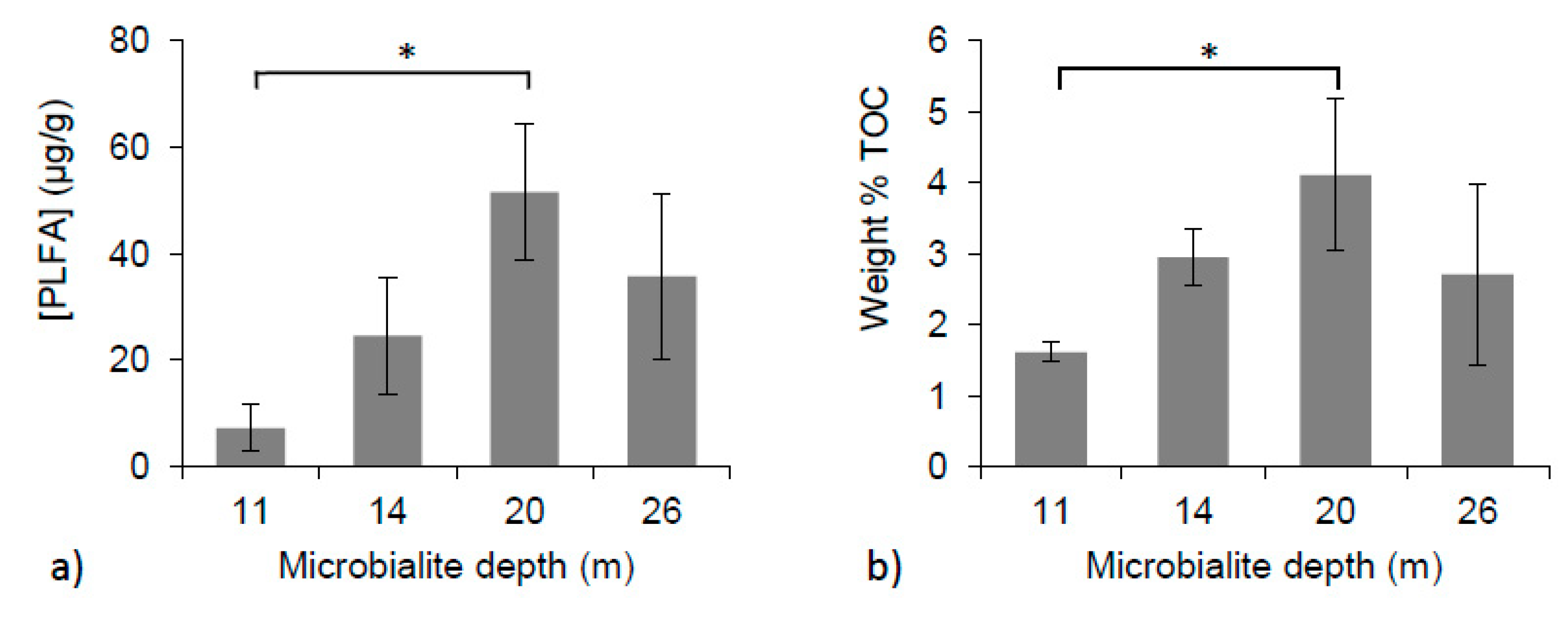
3.4. PLFA Concentrations and % Total Organic Carbon
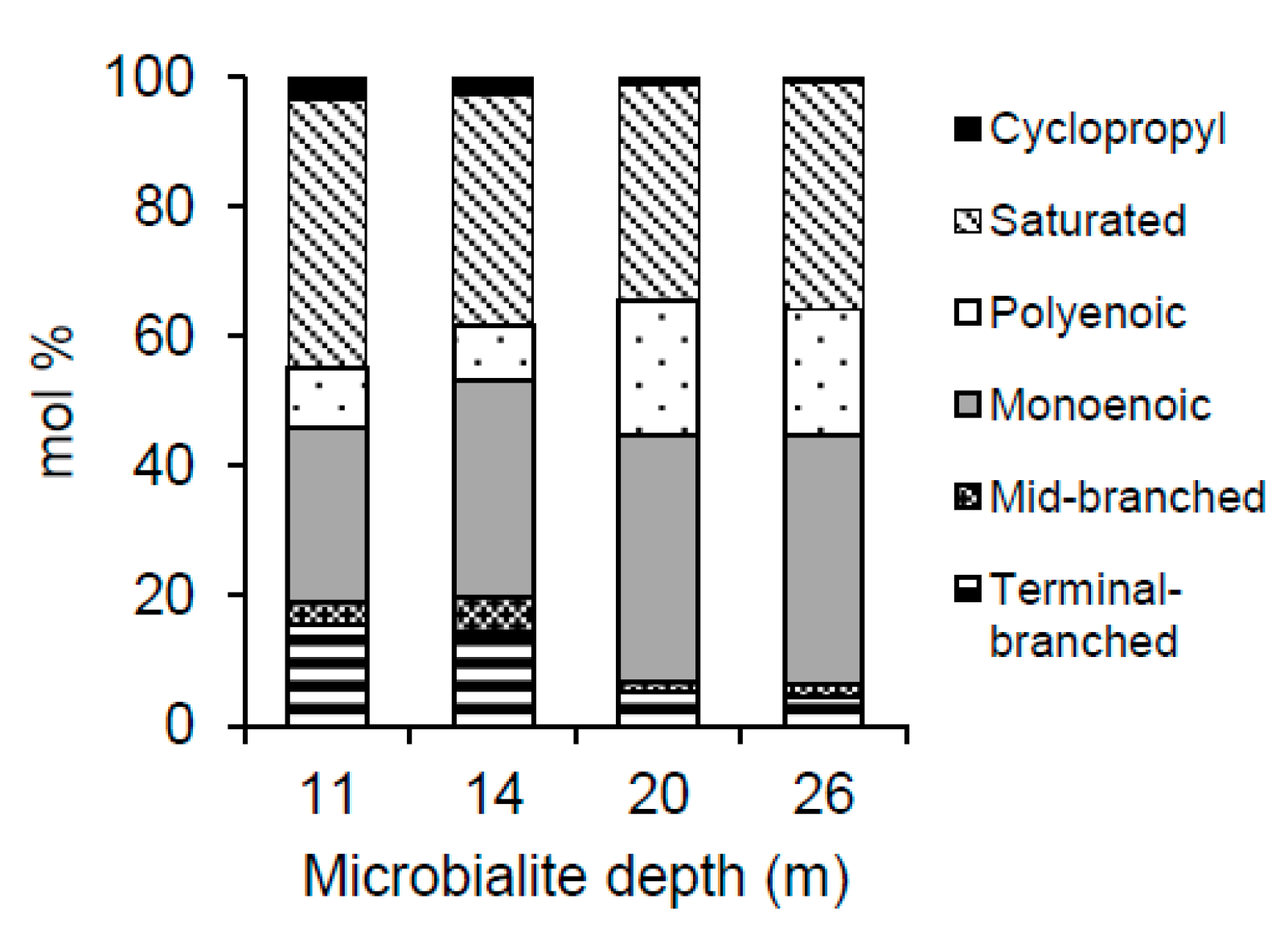
3.5. PLFA Profiles
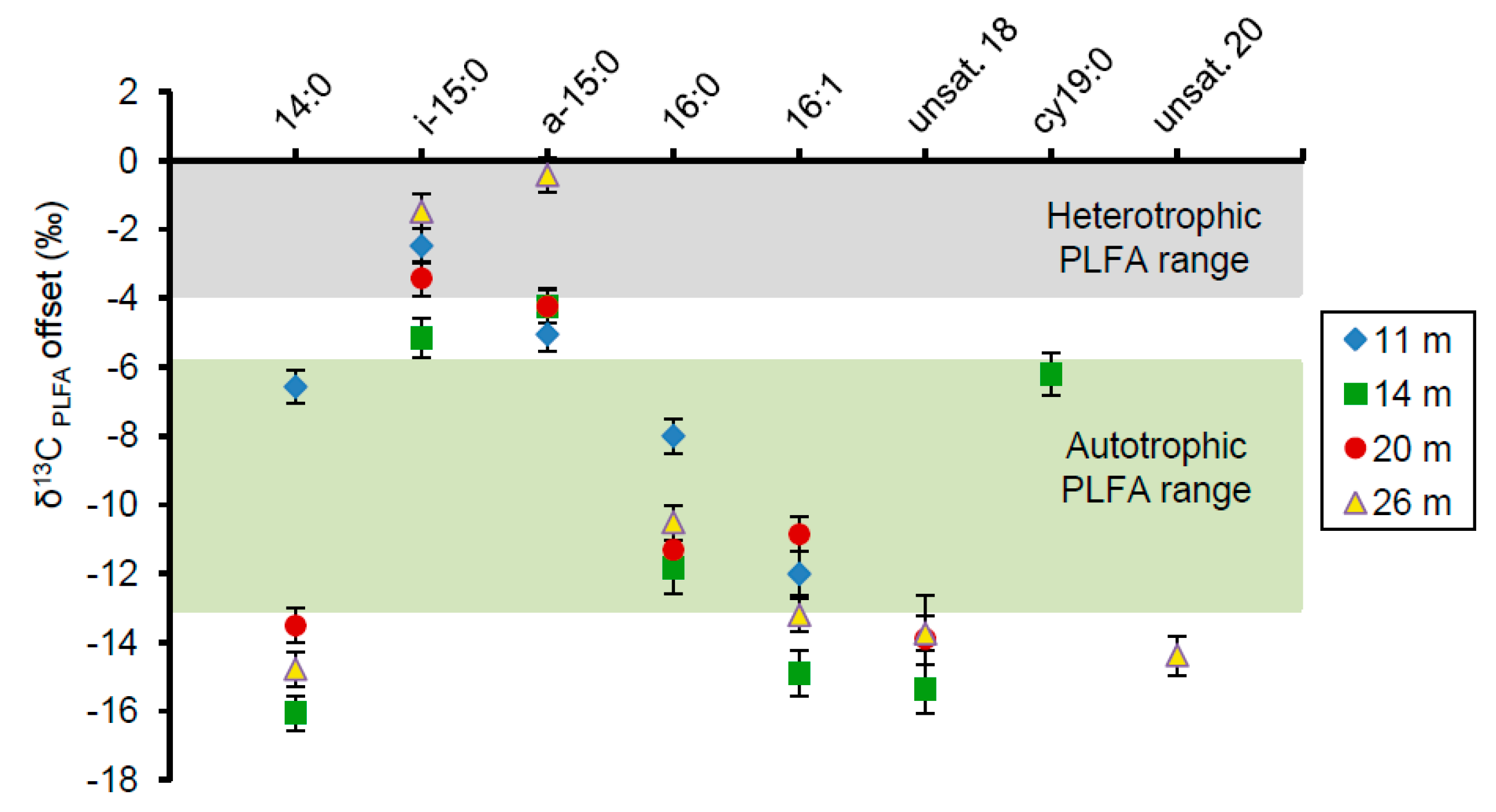
3.6. Isotopic Composition of Microbialite PLFAs
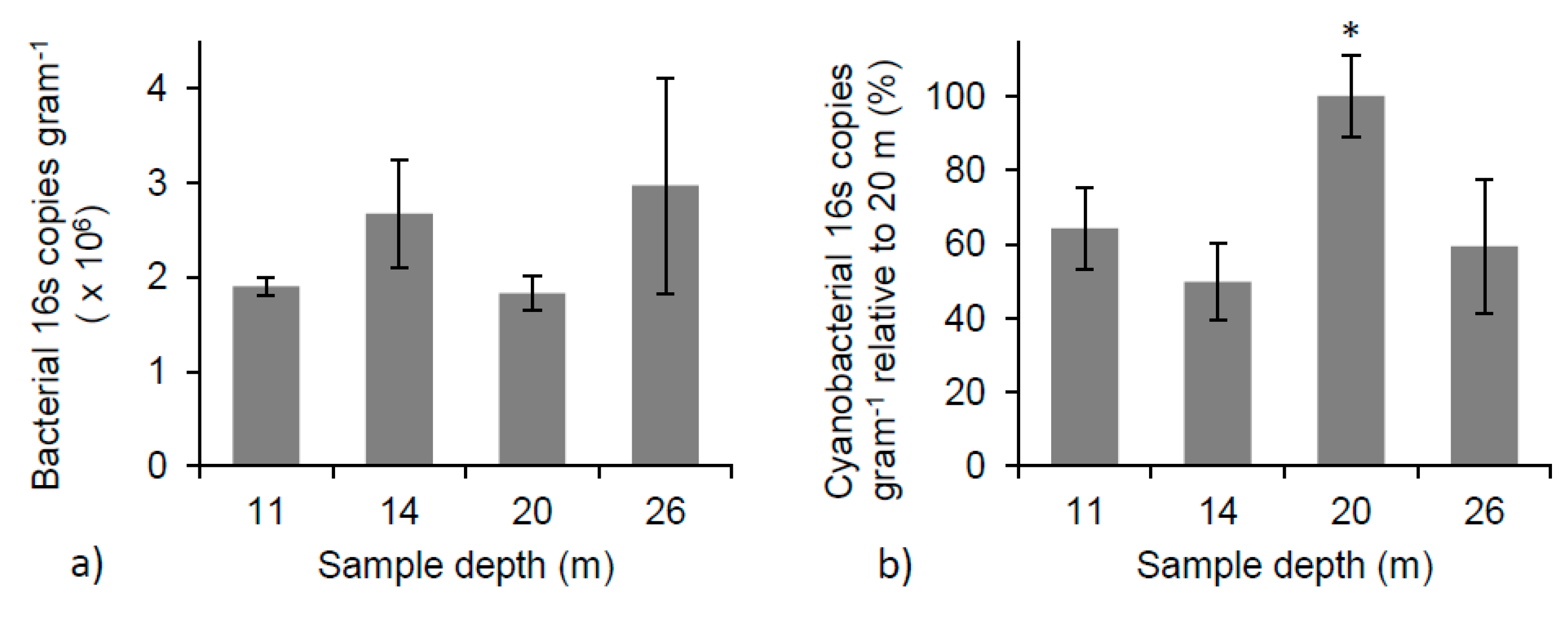
3.7. Microbial Community Relative Abundance
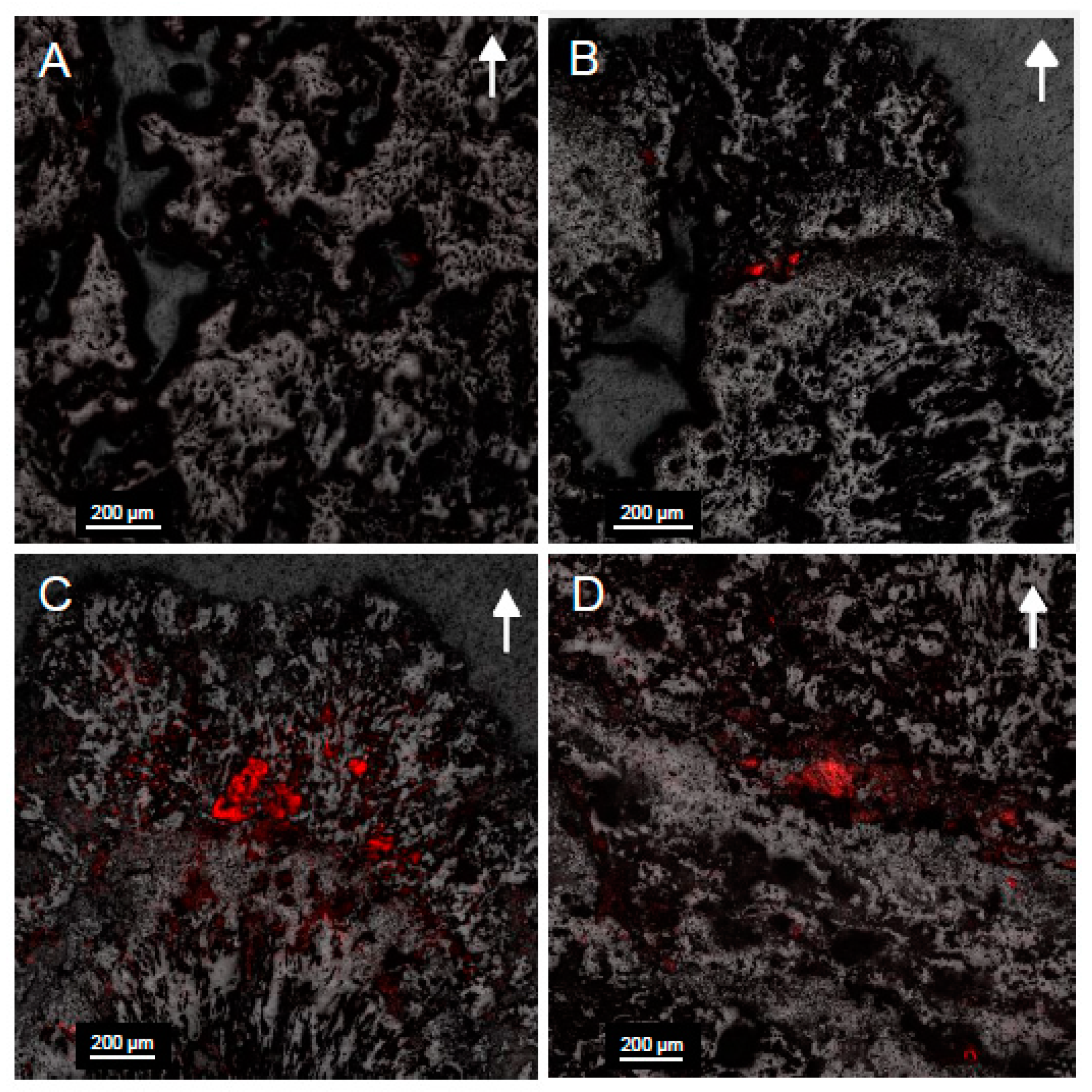
3.8. Microscopy
4. Discussion
4.1. Isotopic Biosignatures in Microbialite Carbonate
4.2. Predominance of Photoautotrophic Microorganisms
4.3. PLFA and Biomass Distribution
4.4. Comparison to Pavilion Lake
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Andres, M.S.; Sumner, D.Y.; Reid, R.P.; Swart, P.K. Isotopic fingerprints of microbial respiration in aragonite from Bahamian stromatolites. Geology 2006, 34, 973–976. [Google Scholar] [CrossRef]
- Breitbart, M.; Hoare, A.; Nitti, A.; Siefert, J.; Haynes, M.; Dinsdale, E.; Edwards, R.; Souza, V.; Rohwer, F.; Hollander, D. Metagenomic and stable isotopic analyses of modern freshwater microbialites in Cuatro Ciénegas, Mexico. Environ. Microbiol. 2009, 11, 16–34. [Google Scholar] [CrossRef] [PubMed]
- Nitti, A.; Daniels, C.A.; Siefert, J.; Souza, V.; Hollander, D.; Breitbart, M. Spatially resolved genomic, stable isotopic, and lipid analyses of a modern freshwater microbialite from Cuatro Ciénegas, Mexico. Astrobiology 2012, 12, 685–698. [Google Scholar] [CrossRef] [PubMed]
- Ruvindy, R.; White, R.A., III; Neilan, B.A.; Burns, B.P. Unravelling core microbial metabolisms in the hypersaline microbial mats of Shark Bay using high-throughput metagenomics. ISME J. 2016, 10, 183–196. [Google Scholar] [CrossRef]
- Kempe, S.; Kazmierczak, J.; Landmann, G.; Konuk, T.; Reimer, A.; Lipp, A. Largest known microbialites discovered in Lake Van, Turkey. Nature 1991, 349, 605–608. [Google Scholar] [CrossRef]
- Arp, G.; Reimer, A.; Reitner, J. Microbialite formation in seawater of increased alkalinity, Satonda Crater Lake, Indonesia. J. Sediment. Res. 2004, 74, 318–325. [Google Scholar] [CrossRef]
- White, R.A., III; Power, I.M.; Dipple, G.M.; Southam, G.; Suttle, C.A. Metagenomic analysis reveals that modern microbialites and polar microbial mats have similar taxonomic and functional potential. Front. Microbiol. 2015, 6, 966. [Google Scholar] [CrossRef]
- Brady, A.L.; Slater, G.F.; Omelon, C.R.; Southam, G.; Druschel, G.; Andersen, D.T.; Hawes, I.; Laval, B.; Lim, D.S.S. Photosynthetic isotope biosignatures in laminated micro-stromatolitic and non-laminated nodules associated with modern, freshwater microbialites in Pavilion Lake, BC. Chem. Geol. 2010, 274, 56–67. [Google Scholar] [CrossRef]
- White, R.A., III; Chan, A.M.; Gavelis, G.S.; Leander, B.S.; AL Brady, A.L.; Slater, G.F.; Lim, D.S.S.; Suttle, C.A. Metagenomic analysis suggests modern freshwater microbialites harbor a distinct core microbial community. Front. Microbiol. 2016, 6, 1531. [Google Scholar] [CrossRef]
- Thompson, J.; Ferris, F. Cyanobacterial Precipitation of Gypsum, Calcite, and Magnesite from Natural Alkaline Lake Water. Geology 1990, 18, 995–998. [Google Scholar] [CrossRef]
- Ludwig, R.; Al-Horani, F.; de Beer, D.; Jonkers, H. Photosynthesis-controlled calcification in a hypersaline microbial mat. Limnol. Oceanogr. 2005, 50, 1836–1843. [Google Scholar] [CrossRef]
- Altermann, W.; Kazmierczak, J.; Oren, A.; Wright, D. Cyanobacterial calcification and its rock-building potential during 3.5 billion years of Earth history. Geobiology 2006, 4, 147–166. [Google Scholar] [CrossRef]
- Bosak, T.; Greene, S.E.; Newman, D.K. A likely role for anoxygenic photosynthetic microbes in the formation of ancient stromatolites. Geobiology 2007, 5, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Power, I.M.; Wilson, S.A.; Thom, J.; Dipple, G.; Southam, G. Biologically induced mineralization of dypingite by cyanobacteria from an alkaline wetland near Atlin, British Columbia. Geochem. Trans. 2007, 8, 13. [Google Scholar] [CrossRef]
- Miller, A.G.; Colman, B. Evidence for HCO3− transport by the blue-green alga (cyanobacterium) Coccochloris Peniocystis. Plant Physiol. 1980, 65, 397–402. [Google Scholar] [CrossRef]
- Visscher, P.; Reid, R.; Bebout, B.; Hoeft, S.; Macintyre, I.; Thompson, J. Formation of lithified micritic laminae in modern marine stromatolites (Bahamas): The role of sulfur cycling. Am. Mineral. 1998, 83, 1482–1493. [Google Scholar] [CrossRef]
- Dupraz, C.; Visscher, P. Microbial lithification in marine stromatolites and hypersaline mats. Trends Microbiol. 2005, 13, 429–438. [Google Scholar] [CrossRef]
- Visscher, P.; Stolz, J. Microbial mats as bioreactors: Populations, processes, and products. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2005, 219, 87–100. [Google Scholar] [CrossRef]
- Merz, M.U.E. The biology of carbonate precipitation by cyanobacteria. Facies 1992, 26, 81–101. [Google Scholar] [CrossRef]
- Ferris, F.; Thompson, J.; Beveridge, T. Modern freshwater microbialites from Kelly Lake, British Columbia, Canada. Palaios 1997, 12, 213–219. [Google Scholar] [CrossRef]
- McConnaughey, T.; Burdett, J.; Whelan, J.; Paull, C. Carbon isotopes in biological carbonates: Respiration and photosynthesis. Geochim. Cosmochim. Acta 1997, 61, 611–622. [Google Scholar] [CrossRef]
- Sumner, D.Y. Microbial influences on local carbon isotopic ratios and their preservation in carbonate. Astrobiology 2001, 1, 57–70. [Google Scholar] [CrossRef] [PubMed]
- Kaźmierczak, J.; Fenchel, T.; Kühl, M.; Kempe, S.; Kremer, B.; Łącka, B.; Małkowski, K. CaCO3 precipitation in multilayered cyanobacterial mats: Clues to explain the alteration of micrite and sparite layers in calcareous stromatolies. Life 2015, 5, 744–769. [Google Scholar] [CrossRef] [PubMed]
- Belan, M.; Brady, A.L.; Kim, S.T.; Lim, D.S.S.; Slater, G.F. Spatial distribution and preservation of carbon isotope biosignatures in freshwater microbialite carbonate. ACS Earth Space Chem. 2019, 3, 335–343. [Google Scholar] [CrossRef]
- Chafetz, H.S.; Buczynski, C. Bacterially induced lithification of microbial mats. Palaios 1992, 7, 277–293. [Google Scholar] [CrossRef]
- Visscher, P.; Reid, R.; Bebout, B. Microscale observations of sulfate reduction: Correlation of microbial activity with lithified micritic laminae in modern marine stromatolites. Geology 2000, 28, 919–922. [Google Scholar] [CrossRef]
- White, D.C.; Davis, W.M.; Nickels, J.S.; King, J.D.; Bobbie, R.J. Determination of the sedimentary microbial biomass by extractible lipid phosphate. Oecologia 1979, 40, 51–62. [Google Scholar] [CrossRef]
- Green, C.; Scow, K. Analysis of phospholipid fatty acids (PLFA) to characterize microbial communities in aquifers. Hydrogeol. J. 2000, 8, 126–141. [Google Scholar] [CrossRef]
- Spring, S.; Schulze, R.; Overmann, J.; Schleifer, K. Identification and characterization of ecologically significant prokaryotes in the sediment of freshwater lakes: Molecular and cultivation studies. FEMS Microbiol. Rev. 2000, 24, 573–590. [Google Scholar] [CrossRef]
- Boschker, H.T.S.; Middelburg, J.J. Stable isotopes and biomarkers in microbial ecology. FEMS Microbiol. Ecol. 2002, 40, 85–95. [Google Scholar] [CrossRef]
- Hayes, J.M. Fractionation of carbon and hydrogen isotopes in biosynthetic processes. Rev. Minerol. Geochem. 2001, 43, 225–277. [Google Scholar] [CrossRef]
- Pancost, R.; Damste, J. Carbon isotopic compositions of prokaryotic lipids as tracers of carbon cycling in diverse settings. Chem. Geol. 2003, 195, 29–58. [Google Scholar] [CrossRef]
- Blair, N.; Leu, A.; Munoze Olsen, J.; Kwong, E.; Desmarais, D. Carbon Isotopic Fractionation in Heterotrophic Microbial-Metabolism. Appl. Environ. Microbiol. 1985, 50, 996–1001. [Google Scholar] [CrossRef] [PubMed]
- Sakata, S.; Hayes, J.; McTaggart, A.; Evans, R.; Leckrone, K.; Togasaki, R. Carbon isotopic fractionation associated with lipid biosynthesis by a cyanobacterium: Relevance for interpretation of biomarker records. Geochim. Cosmochim. Acta 1997, 61, 5379–5389. [Google Scholar] [CrossRef]
- Brady, A.L.; Laval, B.; Lim, D.S.S.; Slater, G.F. photoautotrophic and heterotrophic associated biosignatures in modern freshwater microbialites over seasonal and spatial gradients. Org. Geochem. 2014, 67, 8–18. [Google Scholar] [CrossRef]
- Russell, J.A.; Cardman, Z.; Brady, A.; Slater, G.; Lim, D.; Biddle, J.F. Prokaryote populations of extant microbialites along a depth gradient in Pavilion Lake, British Columbia, Canada. Geobiology 2014, 12, 250–264. [Google Scholar] [CrossRef]
- Lim, D.S.S.; Laval, B.E.; Slater, G.; Antoniades, D.; Forrest, A.L.; Pike, W.; Pieters, R.; Saffari, M.; Reid, D.; Schulze-Makuch, D.; et al. Limnology of Pavilion Lake, BC, Canada—Characterization of a microbialite forming environment. Fundam. Appl. Limnol. 2009, 173, 329–351. [Google Scholar] [CrossRef]
- Ferris, F.; Wiese, R.; Fyfe, W. Precipitation of Carbonate Minerals by Microorganisms—Implications for Silicate Weathering and the Global Carbon-Dioxide Budget. Geomicrobiol. J. 1994, 12, 1–13. [Google Scholar] [CrossRef]
- Pawlowicz, R. Calculating the Conductivity of Natural Waters. Limnol. Oceanogr. Methods 2008, 6, 489–501. [Google Scholar] [CrossRef]
- Bronk Ramsey, C. Bayesian analysis of radiocarbon dates. Radiocarbon 2009, 51, 337–360. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [PubMed]
- Guckert, J.; Antworth, C.; Nichols, P.; White, D. Phospholipid, Ester-Linked Fatty-Acid Profiles as Reproducible Assays for Changes in Prokaryotic Community Structure of Estuarine Sediments. FEMS Microbiol. Ecol. 1985, 31, 147–158. [Google Scholar] [CrossRef]
- Ryter, A.; Kellenberger, E. Etude au microscope electronique de plasmas contendant de l’acid desoxyribonucleique. I. Les nucltoides des bacteries en croissance active. Zeitschrqt Fur Nat. 1958, B13, 597–605. [Google Scholar] [CrossRef]
- Omelon, C.R.; Pollard, W.H.; Ferris, F.G. Chemical and ultrastructural characterization of high arctic cryptoendolithic habitats. Geomicrobiol. J. 2006, 23, 189–200. [Google Scholar] [CrossRef]
- Galland-Irmouli, A.V.; Pons, L.; Luçon, M.; Villaume, C.; Mrabet, N.T.; Guéant, J.L.; Fleurence, J. One-step purification of R-phycoerythrin from the red macroalga Palmaria palmate using preparative polyacrylamide gel electrophoresis. J. Chromatogr. B Biomed. Sci. Appl. 2000, 739, 117–123. [Google Scholar] [CrossRef]
- Papineau, D.; Walker, J.; Mojzsis, S.; Pace, N. Composition and structure of microbial communities from stromatolites of Hamelin Pool in Shark Bay, Western Australia. Appl. Environ. Microbiol. 2005, 71, 4822–4832. [Google Scholar] [CrossRef] [PubMed]
- Bordowitz, J.R.; Montgomery, B.L. Exploiting the autofluorescent properties of photosynthetic pigments for analysis of pigmentation and morphology in live Fremyella diplosiphon cells. Sensors 2010, 10, 6969–6979. [Google Scholar] [CrossRef]
- Fierer, N.; Jackson, J.A.; Vilgalys, R.; Jackson, R.B. Assessment of soil microbial community structure by use of taxon-specific quantitative PCR assays. Appl. Environ. Microbiol. 2005, 71, 4117–4120. [Google Scholar] [CrossRef]
- Waldrop, M.P.; Wickland, K.; Berhe, A.; White, R., III; Harden, J.; Romanovsky, V. Molecular investigations into a globally important carbon pool: Permafrost-protected carbon in Alaskan soils. Glob. Chang. Biol. 2010, 16, 2543–2554. [Google Scholar] [CrossRef]
- Nübel, U.; Garcia Pichel, F.; Muyzer, G. PCR primers to amplify 16S rRNA genes from cyanobacteria. Appl. Environ. Microbiol. 1997, 63, 3327–3332. [Google Scholar] [CrossRef]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE guidelines: Minimum information for publication of quantitative real-time PCR experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef]
- Brady, A.L.; Slater, G.; Laval, B.; Lim, D.S. Constraining carbon sources and growth rates of freshwater microbialites in Pavilion Lake using 14C analysis. Geobiology 2009, 7, 544–555. [Google Scholar] [CrossRef] [PubMed]
- O’Leary, M.H. Carbon isotopes in photosynthesis. Bioscience 1988, 38, 328–336. [Google Scholar] [CrossRef]
- Franzmann, P.D.; Patterson, B.M.; Power, T.R.; Nichols, P.D.; Davis, G.B. Microbial biomass in a shallow, urban aquifer contaminated with aromatic hydrocarbons: Analysis by phospholipid fatty acid content and composition. J. Appl. Bacteriol. 1996, 80, 617–625. [Google Scholar] [CrossRef]
- Volkman, J.K.; Johns, R.B.; Gillan, F.T.; Perry, G.J.; Bavor, H.J. Microbial lipids of an intertidal sediment-I. Fatty acids and hydrocarbons. Geochim. Cosmochim. Acta 1980, 44, 1133–1143. [Google Scholar] [CrossRef]
- Buhring, S.; Smittenberg, R.; Sachse, D.; Lipp, J.; Golubic, S.; Sachs, J.; Hinrichs, K.U.; Summons, R. A hypersaline microbial mat from the Pacific Atoll Kiritimati: Insights into composition and carbon fixation using biomarker analyses and a 13C-labeling approach. Geobiology 2009, 7, 1–16. [Google Scholar] [CrossRef]
- White, D.C. Validation of quantitative analysis for microbial biomass, community structure, and metabolic activity. Adv. Limnol. 1988, 31, 1–18. [Google Scholar]
- Jahnke, L.L.; Embaye, T.; Hope, J.; Turk, K.A.; Van Zuilen, A.; Des Marais, D.J.; Farmer, J.D.; Summons, R.E. Lipid biomarker and carbon isotopic signatures for stromatolite-forming, microbial mat communities and Phormidium cultures from Yellowstone National Park. Geobiology 2004, 2, 31–47. [Google Scholar] [CrossRef]
- Abraham, W.R.; Hesse, C.; Pelz, O. Ratios of carbon isotopes in microbial lipids as an indicator of substrate usage. Appl. Environ. Microbiol. 1998, 64, 4202–4209. [Google Scholar] [CrossRef]
- Teece, M.A.; Fogel, M.L.; Dollhopf, M.E.; Nealson, K.H. Isotopic fractionation associated with biosynthesis of fatty acids by a marine bacterium under oxic and anoxic conditions. Org. Geochem. 1999, 30, 1571–1579. [Google Scholar] [CrossRef]
- Romanek, C.S.; Grossman, E.L.; Morse, J.W. Carbon isotopic fractionation in synthetic aragonite and calcite: Effects of temperature and precipitation rate. Geochim. Cosmochim. Acta 1992, 56, 419–430. [Google Scholar] [CrossRef]
- Thompson, J.; SchultzeLam, S.; Beveridge, T.; DesMarais, D. Whiting events: Biogenic origin due to the photosynthetic activity of cyanobacterial picoplankton. Limnol. Oceanogr. 1997, 42, 133–141. [Google Scholar] [PubMed]
- Vestal, J.R.; White, D.C. Lipid analysis in microbial ecology. BioScience 1989, 39, 535–541. [Google Scholar] [CrossRef]
- Zelles, L. Phospholipid fatty acid profiles in select members of soil microbial communities. Chemosphere 1997, 35, 275–294. [Google Scholar] [CrossRef]
- Kenyon, C.N. Fatty acid composition of unicellular strains of blue-green algae. J. Bacteriol. 1972, 109, 827–834. [Google Scholar] [CrossRef]
- Grimalt, J.O.; de Wit, R.; Teixidor, P.; Albaiges, J. Lipid biogeochemistry of Phormidium and Microcoleus mats. Org. Geochem. 1992, 19, 509–530. [Google Scholar] [CrossRef]
- Park, C.; Berger, L.R. Complex lipids of Rhodomicrobium vannielii. J. Bacteriol. 1967, 93, 221–229. [Google Scholar] [CrossRef]
- Bobbie, R.J.; White, D.C. Characterization of Benthic Microbial Community Structure by High-Resolution Gas-Chromatography of Fatty-Acid Methyl-Esters. Appl. Environ. Microbiol. 1980, 39, 1212–1222. [Google Scholar] [CrossRef]
- Findlay, R.H.; Trexler, M.B.; Guckert, J.B.; White, D.C. Laboratory Study of Disturbance in Marine-Sediments—Response of a Microbial Community. Mar. Ecol. Prog. Ser. 1990, 62, 121–133. [Google Scholar] [CrossRef]
- Cavigelli, M.A.; Robertson, G.P.; Klug, M.J. Fatty-Acid Methyl-Ester (Fame) Profiles as Measures of Soil Microbial Community Structure. Plant Soil 1995, 170, 99–113. [Google Scholar] [CrossRef]
- Harwood, J.L.; Guschina, I.A. The versatility of algae and their lipid metabolism. Biochimie 2009, 91, 679–684. [Google Scholar] [CrossRef] [PubMed]
- Kaur, A.; Chaudhary, A.; Kaur, A.; Choudhary, R.; Kaushik, R. Phospholipid fatty acid—A bioindicator of environmental monitoring and assessment in soil ecosystems. Curr. Sci. 2005, 89, 1103–1113. [Google Scholar]
- Kaneda, T. Iso-and anteiso-fatty acids in bacteria: Biosynthesis, function, and taxonomic significance. Microbiol. Rev. 1991, 55, 288–302. [Google Scholar] [CrossRef] [PubMed]
- Wakeham, S.G. Lipid biomarkers for heterotrophic alteration of suspended particulate organic matter in oxygenated and anoxic water columns of the ocean. Deep-Sea Res. Part I Oceanogr. Res. Pap. 1995, 42, 1749–1771. [Google Scholar] [CrossRef]
- Fang, J.; Hasiotis, S.T.; Das Gupta, S.; Brake, S.S.; Bazylinski, D.A. Microbial biomass and community structure of a stromatolite from an acid mine drainage system as determined by lipid analysis. Chem. Geol. 2007, 243, 191–204. [Google Scholar] [CrossRef]
- O’Leary, W.M.; Wilkinson, S.G. Gram-positive bacteria. In Microbial Lipids; Academic Press: London, UK, 1988; pp. 117–201. [Google Scholar]
- Laval, B.; Cady, S.L.; Pollack, J.C.; McKay, C.P.; Bird, J.S.; Grotzinger, J.P.; Ford, D.C.; Bohm, H.R. Modern freshwater microbialite analogues for ancient dendritic reef structures. Nature 2000, 407, 626–629. [Google Scholar] [CrossRef]
- Omelon, C.; Brady, A.; Slater, G.; Laval, B.; Lim, D.; Southam, G. Microstructure variability in freshwater microbialites, Pavilion Lake, Canada. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2013, 392, 62–70. [Google Scholar] [CrossRef]
- Navarrete, A.; Peacock, A.; Macnaughton, S.J.; Urmeneta, J.; Mas-Castella, J.; White, D.C.; Guerrero, R. Physiological status and community composition of microbial mats of the Ebro Delta, Spain, by signature lipid biomarkers. Microb. Ecol. 2000, 39, 92–99. [Google Scholar] [CrossRef][Green Version]
- Klappenbach, J.A.; Dunbar, J.M.; Schmidt, T.M. rRNA operon copy number reflects ecological strategies of bacteria. Appl. Environ. Microbiol. 2000, 66, 1328–1333. [Google Scholar] [CrossRef]
- Smith, C.J.; Osborn, A.M. Advantages and limitations of quantitative PCR (Q-PCR)-based approaches in microbial ecology. FEMS Microbiol. Ecol. 2009, 67, 6–20. [Google Scholar] [CrossRef]
- Gonzalez, J.M.; Portillo, M.C.; Belda-Ferre, P.; Mira, A. Amplification by PCR Artificially Reduces the Proportion of the Rare Biosphere in Microbial Communities. PLoS ONE 2012, 7, e29973. [Google Scholar] [CrossRef] [PubMed]
- Dolman, A.M.; Ruecker, J.; Pick, F.R.; Fastner, J.; Rohrlack, T.; Mischke, U.; Wiedner, C. Cyanobacteria and Cyanotoxins: The Influence of Nitrogen versus Phosphorus. PLoS ONE 2012, 7, e38757. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.C.Y.; Kuwabara, J.S.; Pasilis, S.P. Phosphate and iron limitation of phytoplankton biomass in Lake Tahoe. Can. J. Fish. Aquat. Sci. 1992, 49, 1206–1217. [Google Scholar] [CrossRef]
- Zohary, T.; Robarts, R.D. Experimental study of microbial P limitation in the eastern Mediterranean. Limnol. Oceanogr. 1998, 43, 387–395. [Google Scholar] [CrossRef]
- Smith, E.M.; Prairie, Y.T. Bacterial metabolism and growth efficiency in lakes: The importance of phosphorus availability. Limnol. Oceanogr. 2004, 49, 137–147. [Google Scholar] [CrossRef]
- Schulze-Makuch, D.; Lim, D.; Laval, B.; Turse, C.; António, M.; Chan, O.; Pointing, S.B.; Brady, A.; Reid, D.; Irwin, L.N. Pavilion Lake microbialites: Morphological, molecular and biochemical evidence for a cold-water transition to colonial aggregates. Life 2013, 3, 21–37. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Product Size (Bp) | Annealing Temp (°C) | Reference |
|---|---|---|---|
| Eub338f ACTCCTACGGGAGGCAGCAG Eub518r ATTACCGCGGCTGCTGG Cya106f CGGACGGGTGAGTAACGCGTGA Cya359r * CCCATTGCGGAAAATTCCC | 200 | 53 | Fierer et al. [48] |
| 265 | 60 | Nübel et al. [50] |
| Sample | [DIC] (mg/L) | δ13CDIC (‰) | pH | Conductivity (µs/cm) | [P] (µg/L) |
|---|---|---|---|---|---|
| Kelly surface water | 36.5 | −5.7 *,‡ | 8.41 | 326 | 2 |
| Kelly deepwater (15 m) | N/A | −6.7 * | N/A | N/A | N/A |
| Kelly deepwater (30 m) † | 37.8 | −7.2 * | 8.28 | 334 | 4 |
| Porcupine creek | 40.7 | −7.3 | 8.27 | 365 | 6 |
| Jim well † | 41.7 | −10.1 | 7.83 | 422 | 6 |
| Guy and Ursula well † | 48.6 | −10.1 | 7.71 | 626 | 7 |
| Sample Depth | [PLFAs] (µg/g Microbialite) | [PLFAs] (mg/g TOC) | δ13Ccarb ‰ (PBD) | δ13Corg ‰ (PDB) |
|---|---|---|---|---|
| 11 m | 7.2 ± 4.4 | 0.4 ± 0.2 | −2.7 ± 0.5 | −29.8 ± 0.8 |
| 14 m | 24.5 ± 11.0 | 0.8 ± 0.3 | −3.8 ± 0.1 | −27.6 ± 2.3 |
| 20 m | 51.5 ± 12.7 | 1.3 ± 0.1 | −3.8 ± 0.5 | −30.8 ± 1.6 |
| 26 m | 35.7 ± 15.6 | 1.4 ± 0.5 | −0.3 ± 0.5 | −34.2 ± 0.7 |
| 11 m | 14 m | 20 m | 26 m | |||||
|---|---|---|---|---|---|---|---|---|
| PLFA I.D. | mol % | δ13C (‰) | mol % | δ13C (‰) | mol % | δ13C (‰) | mol % | δ13C (‰) |
| br14:0 | 0.8 | 1.5 | 0.3 | 0.5 | ||||
| 14:1 | 0.4 | 0.0 | 0.3 | 0.8 | ||||
| 14:0 | 6.0 | −36.6 ± 0.5 | 5.4 | − 43.7 ± 0.5 | 5.0 | −44.3 ± 0.5 | 7.8 | −48.3 ± 0.5 |
| br15:0 | 0.0 | 0.0 | 0.0 | 0.4 | ||||
| i-15:0 | 4.5 | −32.5 ± 0.5 | 5.3 | −32.7 ± 0.6 | 1.4 | −34.2 ± 0.5 | 1.6 | −35.0 ± 0.5 |
| a-15:0 | 3.7 | −35.1 ± 0.5 | 4.7 | −31.8 ± 0.5 | 1.1 | −35.0 ± 0.5 | 1.5 | −34.0 ±0.5 |
| 15:0 | 1.4 | 1.2 | 0.4 | 0.5 | ||||
| br16:0 | 2.1 | 1.3 | 0.0 | 0.0 | ||||
| 16:3 | 0.0 | 0.0 | 1.4 | 1.8 | ||||
| 16:2 | 0.0 | 0.0 | 2.1 | 2.0 | ||||
| 16:1 | 11.6 | −42.0 ± 0.6 | 15.6 | −39.4 ± 0.8 | 14.2 | −41.6 ± 1.5 | 11.0 | −44.1 ± 0.5 |
| 16:0 | 29.7 | −38.0 ± 0.5 | 27.1 | −42.5 ± 0.7 | 25.2 | −42.1 ± 0.5 | 24.8 | −46.8 ± 0.5 |
| br17:1 | 0.0 | 0.6 | 0.5 | 0.2 | ||||
| 10me16:0 | 3.3 | 5.3 | 1.6 | 1.5 | ||||
| i-17:0 | 1.5 | 0.2 | 0.4 | 0.1 | ||||
| a-17:0 | 1.6 | 0.5 | 0.5 | 0.2 | ||||
| 17:1 | 0.0 | 0.0 | 0.4 | 0.2 | ||||
| cy17:0 | 1.9 | 1.2 | 0.6 | 0.4 | ||||
| 17:0 | 1.2 | 0.0 | 0.3 | 0.1 | ||||
| br18:0 | 0.0 | 0.0 | 0.2 | 0.0 | ||||
| 18:3 | 3.1 | 0.3 | −42.9 ± 0.7* | 2.9 | −45.1 ± 0.6 | 1.8 | −47.3 ± 0.5* | |
| 18:2 | 6.4 | 6.3 | 10.7 | −45.7 ± 0.5 | 9.0 | |||
| 18:1 | 15.1 | 17.2 | 21.1 | −43.3 ±0.5 | 25.4 | |||
| 18:0 | 3.1 | 1.5 | 1.4 | 0.8 | ||||
| br19:0 | 1.2 | 0.5 | 0.4 | 0.4 | ||||
| cy19:0 | 1.6 | 1.4 | −33.8 ± 0.6 | 0.7 | 0.6 | |||
| 20:4 | 0.0 | 0.8 | 1.5 | 1.7 | −47.9 ± 0.6† | |||
| 20:5 | 0.0 | 1.0 | 1.0 | 2.0 | ||||
| 20:3 | 0.0 | 0.0 | 0.6 | 0.7 | ||||
| 20:2 | 0.0 | 0.0 | 0.3 | 0.2 | ||||
| 20:1 | 0.0 | 0.6 | 1.3 | 0.9 | ||||
| 20:0 | 0.0 | 0.5 | 0.3 | 0.2 | ||||
| 22:6 | 0.0 | 0.0 | 0.2 | 0.0 | ||||
| 22:5 | 0.0 | 0.0 | 0.2 | 0.2 | ||||
| 22:1 | 0.0 | 0.0 | 0.4 | 0.0 | ||||
| 22:0 | 0.0 | 0.0 | 0.2 | 0.1 | ||||
| 24:1 | 0.0 | 0.0 | 0.2 | 0.0 | ||||
| 24:0 | 0.0 | 0.0 | 0.2 | 0.1 | ||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
White, R.A., III; Soles, S.A.; Brady, A.L.; Southam, G.; Lim, D.S.S.; Slater, G.F. Biosignatures Associated with Freshwater Microbialites. Life 2020, 10, 66. https://doi.org/10.3390/life10050066
White RA III, Soles SA, Brady AL, Southam G, Lim DSS, Slater GF. Biosignatures Associated with Freshwater Microbialites. Life. 2020; 10(5):66. https://doi.org/10.3390/life10050066
Chicago/Turabian StyleWhite, Richard Allen, III, Sarah A. Soles, Allyson L. Brady, Gordon Southam, Darlene S.S. Lim, and Greg F. Slater. 2020. "Biosignatures Associated with Freshwater Microbialites" Life 10, no. 5: 66. https://doi.org/10.3390/life10050066
APA StyleWhite, R. A., III, Soles, S. A., Brady, A. L., Southam, G., Lim, D. S. S., & Slater, G. F. (2020). Biosignatures Associated with Freshwater Microbialites. Life, 10(5), 66. https://doi.org/10.3390/life10050066