Abstract
Recently, a 30-cm object was discovered to graze the Earth’s atmosphere and shift into a Jupiter-crossing orbit. We use the related survey parameters to calibrate the total number of such objects. The number of objects that could have exported terrestrial microbes out of the Solar System is in the range –. We find that – such objects could have been captured by binary star systems over the lifetime of the Solar System. Adopting the fiducial assumption that one polyextremophile colony is picked up by each object, the total number of objects carrying living colonies on them upon capture could be 10–.
1. Introduction
Panspermia is the conjecture that life can propagate from one planet to another [1,2]. One version of panspermia involves Solar System bodies grazing the Earth’s atmosphere, picking up microbes, and being ejected from the Solar System. About ∼1–60 long-period comets and interstellar objects could have to have undergone such a process over the lifetime of the Solar System [3]. It has also been shown that ejected objects can be gravitationally captured by other star systems [4,5].
Recently, Shober et al. [6] reported a detection by the Desert Fireball Network (DFN) (http://fireballsinthesky.com.au/) of a ∼30- object that reached a minimum altitude ∼58.5 in the Earth’s atmosphere during a grazing event, and was transferred from an Apollo-type orbit into a Jupiter-family comet orbit, making it likely to be ejected from the Solar System during a future gravitational encounter with Jupiter. This detection represents a new class of objects that can pick up life in the Earth’s atmosphere before being ejected from the Solar System: rocky, inner-Solar System bodies that are scattered into Jupiter-crossing orbits after their grazing interactions with Earth and are subsequently ejected. These objects are important, since, based on their characteristically higher densities relative to icy, outer-Solar System bodies, they can survive passes through the Earth’s atmosphere at significantly smaller sizes.
Here, we study the likelihood of life-bearing Solar System bodies being captured by exoplanetary systems. First, we discuss the collection of microbial life during the transporting body’s passage through the atmosphere as well as the survival lifetime in space. Next, we use the detection be Shober et al. [6] to calibrate the number of similar objects that could have picked up life in the Earth’s atmosphere and were likely ejected from the Solar System. We then use a Monte Carlo simulation to estimate the total number of such objects that were captured by stellar binary systems over the lifetime of the Solar System, as well as the number of these objects that likely harbor living microbes at the time of capture. Finally, we summarize our main conclusions.
2. Collection of Microbial Life
A 30-cm object grazing the atmosphere for 90 s at a speed of should collect ∼ microbial colonies during its trip through the atmosphere, based on Equation (5) from [3] for the number of colonies collected,
where d is the object diameter, v is the speed, and is the time spent in the atmosphere.
It has been demonstrated that many microbes can likely survive the accelerations associated with transfer onto the grazing object [7,8], although specific detections in the atmosphere linked to microbes that have undergone such laboratory testing have not yet been made.
Polyextremophiles such as Deinococcus radiodurans are estimated to die on an exponential timescale of ∼ with minimal radiation shielding [4,9,10,11]. While the abundance of polyextremophiles in the atmosphere is yet unconstrained, in our analysis, we assume a fiducial estimate of 1 polyextremophile colony (≲10−4 of incident microbe colonies) per transporting object. The interiors of objects with diameters greater than 20 cm are not expected to be heated to more than C during a passage through the atmosphere [12], and many asteroids are known to have significant porosity [13,14], thus we adopt a model in which incident microbes become lodged inside the object and shielded from the exterior heating. We note that this assumption is not critical; increasing the minimum altitude by a few scale heights above the observed altitude of the Shober et al. [6] bolide would result in minimal heating (<100 °C) while only reducing the atmospheric cross section by a small factor, potentially allowing for microbes on the surface in the case of non-porous objects.
3. Number of Ejected Objects
The DFN uses ∼50 cameras to cover ∼2.5 of the sky, or ∼5 of the Earth’s surface [15]. While the DFN only had four cameras in 2007, we conservatively estimate the relevant observation parameters for the Shober et al. [6] detection to be the result of a survey that lasted ∼10 years and covered ∼5 of the Earth’s surface. This leads to a rate estimate of ∼20 of 30-cm objects penetrating the Earth’s atmosphere and eventually likely being ejected from the Solar System, with 95% Poisson bounds of –. Over , this translates to ∼6 ejected objects, with 95% Poisson bounds of –.
We use Equation (9) from [16],
to estimate the yield strength of the [6] meteor to be ∼6 , given its density estimate of 3500 . We then use Equation (10) from [16],
where is the density of air at altitude z, and the known speed of to estimate that the altitude of breakup as 48 , which is ∼ below the observed minimum altitude of the Shober et al. [6] meteor. Breakup is therefore not considered to be a significant factor in this analysis for typical rocky compositions, although it is noted that the Shober et al. [6] meteor experienced a breakup event, which may be indicative of a composite makeup.
The drag force on the object can be written as,
where is the drag coefficient, taken to be unity for typical meteor speeds; m and v are the mass and speed of the object; and A is the cross-sectional area of the object. We adopt the limit of constant mass, assume , and require that the maximum change in speed is , so that the object will possess more than the escape speed from the Earth and not fall to the ground. Over 90 s of travel in the atmosphere, the latter constraint can be expressed as a maximum acceleration, . We then find that,
Adopting 3500 , and calculating the mean density of air over the Shober et al. [6] meteor’s 90 s trajectory to be , the constraint becomes,
This implies that objects with diameters smaller than , or the size of the Shober et al. [6] meteor, slowed significantly by friction, although this estimate depends on the shape and mean density to within a factor of a few. We therefore do not consider objects smaller than .
We estimate the total number of transporting objects over the Earth’s lifetime as the aforementioned values, calibrated by the Shober et al. [6] meteor is ∼6 , with 95% Poisson bounds of –.
4. Number of Captured Objects
We assume that most objects are ejected at near-zero speeds from the Solar System, mirroring the Sun’s motion through the Local Standard of Rest (LSR), [17]. We adopt the following three-dimensional velocity dispersions for local stars, each corresponding to the standard deviation of a Gaussian distribution about the LSR: , , and [17].
Tight stellar binary systems have the largest capture cross sections, thus we exclusively consider these systems in this analysis [4,18]. We apply the scalings in [18,19] to estimate the capture cross section of a stellar binary system of two solar-mass stars (such as Alpha Centauri A and B) as a function of relative speed of the system and the object, assuming that the orbital speed of the bound orbit is near zero,
where is the relative encounter speed in units of and is the semi-major axis of this binary system.
The local number density of solar-type stars is estimated to be [20]. About 20% of solar-type stars are in equal-mass binary systems with [21,22], thus we estimate the number density of solar-type close binaries to be 1.6 . The solar-type binary period distribution is approximately log-normal, peaking at log(P/day) = 4.8 with dispersion of 2.3 orders of magnitude [23,24,25]. Given our assumption that ejected objects mirror the Sun’s speed relative to the LSR, we estimate that each object encounters a binary system every ∼5 .
5. Monte Carlo Simulation
We use the following Monte Carlo method to determine the capture rate of life-bearing Solar System bodies. First, we draw randomly from the Gaussian distributions of local stars, centered around the LSR, to determine the relative speed of the object with the binary system it encounters. Next, we draw the binary period from the aforementioned probability distribution, and, assuming a circular orbit, we use Kepler’s third law to translate the period to a semi-major axis length. We use the speed and semi-major axis to determine the capture cross section of the system, and then use the capture cross section and the assumption of to estimate the capture probability, . We apply the probability to determine whether or not the object is captured. If the object is captured, we record the time of capture, the relative speed, and the semi-major axis of the binary system. If the object is not captured, we step to the next encounter and repeat the process. We ran our code for particles over years, resulting in a capture probability of per encounter. The distributions of v and a for the captured objects are shown in Figure 1 and Figure 2, indicating that binaries with smaller semi-major lead to higher incidences of capture and that the relative speed that leads to the highest chance of capture is within –.
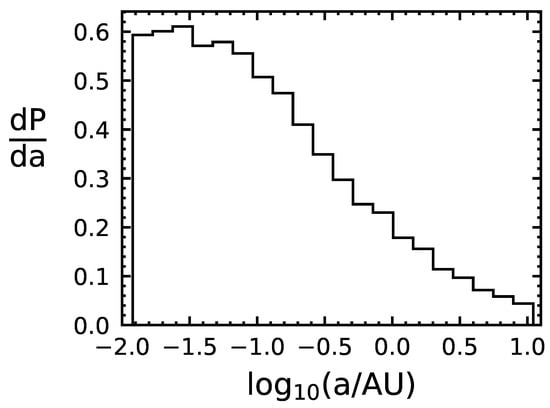
Figure 1.
Distribution of semi-major axes of capturing binary systems for simulated captured objects. P is probability and a is the semi-major axis of the binary.
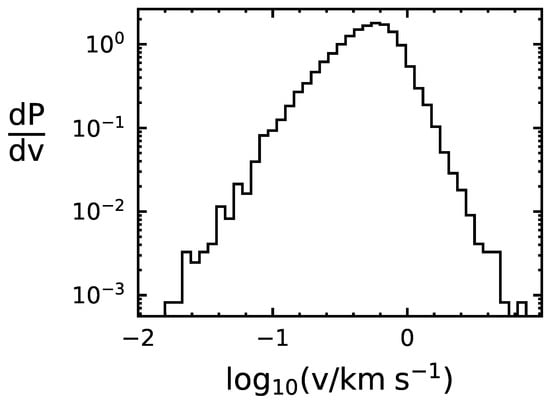
Figure 2.
Distribution of relative speed at capture between simulated captured objects and their respective binary systems. P is probability and v is the relative speed at capture.
Significant slow-down for centimeter-scale objects of density ∼3500 travelling through the ISM occurs when the accumulated ISM mass is comparable to the mass of the object [26],
where is the proton mass; is the mean proton number density of the ISM, taken to be ; L is the distance travelled; A is the cross-sectional area of the object; and is the mass of the object. For the aforementioned values, we find that the timescale on which significant slow-down occurs is several times longer than the age of the Universe. We therefore disregard slow-down effects.
Over 3 Gyr, an order unity assumption for the timescale on which life has likely existed in the atmosphere, the single-encounter capture probability of ∼6.4 leads to a total capture probability of ∼4 , or ∼2 objects, with 95% Poisson bounds of –. By integrating the expression for the surviving proportion of polyextremophile colonies over a time t, , multiplied by the constant single-encounter capture rate, we find that ∼4 of all objects were captured and hosted a living polyextremophile colony at the time of capture, assuming each object obtained one polyextremophile colony after interacting with the Earth’s atmosphere. We therefore estimate that ∼200 potentially life-bearing Solar System bodies could have been captured by exoplanetary systems over the lifetime of the Solar System while there still may have possibly been living microbes on the object, with 95% Poisson bounds of 10–1000 objects.
6. Conclusions
We calculated the likelihood of potentially life-bearing Solar System bodies to get captured by exoplanetary systems. The total number of objects captured by exoplanetary systems over the lifetime of the Solar System is –, with the total number of objects with the possibility of living microbes on them at the time of capture estimated to be 10–1000.
Panspermia is a multi-stage process and not all steps are addressed here, most notably the delivery of microbes to a planet within binary systems. Many of the probability estimates derived here are related to explicitly stated fiducial values, and are easily scaled to accommodate future research. Further detections of Earthgrazing objects transferred into Jupiter-crossing orbits will allow for more precise estimates of this process. Measurements of the abundance of polyextremophiles in the atmosphere will also refine the estimates made in this paper. Additional studies of polyextremophiles in space will advance the biological understanding of this panspermia channel.
Author Contributions
Conceptualization, A.S. and A.L.; methodology, A.S. and A.L.; A.S. and A.L.; formal analysis, A.S. and A.L.; investigation, A.S. and A.L.; writing–original draft preparation, A.S.; writing–review and editing, A.L.; funding acquisition, A.L. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported in part by a grant from the Breakthrough Prize Foundation.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Wesson, P.S. Panspermia, Past and Present: Astrophysical and Biophysical Conditions for the Dissemination of Life in Space. Space Sci. Rev. 2010, 156, 239–252. [Google Scholar] [CrossRef]
- Wickramasinghe, C. The astrobiological case for our cosmic ancestry. Space Sci. Rev. 2010, 9, 119–129. [Google Scholar] [CrossRef]
- Siraj, A.; Loeb, A. Exporting Terrestrial Life Out of the Solar System with Gravitational Slingshots of Earthgrazing Bodies. arXiv 2019, arXiv:1910.06414. [Google Scholar] [CrossRef]
- Ginsburg, I.; Lingam, M.; Loeb, A. Galactic Panspermia. Astrophys. J. 2018, 868, L12. [Google Scholar] [CrossRef]
- Lingam, M.; Loeb, A. Implications of Captured Interstellar Objects for Panspermia and Extraterrestrial Life. Astron. J. 2018, 156, 193. [Google Scholar] [CrossRef]
- Shober, P.M.; Jansen-Sturgeon, T.; Sansom, E.K.; Devillepoix, H.A.R.; Towner, M.C.; Bland, P.A.; Cup, M.; Howie, R.M.; Hartig, B.A.D. Where Did They Come From, Where Did They Go. Grazing Fireballs. Astron. J. 2020, 159, 191. [Google Scholar] [CrossRef]
- Deguchi, S.; Shimoshige, H.; Tsudome, M.; Mukai, S.; Corkery, R.W.; Ito, S.; Horikoshi, K. Microbial growth at hyperaccelerations up to 403,627 × g. Proc. Natl. Acad. Sci. USA 2011, 108, 7997–8002. [Google Scholar] [CrossRef]
- Mastrapa, R.M.E.; Glanzberg, H.; Head, J.N.; Melosh, H.J.; Nicholson, W.L. Survival of bacteria exposed to extreme acceleration: implications for panspermia. Earth Planet. Sci. Lett. 2001, 189, 1–8. [Google Scholar] [CrossRef]
- Burchell, M.J. Panspermia today. Int. J. Astrobiol. 2004, 3, 73–80. [Google Scholar] [CrossRef]
- Mileikowsky, C.; Cucinotta, F.A.; Wilson, J.W.; Gladman, B.; Horneck, G.; Lindegren, L.; Melosh, J.; Rickman, H.; Valtonen, M.; Zheng, J.Q. Natural Transfer of Viable Microbes in Space. 1. From Mars to Earth and Earth to Mars. Icarus 2000, 145, 391–427. [Google Scholar] [CrossRef]
- Phillips, R.; Kondev, J.; Theriot, J.; Garcia, H.G.; Orme, N. Physical Biology of the Cell.The Quarterly Review of Biology. Phys. Biol. Cell Garland Sci. 2013, 142. [Google Scholar] [CrossRef]
- Napier, W.M. A mechanism for interstellar panspermia. MNRAS 2004, 348, 46–51. [Google Scholar] [CrossRef]
- Britt, D.T.; Consolmagno, G.J. The Porosity of Dark Meteorites and the Structure of Low-Albedo Asteroids. Icarus 2000, 146, 213–219. [Google Scholar] [CrossRef]
- Britt, D.T.; Consolmagno, G.J.; Merline, W.J. Small Body Density and Porosity: New Data, New Insights. In Proceedings of the 37th Annual Lunar and Planetary Science Conference, League, TX, USA, 13–17 March 2006; p. 2214. [Google Scholar]
- Devillepoix, H.A.R.; Bland, P.A.; Towner, M.C.; Cup, M.; Sansom, E.K.; Jansen-Sturgeon, T.; Howie, R.M.; Paxman, J.; Hartig, B.A.D. Status of the Desert Fireball Network. In Proceedings of the International Meteor Conference, Egmond, The Netherlands, 2–5 June 2016; p. 60. [Google Scholar]
- Collins, G.S.; Jay, H.M.; Marcus, R.A. Earth Impact Effects Program: A Web-based computer program for calculating the regional environmental consequences of a meteoroid impact on Earth. Meteorit. Planet. Sci. 2010, 40, 817. [Google Scholar] [CrossRef]
- Bland-Hawthorn, J.; Gerhard, O. The Galaxy in Context: Structural, Kinematic, and Integrated Properties. Annu. Rev. Astron. Astrophys. 2016, 54, 529–596. [Google Scholar] [CrossRef]
- Valtonen, M.J. On the capture of comets into the Solar System. Observatory 1983, 103, 1–4. [Google Scholar]
- Valtonen, M.J.; Innanen, K.A. The capture of interstellar comets. Astrophys. J. 1982, 255, 307–315. [Google Scholar] [CrossRef]
- Bovy, J. Stellar inventory of the solar neighbourhood using Gaia DR1. MNRAS 2017, 470, 1360–1387. [Google Scholar] [CrossRef]
- Moe, M.; Di Stefano, R. Mind Your Ps and Qs: The Interrelation between Period (P) and Mass-ratio (Q) Distributions of Binary Stars. Astrophys. J. Suppl. Ser. 2017, 230, 15. [Google Scholar] [CrossRef]
- Moe, M.; Kratter, K.M.; Badenes, C. The Close Binary Fraction of Solar-type Stars Is Strongly Anticorrelated with Metallicity. Astrophys. J. 2019, 875, 61. [Google Scholar] [CrossRef]
- Duquennoy, A.; Mayor, M. Multiplicity among solar-type stars in the solar neighbourhood. II - Distribution of the orbital elements in an unbiased sample. Astron. Astrophys. 1991, 500, 337–376. [Google Scholar]
- Raghavan, D.; McAlister, H.A.; Henry, T.J.; Latham, D.W.; Marcy, G.W.; Mason, B.D.; Gies, D.R.; White, R.J.; ten Brummelaar, T.A. A Survey of Stellar Families: Multiplicity of Solar-type Stars. Astrophys. J. Suppl. Ser. 2010, 190, 1–42. [Google Scholar] [CrossRef]
- Tokovinin, A. From Binaries to Multiples. I. Data on F and G Dwarfs within 67 pc of the Sun. Astron. J. 2014, 147, 86. [Google Scholar] [CrossRef]
- Bialy, S.; Loeb, A. Could Solar Radiation Pressure Explain ‘Oumuamua’s Peculiar Acceleration? Astrophys. J. 2018, 868, L1. [Google Scholar] [CrossRef]


© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).