
Microbiological Advances in Biohydrometallurgy

Abstract
:1. Introduction
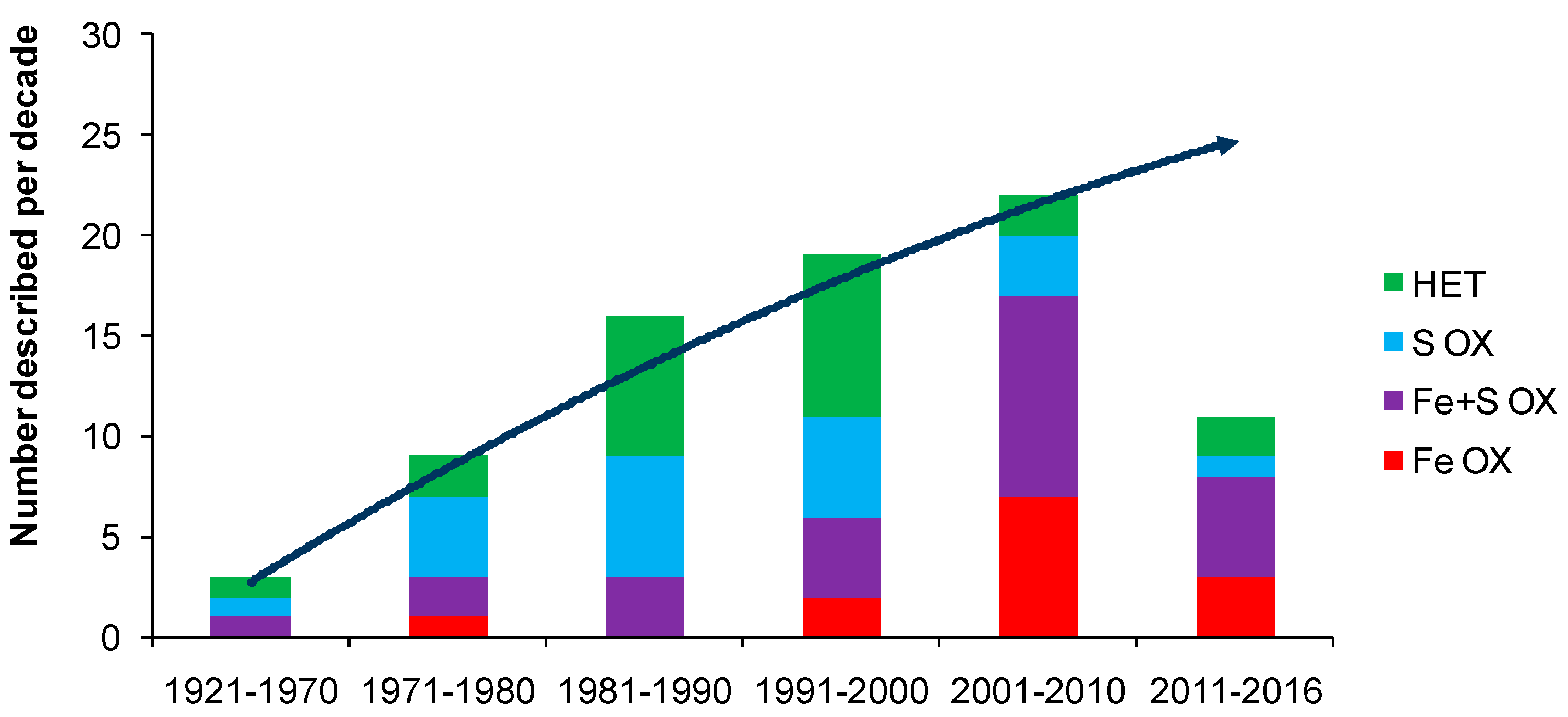
2. Microbiological Discovery
3. Microbial Responses to Extreme and Variable Habitats
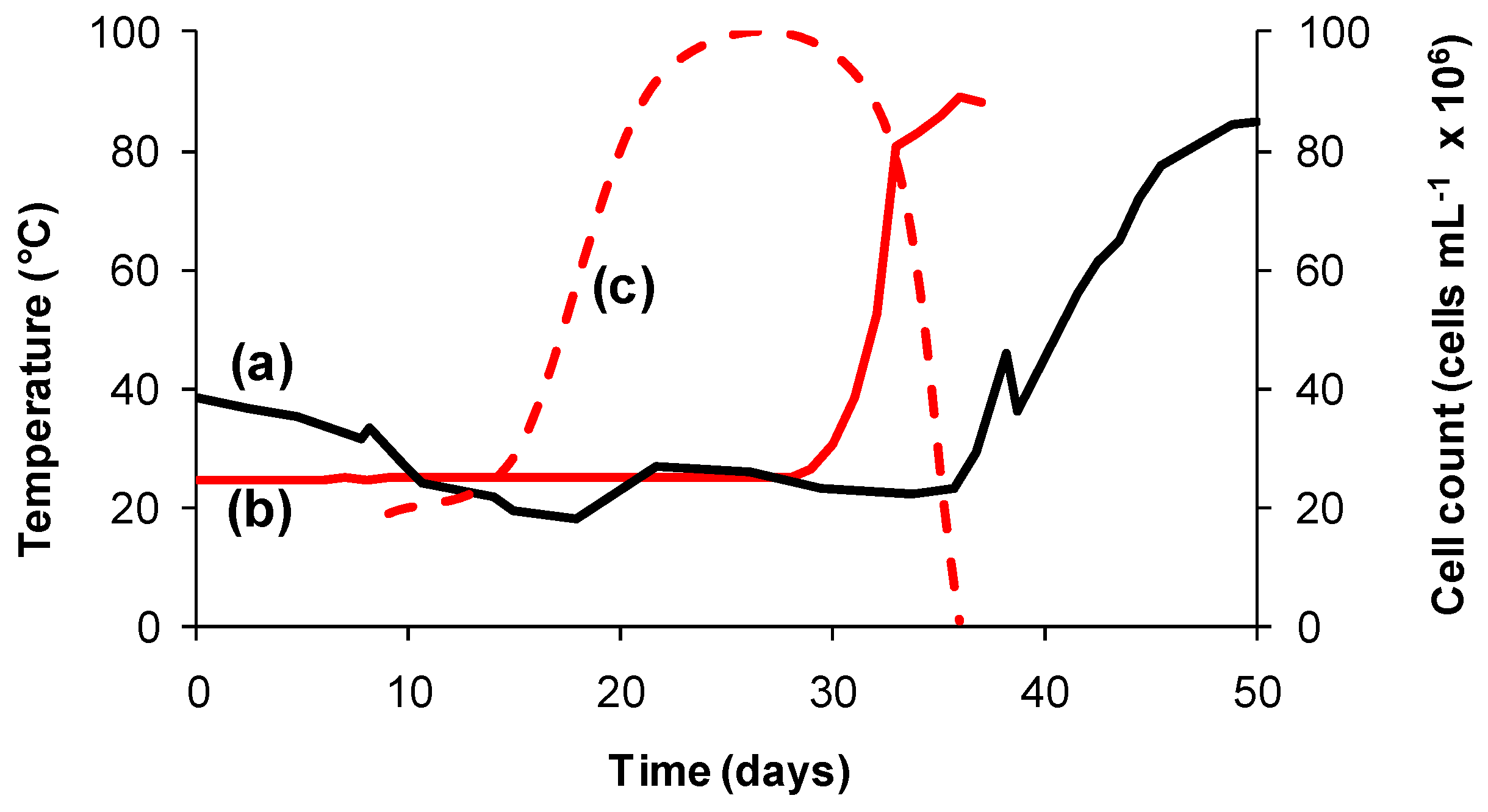
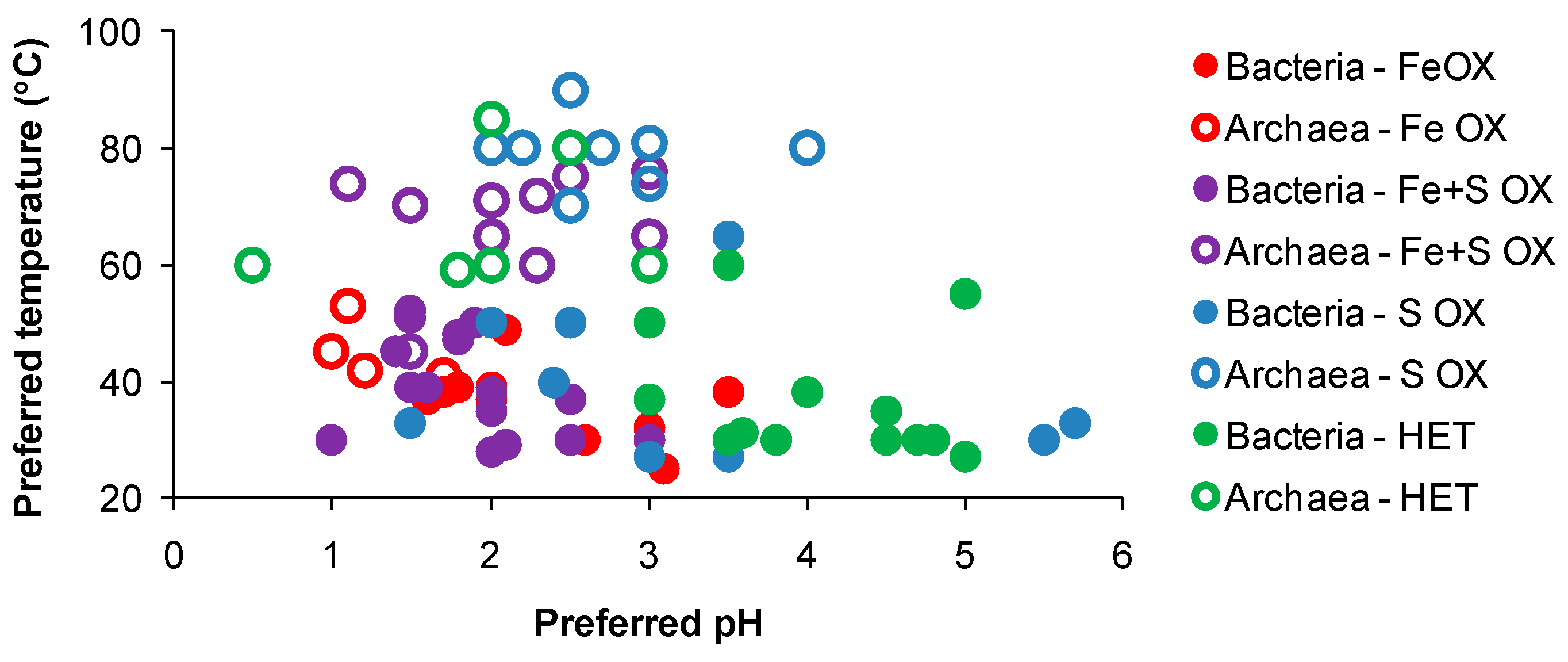
3.1. Temperature
3.2. Acidity
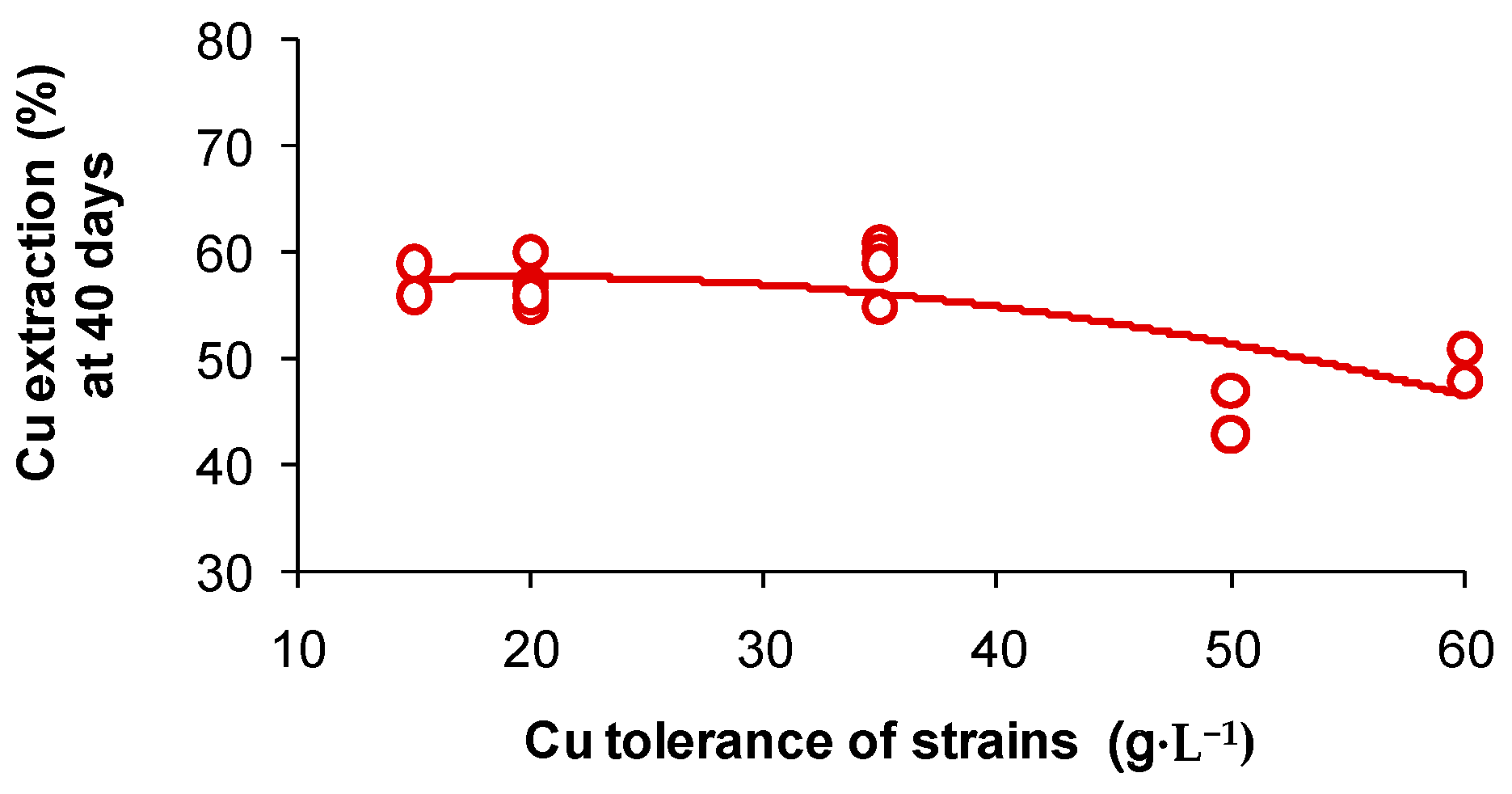
3.3. Cations
3.4. Anions
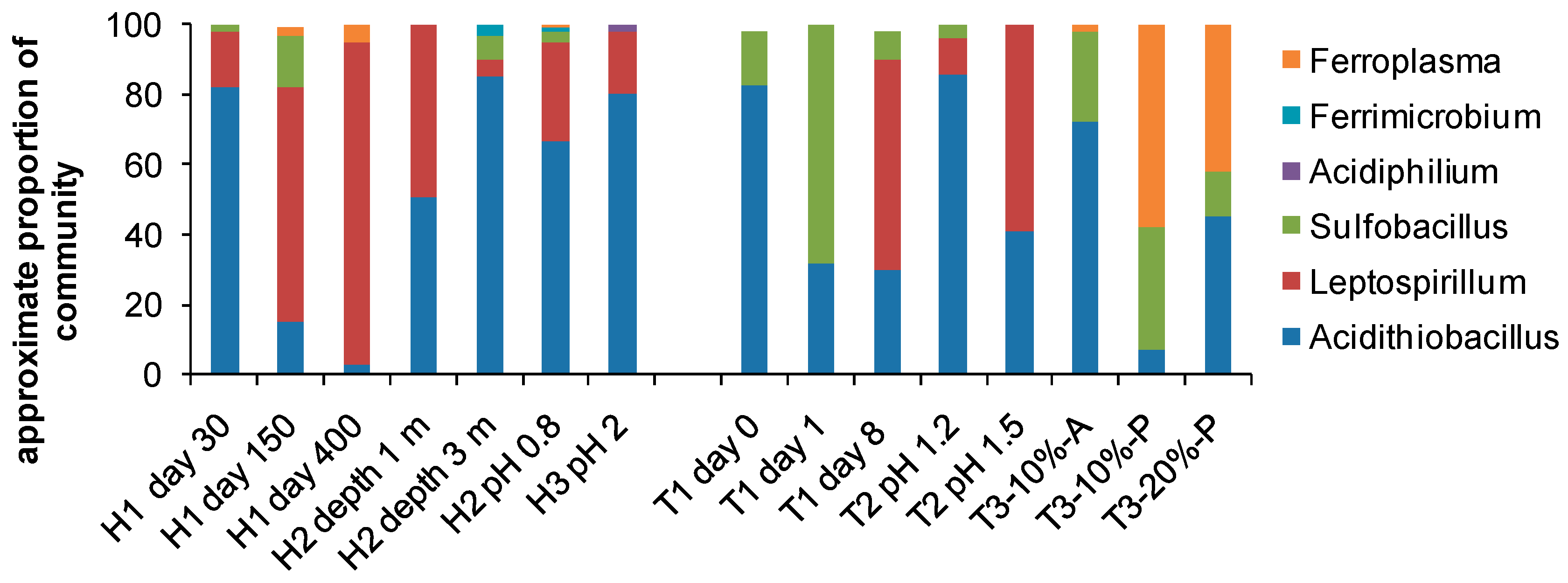
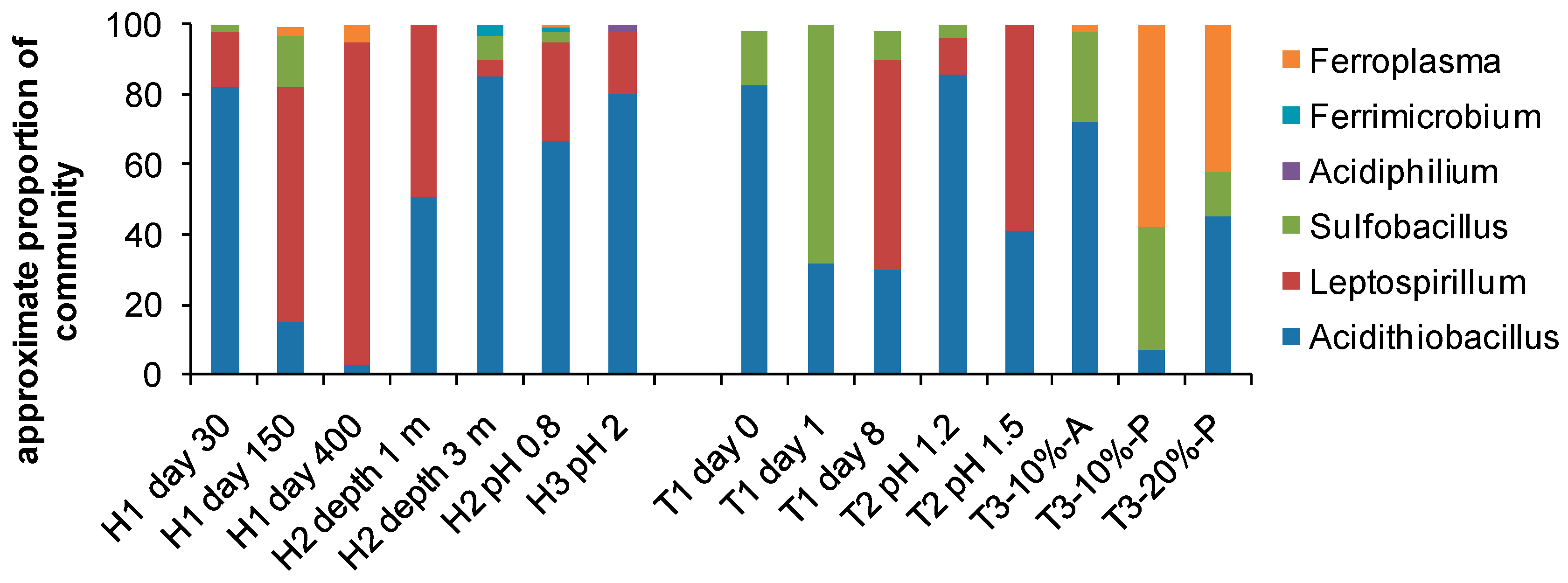
4. Biodiversity in Bioleaching Environments
5. Summary
Acknowledgments
Conflicts of Interest
References
- McDonald, R.G.; Whittington, B.I. Atmospheric leaching of nickel laterites review. Part 1. Sulphuric acid technologies. Hydrometallurgy 2008, 91, 35–55. [Google Scholar] [CrossRef]
- McDonald, R.G.; Whittington, B.I. Atmospheric leaching of nickel laterites review. Part 2. Chloride and bio-technologies. Hydrometallurgy 2008, 91, 56–69. [Google Scholar] [CrossRef]
- McDonald, R.G.; Muir, D.M. Pressure oxidation leaching of chalcopyrite. Part I. Comparison of high and low temperature reaction kinetics and products. Hydrometallurgy 2007, 86, 191–205. [Google Scholar] [CrossRef]
- McDonald, R.G.; Muir, D.M. Pressure oxidation leaching of chalcopyrite. Part II. Comparison of medium temperature kinetics and products and effect of chloride ion. Hydrometallurgy 2007, 86, 206–220. [Google Scholar] [CrossRef]
- Holmes, D.S.; Debus, K.A. Biological opportunities for metal recovery. In Proceedings of the International Symposium on Biotechnology for Energy, Faisalabad, Pakistan, 16–21 December 1989; Malik, K.A., Naqvi, S.H.M., Aleem, N.I.H., Eds.; Nuclear Institute for Agriculture and Biology: Faisalabad, Pakistan, 1991; pp. 341–358. [Google Scholar]
- Poulter, S.; Canterford, J.; Lunt, D. Biotechnology for sulphide minerals. A comparison with other leaching processes. In Proceedings of the Biomine 99 and Waste Management in Metallurgical Operations Conference, Perth, Australia, 23–24 August 1999; Australian Mineral Foundation: Glenside, Australia, 1999; pp. 1–14. [Google Scholar]
- Domic, E. A review of the development and current status of copper bioleaching operations in Chile: 25 years of successful commercial implementation. In Biomining; Rawlings, D.E., Johnson, D.B., Eds.; Springer: Berlin, Germany, 2007; pp. 81–95. [Google Scholar]
- Watling, H.R. The bioleaching of sulphide minerals with emphasis on copper sulphides—A review. Hydrometallurgy 2006, 84, 81–108. [Google Scholar] [CrossRef]
- Van Aswegen, P.C.; Haines, A.K.; Marais, H.J. Design and operation of a commercial bacterial oxidation plant at Fairview. In Proceedings of the Randol Gold Conference, Perth, Australia, 14–17 March 1995; Randol International: Golden, CO, USA, 1988; pp. 144–147. [Google Scholar]
- Van Niekerk, J. Factors affecting the selection of BIOX® as the preferred technology for the treatment of a refractory gold concentrate. Adv. Mater. Res. 2015, 1130, 191–196. [Google Scholar] [CrossRef]
- Hiskey, J.B. In situ leaching of copper: What’s next? In Proceedings of the Hydrometallurgy ’94, Cambridge, UK, 11–15 July 1994; Chapman & Hall: London, UK, 1994; pp. 43–67. [Google Scholar]
- Pliny the Elder: the Natural History. Volume 37. Available online: http://penelope.uchicago.edu/Thayer/E/Roman/Texts/Pliny_the_Elder/home.html (accessed on 20 May 2016).
- Walsh, J. Galen visits the Dead Sea and the copper mines of Cyprus. Bull. Geogr. Soc. Phila. 1927, 25, 93–110. [Google Scholar]
- Lung, T.N. The history of copper cementation on iron—World’s first hydrometallurgical process from medieval China. Hydrometallurgy 1986, 17, 113–129. [Google Scholar] [CrossRef]
- Salkield, L.U. A Technical History of the Rio Tinto Mines: Some Notes on Exploitation from Pre-Phoenician Times to the 1950s; Institution of Mining and Metallurgy: London, UK, 1987. [Google Scholar]
- Agricola, G. De re Metallica; Froben: Basel, Switzerland, 1556; Available online: www.gutenberg.org/ebooks/38015 (accessed on 20 May 2016).
- Lipman, J.G.; Waksman, S.A.; Joffe, J.S. The oxidation of sulfur by soil microorganisms. Soil Sci. 1921, 12, 475–489. [Google Scholar] [CrossRef]
- Waksman, S.A.; Joffe, J.S. Microorganisms concerned in the oxidation of sulfur in the soil. II. Thiobacillus thiooxidans, a new sulfur-oxidizing organism isolated from the soil. J. Bacteriol. 1922, 7, 239–256. [Google Scholar] [PubMed]
- Rudolfs, W. Oxidation of iron pyrites by sulfur-oxidizing organisms and their use for making mineral phosphates available. Soil Sci. 1922, 14, 135–147. [Google Scholar] [CrossRef]
- Rudolfs, W.; Helbronner, A. Oxidation of zinc sulfide by microorganisms. Soil Sci. 1922, 14, 459–464. [Google Scholar] [CrossRef]
- Falck, R.; Kingma, V.T. The methodological and principles of production of organic acids by biological methods with help of aphanomyces. Ber. Deutsch. Chem. Ges. 1924, 57, 915–920. [Google Scholar] [CrossRef]
- Bernhauer, K. The problem of the formation of acids by Aspergillus niger. (Introductory announcement). Biochem. Z. 1924, 153, 517–521. [Google Scholar]
- Colmer, A.R.; Hinkle, M.E. The role of microorganisms in acid mine drainage. Science 1947, 106, 253–256. [Google Scholar] [CrossRef] [PubMed]
- Colmer, A.R.; Temple, K.L.; Hinkle, H.E. An iron-oxidizing bacterium from the acid mine drainage of some bituminous coal mines. J. Bacteriol. 1950, 59, 317–328. [Google Scholar] [PubMed]
- Temple, K.L.; Colmer, A.R. The autotrophic oxidation of iron by a new bacterium: Thiobacillus ferrooxidans. J. Bacteriol. 1951, 62, 605–611. [Google Scholar] [PubMed]
- Oldfield, J.D.; Shaw, D.J.B. VI Vernadskii and the development of biogeochemical understandings of the biosphere, c. 1880s–1968. Brit. J. Hist. Sci. 2013, 46, 287–310. [Google Scholar] [CrossRef]
- Krasilnikov, N.A. The role of micro-organisms in rock weathering. I. Microflora of rock stratum surfaces. Mikrobiologiya 1949, 18, 318–323. [Google Scholar]
- Krasilnikov, N.A. The role of micro-organisms in rock weathering. II. The spread of micro-organisms from foci on rock surfaces. Mikrobiologiya 1949, 18, 492–497. [Google Scholar]
- Leathen, W.W.; Braley, S.S., Sr.; McIntyre, L.D. The role of bacteria in the formation of acid from sulfuritic constituents associated with bituminous coal: II. Ferrous iron oxidizing bacteria. Appl. Microbiol. 1953, 1, 65–68. [Google Scholar] [PubMed]
- Bryner, L.C.; Jameson, A.K. Microorganisms in leaching sulfide minerals. Appl. Microbiol. 1958, 6, 281–287. [Google Scholar] [PubMed]
- Rawlings, D.E. Characteristics and adaptability of iron- and sulfur-oxidizing microorganisms used for the recovery of metals from minerals and their concentrates. Microb. Cell Factories 2005, 4. [Google Scholar] [CrossRef] [PubMed]
- Norris, P.R. Acidophile diversity in mineral sulfide oxidation. In Biomining; Rawlings, D.E., Johnson, D.B., Eds.; Springer: Berlin, Germany, 2007; pp. 199–216. [Google Scholar]
- Johnson, D.B. Biodiversity and interactions of acidophiles: Key to understanding and optimizing microbial processing of ores and concentrates. Trans. Nonferr. Met. Soc. China 2008, 18, 1367–1373. [Google Scholar] [CrossRef]
- Orell, A.; Navarro, C.A.; Aranciba, R.; Mobarec, J.C.; Jerez, C.A. Life in blue: Copper resistance mechanisms of Bacteria and Archaea used in industrial biomining of minerals. Biotechnol. Adv. 2010, 28, 839–848. [Google Scholar] [CrossRef] [PubMed]
- Watling, H.R.; Watkin, E.L.J.; Ralph, D.E. The resilience and versatility of acidophiles that contribute to the bioassisted extraction of metals from mineral sulphides. Environ. Technol. 2010, 31, 915–933. [Google Scholar] [CrossRef] [PubMed]
- Johnson, D.B. Extremophiles: Acidic Environments. In Encyclopedia of Microbiology; Schaechter, M., Ed.; Elsevier: Oxford, UK, 2009; pp. 107–126. [Google Scholar]
- Johnson, D.B.; Hallberg, K.B. Carbon, iron and sulfur metabolism in acidophilic micro-organisms. Adv. Microb. Physiol. 2009, 54, 201–255. [Google Scholar]
- Norris, P.R.; Burton, N.P.; Foulis, N.A.M. Acidophiles in bioreactor mineral processing. Extremophiles 2000, 4, 71–76. [Google Scholar] [CrossRef] [PubMed]
- Baker-Austin, C.; Dopson, M. Life in acid: pH homeostasis in acidophiles. Trends Microbiol. 2007, 15, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Auernik, K.S.; Cooper, C.R.; Kelly, R.M. Life in hot acid: Pathway analyses in extremely thermoacidophilic archaea. Curr. Opin. Biotechnol. 2008, 19, 445–453. [Google Scholar] [CrossRef] [PubMed]
- Kato, S.; Itoh, T.; Yamagishi, A. Archaeal diversity in a terrestrial acidic spring field revealed by PCR primer targeting archaeal 16S rRNA genes. FEMS Microbiol. Lett. 2011, 319, 34–43. [Google Scholar] [CrossRef] [PubMed]
- Ehrlich, H.L.; Newman, D.K.; Kappler, A. Geomicrobiology, 6th ed.; CRC Press: Boca Raton, FA, USA, 2015; p. 635, (and references therein). [Google Scholar]
- Alexander, B.; Leach, S.; Ingledew, W.J. The relationship between chemiosmotic parameters and sensitivity to anions and organic acids in the acidophile Thiobacillus ferrooxidans. J. Gen. Microbiol. 1987, 133, 1171–1179. [Google Scholar] [CrossRef]
- Wheaton, G.; Counts, J.; Mukherjee, A.; Kruh, J.; Kelly, R. The confluence of heavy metal biooxidation and heavy metal resistance: Implications for bioleaching by extreme thermoacidophiles. Minerals 2015, 5, 397–451. [Google Scholar] [CrossRef]
- Rawlings, D.E. Relevance of cell physiology and genetic adaptability of biomining microorganisms to industrial processes. In Biomining; Rawlings, D.E., Johnson, D.B., Eds.; Springer-Verlag: Heidelberg, Germany, 2007; pp. 177–198. [Google Scholar]
- Dixon, D.G. Analysis of heat conservation during copper sulphide heap leaching. Hydrometallurgy 2000, 58, 27–41. [Google Scholar] [CrossRef]
- Hunter, C. BioHeap™ leaching of a primary nickel-copper sulphide ore. In Proceedings of the Nickel-Cobalt-8 Technical Proceedings, Perth, Australia, 20–22 May 2002; ALTA Metallurgical Services: Melbourne, Australia, 2002. [Google Scholar]
- Readett, D.; Sylwestrzak, L.; Franzmann, P.D.; Plumb, J.J.; Robertson, W.R.; Gibson, J.A.E.; Watling, H. The life cycle of a chalcocite heap bioleach system. In Hydrometallurgy 2003; Young, C.A., Alfantazi, A.M., Anderson, C.G., Dreisinger, D.B., Harris, B., James, A., Eds.; TMS: Warrendale, PA, USA, 2003; pp. 365–374. [Google Scholar]
- Plumb, J.J.; Hawkes, R.B.; Franzmann, P.D. The microbiology of moderately thermophilic and transiently thermophilic ore heaps. In Biomining; Rawlings, D.E., Johnson, D.B., Eds.; Springer-Verlag: Berlin, Germany, 2007; pp. 217–235. [Google Scholar]
- Shiers, D.W.; Maree, M.D.; Collinson, D.M.; Watling, H.R.; Hosken, T.; Ingram, G.D. Use of a dynamically controlled column to assess the impact of temperature on copper extraction and microbial activity during copper sulfide bioleaching. In Proceeding of the Goldschmidt 2015 25th Anniversary, Praguz, Czech Republic, 16–21 August 2015.
- Franzmann, P.D.; Haddad, C.M.; Hawkes, R.B.; Robertson, W.J.; Plumb, J.J. Effects of temperature on the rates of iron and sulfur oxidation by selected bioleaching Bacteria and Archaea: Application of the Ratkowsky equation. Min. Eng. 2005, 18, 1304–1314. [Google Scholar] [CrossRef]
- Watling, H.R.; Perrot, F.A.; Shiers, D.W. Comparison of selected characteristics of Sulfobacillus species and review of their occurrence in acidic and bioleaching environments. Hydrometallurgy 2008, 93, 57–65. [Google Scholar] [CrossRef]
- Brierley, J.A. Response of microbial systems to thermal stress in biooxidation-heap pretreatment of refractory gold ores. Hydrometallurgy 2003, 71, 13–19. [Google Scholar] [CrossRef]
- Logan, T.C.; Seal, T.; Brierley, J.A. Whole ore heap biooxidation of sulfidic gold-bearing ores. In Biomining; Rawlings, D.E., Johnson, D.B., Eds.; Springer: Berlin, Germany, 2007; pp. 113–138. [Google Scholar]
- Soto, P.; Acosta, M.; Tapia, P.; Contador, Y.; Velásquez, A.; Espoz, C.; Pinilla, C.; Galleguillos, P.; Demergasso, C. From mesophilic to moderate thermophilic populations in an industrial heap bioleaching process. Adv. Mater. Res. 2013, 825, 376–379. [Google Scholar] [CrossRef]
- Plumb, J.J.; Muddle, R.; Franzmann, P.D. Effect of pH on rates of iron and sulfur oxidation by bioleaching microorganisms. Min. Eng. 2008, 21, 76–82. [Google Scholar] [CrossRef]
- Yahya, A.; Johnson, D.B. Bioleaching of pyrite at low pH and low redox potentials by novel mesophilic gram-positive bacteria. Hydrometallurgy 2002, 63, 181–188. [Google Scholar] [CrossRef]
- Tupikina, O.V.; Ngoma, I.E.; Minnaar, S.; Harrison, S.T.L. Some aspects of the effect of pH and acid stress in heap bioleaching. Min. Eng. 2011, 24, 1209–1214. [Google Scholar] [CrossRef]
- Zepeda, V.J.; Cautivo, D.; Galleguillos, P.A.; Salazar, C.N.; Velasquez, A.; Pinilla, C.; Demergasso, C.S. Effect of increased acid concentration on the microbial population inhabiting an industrial heap bioleaching plant. Adv. Mater. Res. 2013, 825, 348–351. [Google Scholar] [CrossRef]
- Hawkes, R.B.; Franzmann, P.D.; O’Hara, G.; Plumb, J.J. Ferroplasma cupricumulans sp. nov. a novel moderately thermophilic acidophilic archaeon isolated from an industrial-scale chalcocite bioleach heap. Extremophiles 2008, 10, 525–530. [Google Scholar] [CrossRef] [PubMed]
- Golyshina, O.V.; Yakimov, M.M.; Lunsdorf, H.; Ferrer, M.; Nimtz, M.; Timmis, K.N.; Wray, V.; Tindall, B.J.; Golyshin, P.N. Acidiplasma aeolicum gen. nov., sp. nov., a euryarchaeon of the family Ferroplasmaceae isolated from a hydrothermal pool and transfer of Ferroplasma cupricumulans to Acidiplasma cupricumulans comb. nov. Int. J. Syst. Evol. Microbiol. 2009, 59, 2815–2823. [Google Scholar] [CrossRef] [PubMed]
- Schleper, C.; Pühler, G.; Klenk, H.-P.; Zillig, W. Picrophilus oshimae and Picrophilus torridus fam. nov., gen. nov., sp. nov., two species of hyperacidophilic, thermophilic, heterotrophic, aerobic archaea. Int. J. Syst. Bacteriol. 1996, 46, 814–816. [Google Scholar] [CrossRef]
- Plumb, J.J.; Haddad, C.M.; Gibson, J.A.E.; Franzmann, P.D. Acidianus sulfidivorans sp. nov., an extremely acidophilic, thermophilic archaeon isolated from a solfatara on Lihir Island, Papua New Guinea, and emendation of the genus description. Int. J. Syst. Evol. Microbiol. 2007, 57, 1418–1423. [Google Scholar] [CrossRef] [PubMed]
- Watling, H.R. Adaptability of biomining organisms in hydrometallurgical processes. In Biohydrometallurgical Processes: A Practical Approach; Santos Sobral, L.G., Monteiro de Oliveira, D., Gomes de Souza, C.E., Eds.; CETEM/MCTI: Rio de Janeiro, Brazil, 2011; pp. 39–70. [Google Scholar]
- Watkin, E.L.J.; Keeling, S.E.; Perrot, F.A.; Shiers, D.W.; Palmer, M.-L.; Watling, H.R. Metals tolerance in moderately thermophilic isolates from a spent copper sulfide heap, closely related to Acidithiobacillus caldus, Acidimicrobium ferrooxidans and Sulfobacillus thermosulfidooxidans. J. Ind. Microbiol. Biotechnol. 2009, 36, 461–465. [Google Scholar] [CrossRef] [PubMed]
- Battaglia, F.; Morin, D.; Ollivier, P. Dissolution of cobaltiferous pyrite by Thiobacillus ferrooxidans and Thiobacillus thiooxidans: Factors influencing bacterial leaching efficiency. J. Biotechnol. 1994, 32, 11–16. [Google Scholar] [CrossRef]
- D’Hughes, P.; Cezac, P.; Cabral, T.; Battaglia, F.; Truong-Meyer, X.M.; Morin, D. Bioleaching of a cobaltiferous pyrite: A continuous laboratory-scale study at high solids concentration. Min. Eng. 1997, 10, 507–527. [Google Scholar]
- Heinzle, T.; Miller, D.; Nagel, V. Results of an integrated pilot plant operation using the BioNIC® process to produce nickel metal. In Proceedings of the Biomine 99 and Water Management in Metallurgical Operations 99, Perth, Australia, 24–25 August 1999; AusIMM: Melbourne, Australia, 1999; pp. 16–25. [Google Scholar]
- Du Plessis, C.A.; Batty, J.D.; Dew, D.W. Commercial applications of thermophile bioleaching. In Biomining; Rawlings, D.E., Johnson, D.B., Eds.; Springer-Verlag: Berlin, Germany, 2007; pp. 57–80. [Google Scholar]
- Rea, S.M.; McSweeney, N.J.; Degens, B.P.; Morris, C.; Siebert, H.M.; Kaksonen, A.H. Salt-tolerant microorganisms potentially useful for bioleaching operations where fresh water is scarce. Min. Eng. 2015, 75, 126–132. [Google Scholar] [CrossRef]
- Harahuc, L.; Lizama, H.M.; Suzuki, I. Selective inhibition of the oxidation of ferrous iron or sulfur in Thiobacillus ferrooxidans. Appl. Environ. Microbiol. 2000, 66, 1031–1037. [Google Scholar] [CrossRef] [PubMed]
- Zammit, C.M.; Mangold, S.; rao Jonna, V.; Mutch, L.A.; Watling, H.R.; Dopson, M.; Watkin, E.L.J. Bioleaching in brackish waters—Effect of chloride ions on the acidophilic population and proteomes of model species. Appl. Microbiol. Biotechnol. 2012, 93, 319–329. [Google Scholar] [CrossRef] [PubMed]
- Johnson, D.B.; Grail, B.M.; Bonnefoy, V. New insights into salt-tolerance in acidophilic iron-oxidising bacteria. Adv. Mater. Res. 2015, 1130, 3–6. [Google Scholar] [CrossRef]
- Watling, H.R.; Shiers, D.W.; Zhang, G.J. Microbial behaviour under conditions relevant to heap leaching: Studies using the sulfur-oxidising, moderate thermophile Acidithiobacillus caldus. Hydrometallurgy 2012, 127–128, 104–111. [Google Scholar] [CrossRef]
- Cardenas, J.P.; Ortiz, R.; Norris, P.R.; Watkin, E.; Holmes, D.S. Reclassification of ‘Thiobacillus prosperus’ Huber and Stetter 1989 as Acidihalobacter prosperus gen. nov., sp. nov., a member of the family Ectothiorhodospiraceae. Int. J. Syst. Evol. Microbiol. 2015, 65, 3641–3644. [Google Scholar] [CrossRef] [PubMed]
- Kelly, D.P.; Wood, A.P. Reclassification of some species of Thiobacillus to the newly designated genera Acidithiobacillus gen. nov., Halothiobacillus gen. nov., and Thermithiobacillus gen. nov. Int. J. Syst. Evol. Microbiol. 2000, 50, 511–516. [Google Scholar] [CrossRef] [PubMed]
- Brierley, J.A.; Kuhn, M.C. Fluoride toxicity in a chalcocite bioleach heap process. Hydrometallurgy 2010, 104, 410–413. [Google Scholar] [CrossRef]
- Murr, L.E.; Brierley, J.A. The use of large-scale test facilities in commercial leaching operations. In Metallurgical Applications of Bacterial Leaching and Related Microbiological Phenomena; Murr, L., Torma, A., Brierley, J., Eds.; Academic Press: New York, NY, USA, 1978; pp. 491–520. [Google Scholar]
- Soljanto, P.; Rehtijarvi, P.; Tuovinen, O.H. Ferrous iron oxidation by Thiobacillus ferrooxidans: Inhibition by finely ground particles. Geomicrobiol. J. 1980, 2, 1–12. [Google Scholar] [CrossRef]
- Suzuki, I.; Lee, D.; McKay, B.; Harahuc, L.; Oh, J.K. Effect of various anions, pH, and osmotic pressure on oxidation of elemental sulfur by Thiobacillus thiooxidans. Appl. Environ. Microbiol. 1999, 65, 5163–5168. [Google Scholar] [PubMed]
- Peng, Z.; Yu, R.; Qiu, G.; Qin, W.; Gu, G.; Wang, Q.; Li, Q.; Liu, X. Really active form of fluorine toxicity affecting Acidithiobacillus ferrooxidans activity in bioleaching uranium. Trans. Nonferr. Met. Soc. China 2013, 23, 812–817. [Google Scholar] [CrossRef]
- Ma, L.; Li, Q.; Xiao, Y.; Wang, Q.; Yin, H.; Liang, Y.; Qiu, G.; Liu, X. Comparative study of fluoride tolerance of five typical bioleaching microorganisms. Adv. Mater. Res. 2013, 825, 214–218. [Google Scholar] [CrossRef]
- Wang, Q.; Liu, Y.; Hu, K.; Hu, E. Experimental study on acclimatization and culture of acid-tolerant and fluoride-tolerant bacteria. Min. Res. Dev. 2005, 25, 35–36. [Google Scholar]
- Sundkvist, J.-E.; Sandström, Å.; Gunneriusson, L.; Lindstrom, E.B. Fluorine toxicity in bioleaching systems. In Proceedings of the 16th International Biohydrometallurgy Symposium, Cape Town, South Africa, 25–29 September 2005; Harrison, S.T.L., Rawlings, D.E., Petersen, J., Eds.; IBS: Cape Town, South Africa, 2005; pp. 19–28. [Google Scholar]
- Corbillon, M.S.; Olazabal, M.A.; Madariaga, J.M. Potentiometric study of aluminium fluoride complexation equilibria and definition of the thermodynamic model. J. Solut. Chem. 2008, 37, 567–579. [Google Scholar] [CrossRef]
- Radic, N.; Bralic, M. Aluminium fluoride complexation and its ecological importance in the aquatic environment. Sci. Tot. Environ. 1995, 172, 237–243. [Google Scholar] [CrossRef]
- Soli, A.L.; Byrne, R.H. The hydrolysis and fluoride complexation behaviour of Fe(III) at 25 °C and 0.68 molal ionic strength. J. Solut. Chem. 1996, 25, 773–785. [Google Scholar] [CrossRef]
- Sicupira, L.; Veloso, T.; Reis, F.; Leao, V. Assessing metal recovery from low-grade copper ores containing fluoride. Hydrometallurgy 2011, 109, 202–210. [Google Scholar] [CrossRef]
- Razzell, W.E.; Trussell, P.C. Isolation and properties of an iron-oxidizing Thiobacillus. J. Bacteriol. 1963, 85, 595–603. [Google Scholar] [PubMed]
- Harahuc, L.; Lizama, H.; Suzuki, I. Effect of anions on selective solubilisation of zinc and copper in bacterial leaching of sulfide ores. Biotechnol. Bioeng. 2000, 69, 196–203. [Google Scholar] [CrossRef]
- Shiers, D.W.; Ralph, D.E.; Watling, H.R. The effects of nitrate on substrate utilisation by some iron(II)- and sulfur-oxidising Bacteria and Archaea. Hydrometallurgy 2014, 150, 259–268. [Google Scholar] [CrossRef]
- Romero, J.; Yañez, C.; Vásquez, M.; Moore, E.R.B.; Espejo, R.T. Characterization and identification of an iron-oxidizing, Leptospirillum-like bacterium present in the high-sulfate leaching solution of a commercial bioleaching plant. Res. Microbiol. 2003, 154, 353–359. [Google Scholar] [CrossRef]
- Ruan, R.; Liu, X.; Zou, G.; Chen, J.; Wen, J.; Wang, D. Industrial practice of a distinct bioleaching system operated at low pH, high ferric concentration, elevated temperature and low redox potential for secondary copper sulfide. Hydrometallurgy 2011, 108, 130–135. [Google Scholar] [CrossRef]
- Shiers, D.W.; Blight, K.R.; Ralph, D.E. Sodium sulphate and sodium chloride effects on batch culture of iron oxidising bacteria. Hydrometallurgy 2005, 80, 75–82. [Google Scholar] [CrossRef]
- Watling, H.R.; Collinson, D.M.; Li, J.; Mutch, L.A.; Perrot, F.A.; Rea, S.M.; Reith, F.; Watkin, E.L.J. Bioleaching of a low-grade copper ore linking leach chemistry and microbiology. Min. Eng. 2014, 56, 35–44. [Google Scholar] [CrossRef]
- Cautivo, D.; Soto, P.; Zepeda, V.J.; Galleguillos, P.A.; Velasquez, A.; Pinilla, C.; Demergasso, C.S. Estimation of ionic load effect on the oxidizing activity of the microbial population in the heap bioleaching process at Escondida Mine. Adv. Mater. Res. 2013, 825, 219–222. [Google Scholar] [CrossRef]
- He, Z.; Xiao, S.; Xie, X.; Hu, Y. Microbial diversity in acid mineral bioleaching systems of Dongxiang copper mine and Yinshan lead-zinc mine. Extremophiles 2008, 12, 225–234. [Google Scholar] [CrossRef] [PubMed]
- Hao, C.; Wang, L.; Dong, H.; Zhang, H. Succession of acidophilic bacterial community during bio-oxidation of refractory gold-containing sulfides. Geomicrobiol. J. 2010, 27, 683–691. [Google Scholar] [CrossRef]
- Remonsellez, F.; Galleguillos, F.; Moreno-Paz, M.; Parro, V.; Acosta, M.; Demeragasso, C. Dynamic of active microorganisms inhabiting a bioleaching industrial heap of low-grade copper sulfide ore monitored by real-time PCR and olignucleotide prokaryotic acidophile microarray. Microbiol. Biotechnol. 2009, 2, 613–624. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Chen, B.; Wen, J.; Ruan, R. Leptospirillum forms a minor portion of the population in Zijinshan commercial non-aeration copper bioleaching heap identified by 16S rRNA clone libraries and real-time PCR. Hydrometallurgy 2010, 104, 399–403. [Google Scholar]
- Liu, X.; Wu, B.; Chen, B.; Wen, J.; Ruan, R.; Yao, G.; Wang, D. Bioleaching of chalcocite started at different pH: Response of the microbial community to environmental stress and leaching kinetics. Hydrometallurgy 2010, 103, 1–6. [Google Scholar]
- Wang, Y.; Su, L.; Zeng, W.; Wan, L.; Chen, Z.; Zhang, L.; Qiu, G.; Chen, X.; Zhou, H. Effect of pulp density on planktonic and attached community dynamics during bioleaching of chalcopyrite by a moderately thermophilic microbial culture under uncontrolled conditions. Min. Eng. 2014, 61, 66–72. [Google Scholar] [CrossRef]
- Demergasso, C.; Galleguillos, F.; Soto, P.; Serón, M.; Iturriaga, V. Microbial succession during heap bioleaching cycle of low grade copper sulfides. Does this knowledge mean a real input for industrial process design and control? Hydrometallurgy 2010, 104, 382–390. [Google Scholar] [CrossRef]
- Okibe, N.; Gericke, M.; Hallberg, K.B.; Johnson, D.B. Enumeration and characterization of acidophilic microorganisms isolated from a pilot plant stirred-tank bioleaching operation. Appl. Environ. Microbiol. 2003, 69, 1936–1943. [Google Scholar] [CrossRef] [PubMed]
- Kawabe, Y.; Inoue, C.; Suto, K.; Chida, T. Inhibitory effect of high concentrations of ferric ions on the activity of Acidithiobacillus ferrooxidans. J. Biosci. Bioeng. 2003, 96, 375–379. [Google Scholar] [CrossRef]
- Cameron, R.A.; Yeung, C.W.; Greer, C.W.; Gould, W.D.; Mortazavi, S.; Bedard, P.L.; Morin, L.; Lortie, L.; Dinardo, O.; Kennedy, K.J. The bacterial community structure during bioleaching of a low-grade nickel sulphide ore in stirred tank reactors at different combinations of temperature and pH. Hydrometallurgy 2010, 104, 207–215. [Google Scholar] [CrossRef]
- Mutch, L.A.; Watling, H.R.; Watkin, E.L.J. Microbial population dynamics of inoculated low-grade chalcopyrite bioleaching columns. Hydrometallurgy 2010, 104, 391–398. [Google Scholar] [CrossRef]
- Yu, R.; Shi, L.; Gu, G.; Zhou, D.; You, L.; Chen, M.; Qiu, G.; Zeng, W. The shift in microbial community under the adjustment of initial and processing pH during bioleaching of chalcopyrite concentrate by moderate thermophiles. Bioresour. Technol. 2014, 162, 300–307. [Google Scholar] [CrossRef] [PubMed]
- Segerer, A.H.; Burggraf, S.; Fiala, G.; Huber, G.; Huber, R.; Pley, U.; Stetter, K.O. Life in hot springs and hydrothermal vents. Orig. Life Evolut. Biosph. 1993, 23, 77–90. [Google Scholar] [CrossRef]
- Childs, A.M.; Mountain, B.W.; O’Toole, R.; Stott, M.B. Relating microbial community and physicochemical parameters of a hot spring: Champagne Pool, Wai-o-tapu, New Zealand. Geomicrobiol. J. 2008, 25, 441–453. [Google Scholar] [CrossRef]
- Stott, M.B.; Sutton, D.C.; Watling, H.R.; Franzmann, P.D. Comparative leaching of chalcopyrite by selected acidophilic Bacteria and Archaea. Geomicrobiol. J. 2003, 20, 215–230. [Google Scholar] [CrossRef]
- Zhang, L.; Peng, J.; Wei, M.; Ding, J.; Zhou, H. Bioleaching of chalcopyrite with Acidianus manzaensis YN25 under contact and non-contact conditions. Trans. Nonferr. Met. Soc. China 2010, 20, 1981–1986. [Google Scholar] [CrossRef]
- Zhu, W.; Xia, J.; Yang, Y.; Nie, Z.; Peng, A.; Liu, H.; Qiu, G. Thermophilic archaeal community succession and function change associated with the leaching rate in bioleaching of chalcopyrite. Bioresour. Technol. 2013, 133, 405–413. [Google Scholar] [CrossRef] [PubMed]
- Shiers, D.W.; Ralph, D.E.; Bryan, C.G.; Watling, H.R. Substrate utilisation by Acidianus brierleyi, Metallospaera hakonensis and Sulfolobus metallicus in mixed ferrous ion and tetrathionate growth media. Min. Eng. 2013, 48, 86–93. [Google Scholar] [CrossRef]
- Demergasso, C.; Galleguillos, P.; Escudero, L.; Zepeda, V.; Castillo, D.; Casamayor, E. Molecular characterization of microbial populations in a low grade copper ore bioleaching test heap. Hydrometallurgy 2005, 80, 241–253. [Google Scholar] [CrossRef]
- Fuchs, T.; Huber, H.; Teiner, K.; Burggraf, S.; Stetter, K.O. Metallosphaera prunae, sp. nov., a novel metal-mobilizing, thermoacidophilic Archaeum isolated from a uranium mine in Germany. Syst. Appl. Microbiol. 1995, 18, 560–566. [Google Scholar] [CrossRef]
- Xiao, S.; Xie, X.; Liu, J.; He, Z.; Hu, Y. Composition and structures of archaeal communities in acid mineral bioleaching system of Dongxiang copper mine and Yinshan lead-Zinc Mine, China. Curr. Microbiol. 2008, 57, 239–244. [Google Scholar] [CrossRef] [PubMed]
- Xie, X.; Xiao, S.; He, Z.; Liu, J.; Qiu, G. Microbial populations in acid mineral bioleaching systems of Tong Shankou copper mine, China. J. Appl. Microbiol. 2007, 103, 1227–1238. [Google Scholar] [CrossRef] [PubMed]
- Halinen, A.-K.; Beecroft, N.J.; Määttä, K.; Nurmi, P.; Laukkanen, K.; Kaksonen, A.H.; Riekkola-Vanhanen, M.; Puhakka, J.A. Microbial community dynamics during a demonstration-scale bioheap. Hydrometallurgy 2012, 125–126, 34–41. [Google Scholar] [CrossRef]
- Selkov, E.; Overbeek, R.; Kogan, Y.; Chu, L.; Vonstein, V.; Holmes, D.S.; Silver, S.; Haselkorn, R.; Fonstein, M. Functional analysis of gapped microbial genomes: Amino acid metabolism of Thiobacillus ferrooxidans. PNAS 2000, 97, 3509–3514. [Google Scholar] [CrossRef] [PubMed]
- Cárdenas, J.P.; Valdes, J.; Quatrini, R.; Duarte, F.; Holmes, D.S. Lessons from the genomes of extremely acidophilic bacteria and archaea with special emphasis on bioleaching microorganisms. Appl. Microbiol. Biotechnol. 2010, 88, 605–620. [Google Scholar] [CrossRef] [PubMed]
- Justice, N.B.; Norman, A.; Brown, C.T.; Singh, A.; Thomas, B.C.; Banfield, J.F. Comparison of environmental and isolate Sulfobacillus genomes reveals diverse carbon, sulfur, nitrogen, and hydrogen metabolisms. BMC Genom. 2014, 15. [Google Scholar] [CrossRef] [PubMed]
- Song, J.-L.; Jiang, C.-Y.; Liu, S.-J. Insight into the sulfur metabolism by thermoacidophilic archaeon Metallosphaera cuprina with genomic, proteomic and biochemical tools. Adv. Mater. Res. 2015, 1130, 145–148. [Google Scholar] [CrossRef]
- Ullrich, S.R.; Poehlein, A.; Daniel, R.; Tischler, J.S.; Vogel, S.; Schlömann, M.; Mühling, M. Comparative genomics underlines the functional and taxonomic diversity of novel “Ferrovum” related iron oxidizing bacteria. Adv. Mater. Res. 2015, 1130, 15–18. [Google Scholar] [CrossRef]
- Goltsman, D.S.; Denef, V.J.; Singer, S.W.; VerBerkmoes, N.C.; Lefsrud, M.; Mueller, R.S.; Dick, G.J.; Sun, C.L.; Wheeler, K.E.; Zemla, A.; et al. Community genomic and proteomic analyses of chemoautotrophic iron-oxidizing “Leptospirillum rubarum” (Group II) and “Leptospirillum ferrodiazotrophum” (Group III) bacteria in acid mine drainage biofilms. Appl. Environ. Microbiol. 2009, 75, 4599–4615. [Google Scholar] [CrossRef] [PubMed]
- Hemme, C.L.; Deng, Y.; Gentry, T.J.; Fields, M.W.; Wu, L.; Barua, S.; Barry, K.; Tringe, S.G.; Watson, D.B.; He, Z.; et al. Metagenomic insights into evolution of a heavy metal-contaminated groundwater microbial community. ISME J. 2010, 4, 660–672. [Google Scholar] [CrossRef] [PubMed]
- Méndez-García, C.; Peláez, A.I.; Mesa, V.; Sánchez, J.; Golyshina, O.V.; Ferrer, M. Microbial diversity and metabolic networks in acid mine drainage habitats. Front. Microbiol. 2015, 6. [Google Scholar] [CrossRef]
- Chen, L.; Huang, L.; Méndez-Garcia, C.; Kuang, J.; Hua, Z.; Liu, J.; Shu, W. Microbial communities, processes and functions in acid mine drainage systems. Curr. Opin. Biotechnol. 2016, 38, 150–158. [Google Scholar] [CrossRef] [PubMed]
- Banfield, J.F.; Verboekmoes, N.C.; Hettich, R.L.; Thelen, M.P. Proteogenomic approaches for the molecular characterization of natural microbial communities. OMICS 2005, 9, 301–333. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Niu, J.; Liang, Y.; Liu, X.; Yin, H. Metagenome-scale analysis yields insights into the structure and function of microbial communities in a copper bioleaching heap. BMC Genet. 2016, 17. [Google Scholar] [CrossRef] [PubMed]
- Valenzuela, L.; Chi, A.; Beard, S.; Orell, A.; Guiliani, N.; Shabanowitz, J.; Hunt, D.F.; Jerez, C.A. Genomics, metagenomics and proteomics in biomining microorganisms. Biotechnol. Adv. 2006, 24, 197–211. [Google Scholar] [CrossRef] [PubMed]
- Jerez, C.A. Metal extraction and biomining. In The Desk Encyclopedia of Microbiology; Schaechter, M., Ed.; Elsevier: Oxford, UK, 2009; pp. 762–775. [Google Scholar]
- Stoughton, R.B. Applications of DNA microarrays in biology. Ann. Rev. Biochem. 2005, 74, 53–82. [Google Scholar] [CrossRef] [PubMed]
- Seidel, M.; Niessner, R. Automated analytical microarrays: A critical review. Anal. Bioanal. Chem. 2008, 391, 1521–1544. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Deng, Y.; Zhou, Y. Development of functional gene arrays for microbial community analysis. Curr. Opin. Biotechnol. 2012, 23, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Cao, L.; Qiu, G.; Wang, D.; Kellogg, L.; Zhou, J.; Dai, Z.; Liu, X. Development and evaluation of 50-mer oligonucleotide arrays for detecting microbial populations in acid mine drainages and bioleaching systems. J. Microbiol. Methods 2007, 70, 165–178. [Google Scholar] [CrossRef] [PubMed]
- Tu, Q.; Yu, H.; He, Z.; Deng, Y.; Wu, L.; Van Nostrand, J.D.; Zhou, A.; Voordeckers, J.; Lee, Y.-J.; Qin, Y.; et al. GeoChip 4: A functional gene-array-based high-throughput environmental technology for microbial community analysis. Mol. Ecol. Resour. 2014, 14, 914–928. [Google Scholar] [CrossRef] [PubMed]
- Quatrini, R.; Appia-Ayme, C.; Denis, Y.; Ratouchniak, J.; Veloso, F.; Valdes, J.; Lefimil, C.; Silver, S.; Roberto, F.; Orellana, O.; et al. Insights into the iron and sulfur energetic metabolism of Acidithiobacillus ferrooxidans by microarray transcriptome profiling. Hydrometallurgy 2006, 83, 263–272. [Google Scholar] [CrossRef]
- Quatrini, R.; Appia-Ayme, C.; Denis, Y.; Jedlicki, E.; Holmes, D.S.; Bonnefoy, V. Extending the models for iron and sulfur oxidation in the extreme acidophile Acidithiobacillus ferrooxidans. BMC Genom. 2009, 10, 394. [Google Scholar] [CrossRef] [PubMed]
- Valdés, J.; Pedroso, I.; Quatrini, R.; Holmes, D.S. Comparative genome analysis of Acidithiobacillus ferrooxidans, A. thiooxidans and A. caldus: Insights into their metabolism and ecophysiology. Hydrometallurgy 2008, 94, 180–184. [Google Scholar] [CrossRef]
- Auernik, K.S.; Kelly, R.M. Physiological versatility of the extremely thermoacidophilic archaeon Metallosphaera sedula supported by transcriptomic analysis of heterotrophic, autotrophic and mixotrophic growth. Appl. Environ. Microbiol. 2010, 76, 931–935. [Google Scholar] [CrossRef] [PubMed]
- Auernik, K.S.; Maezato, Y.; Blum, P.H.; Kelly, R.M. The genome sequence of the metal-mobilizing, extremely thermoacidophilic archaeon Metallosphaera sedula provides insights into bioleaching-associated metabolism. Appl. Environ. Microbiol. 2008, 74, 682–692. [Google Scholar] [CrossRef] [PubMed]
- Haward, R.E.; Derisi, J.L.; Alfadhli, S.; Kaslow, D.C.; Brown, P.O.; Rathod, P.K. Shotgun DNA microarrays and stage-specific gene expression in Plasmodium falciparum malaria. Mol. Microbiol. 2000, 35, 6–14. [Google Scholar] [CrossRef]
- Parro, V.; Moreno-Paz, M. Gene function analysis in environmental isolates: The nif regulon of the strict iron oxidizing bacterium Leptospirillum ferrooxidans. PNAS 2003, 100, 7883–7888. [Google Scholar] [CrossRef] [PubMed]
- Tyson, G.W.; Lo, I.; Baker, B.J.; Allen, E.E.; Hugenholtz, P.; Banfield, J.F. Genome-directed isolation of the key nitrogen fixer Leptospirillum ferrodiazotrophum sp. nov. from an acidophilic microbial community. Appl. Environ. Microbiol. 2005, 71, 6319–6324. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Paz, M.; Gómez, M.J.; Arcas, A.; Parro, V. Environmental transcriptome analysis reveals physiological differences between biofilm and planktonic modes of life of the iron oxidizing bacteria Leptospirillum spp. in their natural microbial community. BMC Genomics 2010, 11. [Google Scholar] [CrossRef] [PubMed]
- Vera, M.; Krok, B.; Bellenberg, S.; Sand, W.; Poetsch, A. Shotgun proteomics study of early biofilm formation process of Acidithiobacillus ferrooxidans ATCC 23270 on pyrite. Proteomics 2013, 13, 1133–1144. [Google Scholar] [CrossRef] [PubMed]
- Laska, S.; Lottspeich, F.; Kletzin, A. Membrane-bound hydrogenase and sulfur reductase of the hyperthermophilic and acidophilic archaeon Acidianus ambivalens. Microbiology 2003, 149, 2357–2371. [Google Scholar] [CrossRef] [PubMed]
- Golyshina, O.V. Environmental, biogeographic, and biochemical patterns of archaea of the family Ferroplasmaceae. Appl. Environ. Microbiol. 2011, 77, 5071–5078. [Google Scholar] [CrossRef] [PubMed]
- Brodie, E.L.; DeSantis, T.Z.; Joyner, D.C.; Baek, S.M.; Larsen, J.T.; Andersen, G.L.; Hazen, T.C.; Richardson, P.M.; Herman, D.J.; Tokunaga, T.K.; et al. Application of a high-density oligonucleotide microarray approach to study bacterial population dynamics during uranium reduction and reoxidation. Appl. Environ. Microbiol. 2006, 72, 6288–6298. [Google Scholar] [CrossRef] [PubMed]
- Garrido, P.; Gonzalez-Toril, E.; Garcia-Moyano, A.; Moreno-Paz, M.; Amils, R.; Parro, V. An oligonucleotide prokaryotic acidophile microarray: Its validation and its use to monitor seasonal variations in extreme acidic environments with total environmental RNA. Environ. Microbiol. 2008, 10, 836–850. [Google Scholar] [CrossRef] [PubMed]
- Shen, L.; Yin, H.; Hu, Q.; Liu, X.; Qiu, G. Microarray-based characterization of microbial community functional structure and heterogeneity associated with acid mine drainages. Adv. Mater. Res. 2015, 1130, 40–44. [Google Scholar] [CrossRef]
- Acosta, M.; Galleguillos, P.; Ghorbani, Y.; Tapia, P.; Contador, Y.; Velásquez, A.; Espoz, C.; Pinilla, C.; Demergasso, C. Variation in microbial community from predominantly mesophilic to thermotolerant and moderately thermophilic species in an industrial copper heap bioleaching operation. Hydrometallurgy 2014, 150, 281–289. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chronology | Observation—Report |
|---|---|
| 25–220 A.D. | The Roman Gaius Plinius Secundus (Pliny the Elder, 23–79 A.D.) wrote to the effect that chrysocolla is also artificially produced by slowly passing water through the mine during the winter until the month of June; subsequently the water is evaporated in June and July. |
| Notes in old Chinese texts on leaching of rocks and the formation of gall (CuSO4) springs; copper recovery was by cementation on iron. | |
| The scientist Galen described in situ leaching at a mine in Cyprus. | |
| 1086 A.D. | The Gall (CuSO4)—copper process described: copper leaching and recovery using cementation on iron as a commercial process; the copper was used for coinage. |
| 1500s | Copper leached from some ores (in situ) and mines (Spain, Hungary, Germany) and copper recovered by cementation on iron. A wood cut illustration in Agricola’s De re metallica represents the manual collection of copper solutions in wooden ‘vats’ and transfer to evaporative ponds to concentrate the solutions. |
| 1600s | The extraction of copper from mine water (Peru) described by Alvaro Alonso Barba de Garfias (priest and metallurgist); he was granted a patent to recover copper from mine water. |
| 1800s | Large-scale copper heap leaching (Spain): Heaps of low-grade ore were left for 1–3 years to undergo natural decomposition. |
| 1900–1920 | Commercial copper production by leaching and copper cementation (Butte, MT, USA); in place (stope) leaching (Cananea, Sonora, Mexico); recovery of copper from underground mine water (Bisbee, AZ, USA). |
| 1921–1940 | The oxidation of sulfur by soil microorganisms described; At. thiooxidans (previously Thiobacillus) isolated and described. |
| The concept of the ‘biosphere’ in which living organisms shaped the environment (biogeochemistry) expounded by Vladimir Vernadskii (1863–1945). | |
| Biological production of organic acids reported. | |
| Biooxidation of pyrite first reported and the suggested commercial application of biohydrometallurgy for zinc sulfide. | |
| Large scale dump leaching of open pit waste rock with recirculation of leach solutions (Bingham Canyon, UT, USA). | |
| 1941–1951 | Roles of microorganisms in rock-weathering processes recognised. |
| Acidithiobacillus ferrooxidans isolated from acid drainage of a coal mine and its role in oxidising Fe(II) and reduced inorganic sulfur compounds (RISC) in acid mine drainage (AMD) described. |
| Term | Definition |
|---|---|
| Genomics | The branch of molecular biology concerned with the structure, function, evolution and mapping of genomes (the complete set of genes present in an organism). |
| Metagenomics | The study of the collected genetic material (genomes) from a mixed community of organisms. |
| Proteomics | The analysis of the structure, function and interactions of the proteins produced by the genes of a particular cell, tissue or organism; a proteome is the total complement of proteins present in a cell. |
| Metaproteomics | The study of all protein samples recovered directly from environmental sources; also called community proteomics or environmental proteomics. |
| Metabolomics | Systematic study of the unique chemical fingerprints that specific cellular processes leave behind; the study of their small-molecule metabolite profiles. |
© 2016 by the author; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Watling, H. Microbiological Advances in Biohydrometallurgy. Minerals 2016, 6, 49. https://doi.org/10.3390/min6020049
Watling H. Microbiological Advances in Biohydrometallurgy. Minerals. 2016; 6(2):49. https://doi.org/10.3390/min6020049
Chicago/Turabian StyleWatling, Helen. 2016. "Microbiological Advances in Biohydrometallurgy" Minerals 6, no. 2: 49. https://doi.org/10.3390/min6020049
APA StyleWatling, H. (2016). Microbiological Advances in Biohydrometallurgy. Minerals, 6(2), 49. https://doi.org/10.3390/min6020049