
Novel Biotechnological Approaches for the Recovery of Metals from Primary and Secondary Resources

 , ,
, ,
Abstract
:
1. Introduction
2. Bioleaching
2.1. Example: Bioleaching of Copper Shale
2.2. Example: Bioleaching of Rare Earth Elements
3. Biosorption
3.1. Bacterial S-Layers
3.2. Bio-Nanomaterials
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Lovley, D.R.; Phillips, E.J.P. Reduction of uranium by Desulfovibrio desulfuricans. Appl. Environ. Microbiol. 1992, 58, 850–856. [Google Scholar] [PubMed]
- Lovley, D.R.; Phillips, E.J.P.; Gorby, Y.A.; Landa, E.R. Microbial reduction of uranium. Nature 1991, 350, 413–416. [Google Scholar] [CrossRef]
- Barkay, T.; Schaefer, J. Metal and radionuclide bioremediation: Issues, coniderations and potentials. Curr. Opin. Microbiol. 2001, 4, 318–323. [Google Scholar] [PubMed]
- Iwamoto, T.; Nasu, M. Current bioremediation practice and perspective. Biosci. Bioeng. 2001, 92, 1–8. [Google Scholar] [CrossRef]
- Botero, A.E.C.; Torem, M.L.; de Mesquita, L.M.S. Fundamental studies of Rhodococcus opacus as a biocollector of calcite and magnesite. Miner. Eng. 2007, 20, 1026–1032. [Google Scholar] [CrossRef]
- Natarajan, K.A.; Deo, N. Utility of microorganisms in mineral beneficiation. Met. Mater. Process. 1998, 10, 55–65. [Google Scholar]
- Patra, P.; Natarajan, K.A. Microbially induced flocculation and flotation for separation of chalcopyrite from quartz and calcite. Int. J. Miner. Process. 2004, 74, 143–155. [Google Scholar] [CrossRef]
- Curtis, S.B.; Hewitt, J.; MacGiliivray, R.T.A.; Dunbar, W.S. Biomining with bacteriophage: Selectivity of displayed peptides for naturally occurring sphalerite and chalcopyrite. Biotechnol. Bioeng. 2009, 102, 644–650. [Google Scholar] [CrossRef] [PubMed]
- Johnson, D.B. Development and application of biotechnologies in the metal mining industry. Environ. Sci. Pollut. Res. 2013, 20, 7768–7776. [Google Scholar] [CrossRef] [PubMed]
- Rawlings, D.E. Heavy metal mining using microbes. Annu. Rev. Microbiol. 2002, 56, 65–91. [Google Scholar] [CrossRef] [PubMed]
- Rawlings, D.E. Biomining: Theory, Microbes and Industrial Processes; Springer/Landes Biosciences: Georgetown, TX, USA, 1997. [Google Scholar]
- Jain, N.; Sharma, D.K. Biohydrometallurgy for nonsulfidic minerals—A review. Geomicrobiol. J. 2004, 21, 135–144. [Google Scholar] [CrossRef]
- Veglio, F.; Beolchini, F.; Gasbarro, A.; Toro, L.; Ubaldini, S.; Abbruzzese, C. Batch and semi-continuous tests in the bioleaching of manganiferous minerals by heterotrophic mixed microorganisms. Int. J. Miner. Process. 1997, 50, 255–273. [Google Scholar] [CrossRef]
- Kutschke, S.; Guezennec, A.G.; Hedrich, S.; Schippers, A.; Borg, G.; Kamradt, A.; Gouin, J.; Giebner, F.; Schopf, S.; Schlomann, M.; et al. Bioleaching of Kupferschiefer blackshale—A review including perspectives of the Ecometals project. Miner. Eng. 2015, 75, 116–125. [Google Scholar] [CrossRef]
- Borg, G.; Piestrzynski, A.; Bachmann, G.; Püttmann, W.; Walther, S.; Fiedler, M. An overview of the European Kupferschiefer deposits. Econ. Geol. Spec. Publ. 2012, 16, 455–486. [Google Scholar]
- Kostudis, S.; Bachmann, K.; Kutschke, S.; Pollmann, K.; Gutzmer, J. Leaching of copper from Kupferschiefer by glutamic acid and heterotrophic bacteria. Miner. Eng. 2015, 75, 38–44. [Google Scholar] [CrossRef]
- D'Hugues, P.; Norris, P.R.; Hallberg, K.B.; Sanchez, F.; Langwaldt, J.; Grotowski, A.; Chmielewski, T.; Groudev, S.; Consortium, B. Bioshale FP6 European project: Exploiting black shale ores using biotechnologies? Miner. Eng. 2008, 21, 111–120. [Google Scholar] [CrossRef]
- D'Hugues, P.; Spolaore, P.; Consortium, B. Biohydrometallurgy applied to exploitation of black shale resources: Overview of bioshale FP6 European project. Trans. Nonferr. Met. Soc. China 2008, 18, 1485–1490. [Google Scholar] [CrossRef]
- Spolaore, P.; Joulian, C.; Gouin, J.; Ibanez, A.; Auge, T.; Morin, D.; d'Hugues, P. Bioleaching of an organic-rich polymetallic concentrate using stirred-tank technology. Hydrometallurgy 2009, 99, 137–143. [Google Scholar] [CrossRef]
- Sposito, G. The Chemistry of Soils; Oxford University Press: Oxford, UK, 2008. [Google Scholar]
- Serebryanaya, M.Z.; Petrova, L.N. Complexing as a mechanism of biodegradation of industrial waste, containing manganese dioxide. In Biohydrometallurgical Processing. Volume II; Jerez, C.A., Vargas, T., Tolado, H., Wiertz, J.V., Eds.; University of Chile Press: Santiago, Chile, 1995; pp. 351–357. [Google Scholar]
- Kostudis, S.; Kutschke, S.; Pollmann, K. Mobilisation of metals from Kupferschiefer sensu strictu in neutral to alkaline environment using bulk and biotechnologically-produced organic acids. In Proceedings of the 8th International Symposium on Biohydrometallurgy, Biohydrometallurgy 2016, Falmouth, UK, 20–22 June 2016.
- Aung, K.M.M.; Ting, Y.-P. Bioleaching of spent fluid catalytic cracking catalyst using aspergillus niger. J. Biotechnol. 2005, 116, 159–170. [Google Scholar] [CrossRef] [PubMed]
- Bharadwaj, A.; Ting, Y.-P. Bioleaching of spent hydrotreating catalyst by acidophilic thermophile acidianus brierleyi: Leaching mechanism and effect of decoking. Bioresour. Technol. 2013, 130, 673–680. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.-Y.; Ting, Y.-P. Metal extraction from municipal solid waste (MSW) incinerator fly ash—Chemical leaching and fungal bioleaching. Enzym. Microb. Technol. 2006, 38, 839–847. [Google Scholar] [CrossRef]
- Rieuwerts, J.S.; Thornton, I.; Farago, M.E.; Ashmore, M.R. Factors influencing metal bioavailability in soils: Preliminary investigations for the development of a critical loads approach for metals. Chem. Speciat. Bioavailab. 1998, 10, 61–75. [Google Scholar] [CrossRef]
- Golev, A.; Scott, M.; Erskine, P.D.; Ali, S.H.; Ballantyne, G.R. Rare Earths supply chains: Current status, constraints and opportunities. Resour. Policy 2014, 41, 52–59. [Google Scholar] [CrossRef]
- Schüler, D.; Buchert, M.; Liu, R.; Dittrich, S.; Merz, C. Study on Rare Earths and Their Recycling; Öko-Institut e.V.: Freiburg, Germany, 2011. [Google Scholar]
- Schwartz, W.; Naveke, R. Biological leaching of lowgrade ores with heterotrophic microorganisms. Metall 1980, 34, 847–850. [Google Scholar]
- Iske, U.; Glombitza, F. Possibilities for the application of biotechnological processes for the production of metals from ores, minerals and industrial-wastes. ChemInform 1988, 40, 464–468. [Google Scholar] [CrossRef]
- Glombitza, F.; Iske, U.; Bullmann, M. Mikrobielle Laugung von Seltenen Erdelementen und Spurenelementen. Bioengineering 1988, 4, 37–43. (In German) [Google Scholar]
- Talasova, I.I.; Khavski, N.N.; Khairullina, R.T.; Karavaiko, G.L.; Dudeney, A.W.L. Red mud leaching with fungal metabolites. In Biohydrometallurgical Processing, Volume II; Jerez, C.A., Vargas, T., Tolado, H., Wiertz, J.V., Eds.; University of Chile Press: Santiago, Chile, 1995; pp. 379–384. [Google Scholar]
- Hassanien, A.G.; Desouky, O.A.N.; Hussien, S.S.E. Bioleaching of some Rare Earth Elements from Egyptian monazite using Aspergillus ficuum and Pseudomonas aeruginosa. Walailak J. Sci. Technol. 2014, 11, 809–823. [Google Scholar]
- Shin, D.; Kim, J.; Kim, B.S.; Jeong, J.; Lee, J.C. Use of phosphate solubilizing bacteria to leach Rare Earth Elements from monazite-bearing ore. Minerals 2015, 5, 189–202. [Google Scholar] [CrossRef]
- Hopfe, S.; Flemming, K.; Lehmann, F.; Möckel, R.; Kutschke, S.; Pollmann, K. Leaching of rare earth elements from fluorescent powder using the tea fungus Kombucha. Appl. Microbiol. Biotechnol. 2016; submitted. [Google Scholar]
- Jayabalan, R.; Malini, K.; Sathishkumar, M.; Swaminathan, K.; Yun, S.-E. Biochemical characteristics of tea fungus produced during Kombucha fermentation. Food Sci. Biotechnol. 2010, 19, 843–847. [Google Scholar] [CrossRef]
- Marsh, A.J.; O'Sullivan, O.; Hill, C.; Ross, R.P.; Cotter, P.D. Sequence-based analysis of the bacterial and fungal compositions of multiple Kombucha (tea fungus) samples. Food Microbiol. 2014, 38, 171–178. [Google Scholar] [CrossRef] [PubMed]
- Volesky, B. Biosorption and me. Water Res. 2007, 41, 4017–4029. [Google Scholar] [CrossRef] [PubMed]
- Volesky, B.; Holan, Z.R. Biosorption of heavy metals. Biotechnol. Prog. 1995, 11, 235–250. [Google Scholar] [CrossRef] [PubMed]
- Das, N. Recovery of precious metals through biosorption—A review. Hydrometallurgy 2010, 103, 180–189. [Google Scholar] [CrossRef]
- Pollmann, K.; Raff, J.; Merroun, M.; Fahmy, K.; Selenska-Pobell, S. Metal binding by bacteria from uranium mining waste piles and its potential applications. Biotechnol. Adv. 2006, 24, 58–68. [Google Scholar] [CrossRef] [PubMed]
- He, J.S.; Chen, J.P. A comprehensive review on biosorption of heavy metals by algal biomass: Materials, performances, chemistry, and modeling simulation tools. Bioresour. Technol. 2014, 160, 67–78. [Google Scholar] [CrossRef] [PubMed]
- Daubert, L.N.; Brennan, R.A. Passive remediation of acid mine drainage using crab shell chitin. Environ. Eng. Sci. 2007, 24, 1475–1480. [Google Scholar] [CrossRef]
- Salamun, N.; Triwahyono, S.; Jalil, A.A.; Matsuura, T.; Salleh, N.F.M. Acid-vacuo heat treated low cost banana stems fiber for efficient biosorption of Hg(II). RSC Adv. 2015, 5, 14129–14137. [Google Scholar] [CrossRef]
- Kuroda, K.; Ueda, M. Engineering of microorganisms towards recovery of rare metal ions. Appl. Microbiol. Biotechnol. 2010, 87, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Bruhlmann, F.; Richins, R.D.; Mulchandani, A. Engineering of improved microbes and enzymes for bioremediation. Curr. Opin. Biotech. 1999, 10, 137–141. [Google Scholar] [CrossRef]
- Nian, R.; Kim, D.S.; Thuong, N.; Tan, L.H.; Kim, C.W.; Yoo, I.K.; Choe, W.S. Chromatographic biopanning for the selection of peptides with high specificity to Pb2+ from phage displayed peptide library. J. Chromatogr. A 2010, 1217, 5940–5949. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.L.; Rosi, N.L. Peptide-based methods for the preparation of nanostructured inorganic materials. Angew. Chem. Int. Ed. 2010, 49, 1924–1942. [Google Scholar] [CrossRef] [PubMed]
- Sarikaya, M.; Tamerler, C.; Jen, A.K.Y.; Schulten, K.; Baneyx, F. Molecular biomimetics: Nanotechnology through biology. Nat. Mater. 2003, 2, 577–585. [Google Scholar] [CrossRef] [PubMed]
- Sára, M.; Sleytr, U.B. S-layer proteins. J. Bacteriol. 2000, 182, 859–868. [Google Scholar] [CrossRef] [PubMed]
- Schultze-Lam, S.; Harauz, G.; Beveridge, T.J. Participation of a cyanobacterial S-layer in fine-grain mineral formation. J. Bacteriol. 1992, 174, 7971–7981. [Google Scholar] [PubMed]
- Phoenix, V.R.; Renaut, R.W.; Jones, B.; Ferris, F.G. Bacterial S-layer preservation and rare arsenic-antimony-sulphide bioimmobilization in siliceous sediments from champagne pool hot spring, Waiotapu, New Zealand. J. Geol. Soc. Lond. 2005, 162, 323–331. [Google Scholar] [CrossRef]
- Schultze-Lam, S.; Beveridge, T.J. Nucleation of celestite and strontianite on a cyanobacterial S-layer. Appl. Environ. Microbiol. 1994, 60, 447–453. [Google Scholar] [PubMed]
- Wang, X.H.; Schröder, H.; Schlossmacher, U.; Müller, W.E.G. Organized bacterial assemblies in manganese nodules: Evidence for a role of S-layers in metal deposition. Geo-Mar. Lett. 2009, 29, 85–91. [Google Scholar] [CrossRef]
- Bertini, I.; Gelis, I.; Katsaros, N.; Luchinat, C.; Provenzani, A. Tuning the affinity for lanthanides of calcium binding proteins. Biochemistry 2003, 42, 8011–8021. [Google Scholar] [CrossRef] [PubMed]
- Fahmy, K.; Merroun, M.; Pollmann, K.; Raff, J.; Savchuk, O.; Hennig, C.; Selenska-Pobell, S. Secondary structure and Pd(II) coordination in S-layer proteins from Bacillus sphaericus studied by infrared and X-ray absorption spectroscopy. Biophys. J. 2006, 91, 996–1007. [Google Scholar] [CrossRef] [PubMed]
- Jankowski, U.; Merroun, M.L.; Selenska-Pobell, S.; Fahmy, K. S-layer protein from Lysinibacillus sphaericus JG-A12 as matrix for Au(III) sorption and Au-nanoparticle formation. Spectrosc-Int. J. 2010, 24, 177–181. [Google Scholar] [CrossRef]
- Merroun, M.; Raff, J.; Rossberg, A.; Hennig, C.; Reich, T.; Selenska-Pobell, S. Complexation of uranium by cells and S-layer sheets of Bacillus sphaericus JG-A12. Appl. Environ. Microbiol. 2005, 71, 5532–5543. [Google Scholar] [CrossRef] [PubMed]
- Lederer, F.L.; Weinert, U.; Guenther, T.J.; Raff, J.; Weiss, S.; Pollmann, K. Identification of multiple putative S-layer genes partly expressed by Lysinibacillus sphaericus JG-B53. Microbiology 2013, 159, 1097–1108. [Google Scholar] [CrossRef] [PubMed]
- Pollmann, K.; Raff, J.; Schnorpfeil, M.; Radeva, G.; Selenska-Pobell, S. Novel surface layer protein genes in Bacillus sphaericus associated with unusual insertion elements. Microbiology 2005, 151, 2961–2973. [Google Scholar] [CrossRef] [PubMed]
- Raff, J.; Merroun, M.; Rossberg, A.; Soltmann, U.; Selenska-Pobell, S. Interactions of the U Mining Waste Pile Isolate Bacillus sphaericus JG-A12 with U. In Water rock interaction; Wanty, R.B., Seal, R.R., Eds.; Balkema Publishers: Boca Raton, FL, USA, 2004; pp. 697–701. [Google Scholar]
- Suhr, M.; Raff, J.; Pollmann, K. Au-interaction of Slp1 polymers and monolayer from Lysinibacillus sphaericus JG-B53—QCM-D, ICP-MS and AFM as tools for biomolecule-metal studies. JoVE 2016, 107, e53572–e53572. [Google Scholar] [CrossRef] [PubMed]
- Suhr, M.; Unger, N.; Viacava, K.E.; Gunther, T.J.; Raff, J.; Pollmann, K. Investigation of metal sorption behavior of Slp1 from Lysinibacillus sphaericus JG-B53: A combined study using QCM-D, ICP-MS and AFM. BioMetals 2014, 27, 1337–1349. [Google Scholar] [CrossRef] [PubMed]
- Raff, J.; Soltmann, U.; Matys, S.; Selenska-Pobell, S.; Boettcher, H.; Pompe, W. Biosorption of uranium and copper by biocers. Chem. Mater. 2003, 15, 240–244. [Google Scholar] [CrossRef]
- Soltmann, U.; Raff, J.; Selenska-Pobell, S.; Matys, S.; Pompe, W.; Böttcher, H. Biosorption of heavy metals by sol-gel immobilized Bacillus sphaericus cells, spores and S-layers. J. Sol-Gel Sci. Technol. 2002, 26, 1209–1212. [Google Scholar] [CrossRef]
- Lederer, F.L.; Günther, T.; Raff, J.; Pollmann, K. E. coli filament formation induced by heterologous S-layer expression. Bioeng. Bugs 2011, 2, 178–181. [Google Scholar] [CrossRef] [PubMed]
- Lederer, F.L.; Günther, T.J.; Flemming, K.; Raff, J.; Fahmy, K.; Springer, A.; Pollmann, K. Heterologous expression of the surface-layer-like protein Sllb induces the formation of long filaments of Escherichia coli consisting of protein-stabilized outer membrane. Microbiology 2010, 156, 3584–3595. [Google Scholar] [CrossRef] [PubMed]
- Pollmann, K.; Matys, S. Construction of an S-layer protein exhibiting modified self-assembling properties and enhanced metal binding capacities. Appl. Microbiol. Biotechnol. 2007, 75, 1079–1085. [Google Scholar] [CrossRef] [PubMed]
- Sleytr, U.B.; Schuster, B.; Egelseer, E.M.; Pum, D. S-layers: Principles and applications. FEMS Microbiol. Rev. 2014, 38, 823–864. [Google Scholar] [CrossRef] [PubMed]
- Merroun, M.; Rossberg, A.; Hennig, C.; Scheinost, A.; Selenska-Pobell, S. Spectroscopic characterization of gold nanoparticles formed by cells and S-layer protein of Bacillus sphaericus JG-A12. Mater. Sci. Eng. C 2007, 27, 288–292. [Google Scholar] [CrossRef]
- Wahl, R.; Mertig, M.; Raff, J.; Selenska-Pobell, S.; Pompe, W. Electron-beam induced formation of highly ordered palladium and platinum nanoparticle arrays on the S-layer of Bacillus sphaericus NCTC9602. Adv. Mater. 2001, 13, 736–740. [Google Scholar] [CrossRef]
- Creamer, N.; Mikheenko, I.; Yong, P.; Deplanche, K.; Sanyahumbi, D.; Wood, J.; Pollmann, K.; Merroun, M.; Selenska-Pobell, S.; Macaskie, L. Novel supported Pd hydrogenation bionanocatalyst for hybrid homogeneous/heterogeneous catalysis. Catal. Today 2007, 128, 80–87. [Google Scholar] [CrossRef]
- Creamer, N.J.; Deplanche, K.; Snape, T.J.; Mikheenko, I.P.; Yong, P.; Samyahumbi, D.; Wood, J.; Pollmann, K.; Selenska-Pobell, S.; Macaskie, L.E. A biogenic catalyst for hydrogenation, reduction and selective dehalogenation in non-aqueous solvents. Hydrometallurgy 2008, 94, 138–143. [Google Scholar] [CrossRef]
- Pollmann, K.; Merroun, M.; Raff, J.; Hennig, C.; Selenska-Pobell, S. Manufacturing and characterization of Pd-nanoparticles formed on immobilized bacterial cells. Lett. Appl. Microbiol. 2006, 43, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Lakatos, M.; Matys, S.; Raff, J.; Pompe, W. Colorimetric As(V) detection based on S-layer functionalized gold nanoparticles. Talanta 2015, 144, 241–246. [Google Scholar] [CrossRef] [PubMed]
- Schmoock, C.; Bornick, H.; Vogel, M.; Lehmann, F.; Kutschke, S.; Raff, J.; Dittmar, T.; Worch, E. S-layer proteins as possible immobilization matrix for photocatalysts-OH radical scavenging capacity and protein stability. J. Photochem. Photobiol. A Chem. 2014, 277, 12–18. [Google Scholar] [CrossRef]
- Raff, J.; Pollmann, K.; Günther, T.; Marquard, A.; Katzschner, B.; Matys, S.; Pompe, W. Verwendung eines Biokompositmaterials zur Entfernung von Arsenverunreinigungen aus Wasser und Verfahren. Patent DE102011006753B3, 24 May 2012. [Google Scholar]

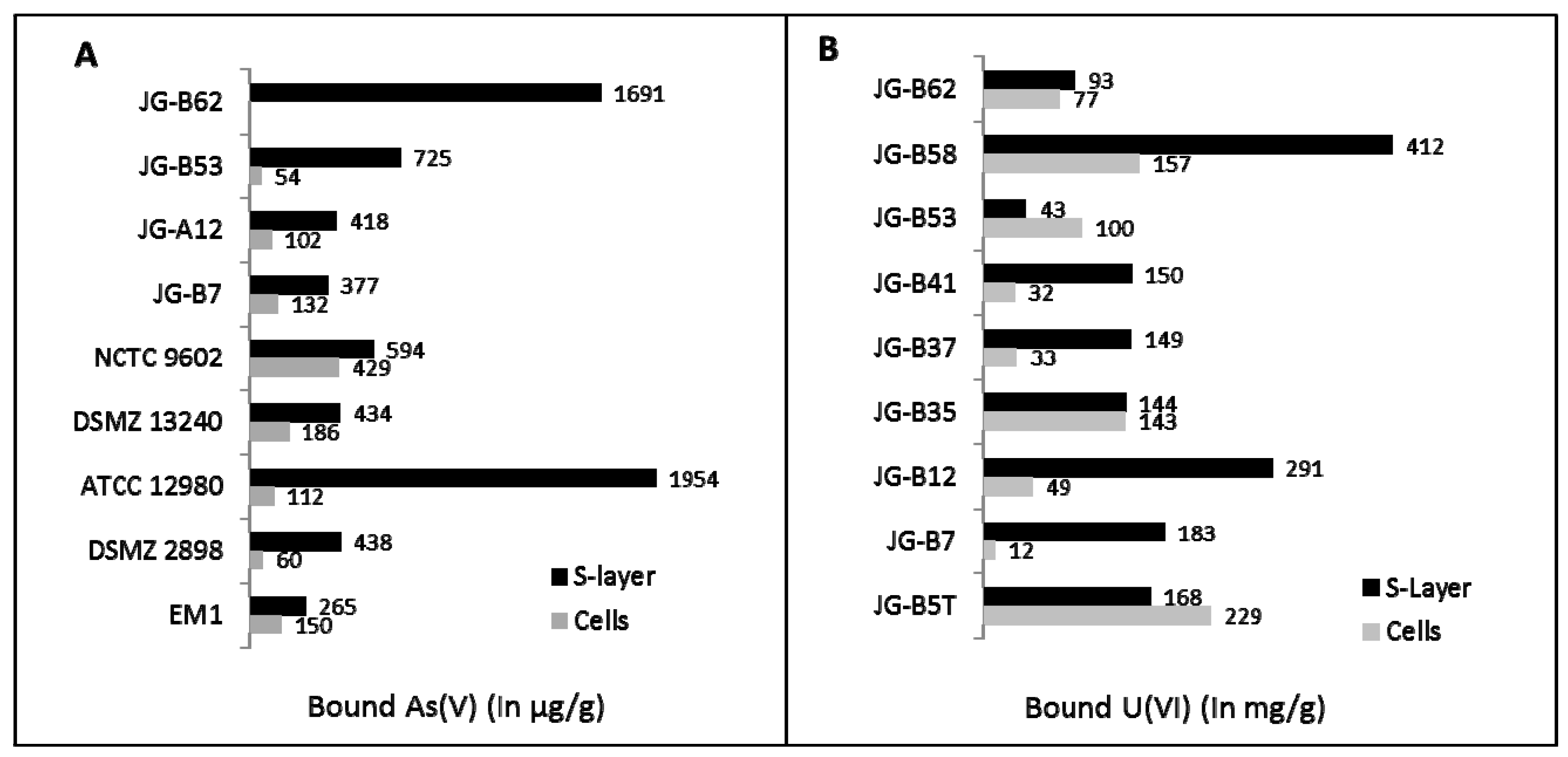

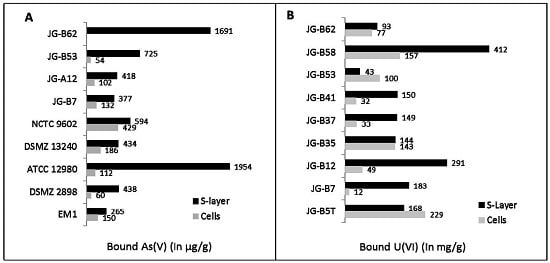
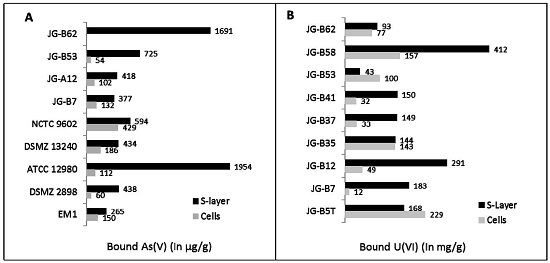{kind=link}
{kind=link}
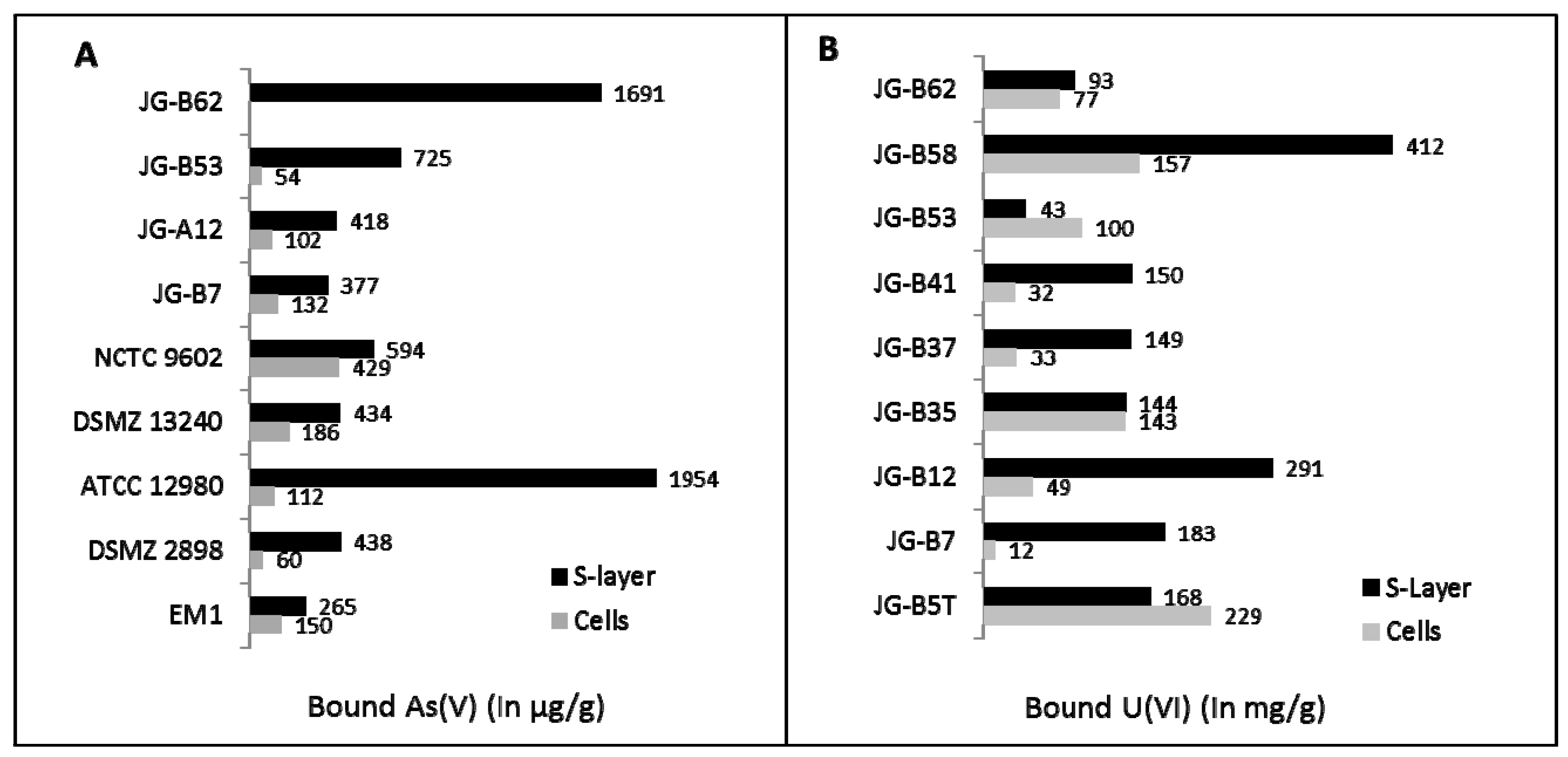{kind=link}
| Sorption Material | Binding of As(V) (in mg/g) |
|---|---|
| Dried magnetite (Fe3O4) | 2.45 |
| Magnetite suspended in water | 0.29 |
| Magnetite suspended in acetone | 0.52 |
| S-layer/magnetite composite | 5.28 |
| Dried TiO2 | 1.23 |
| TiO2 suspended in water | 0.88 |
| S-layer/TiO2 composite | 3.54 |
| FerroSorp | 2.76 |
| S-layer of L. sphaericus JG-A12 | 0.42 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pollmann, K.; Kutschke, S.; Matys, S.; Kostudis, S.; Hopfe, S.; Raff, J. Novel Biotechnological Approaches for the Recovery of Metals from Primary and Secondary Resources. Minerals 2016, 6, 54. https://doi.org/10.3390/min6020054
Pollmann K, Kutschke S, Matys S, Kostudis S, Hopfe S, Raff J. Novel Biotechnological Approaches for the Recovery of Metals from Primary and Secondary Resources. Minerals. 2016; 6(2):54. https://doi.org/10.3390/min6020054
Chicago/Turabian StylePollmann, Katrin, Sabine Kutschke, Sabine Matys, Sophias Kostudis, Stefanie Hopfe, and Johannes Raff. 2016. "Novel Biotechnological Approaches for the Recovery of Metals from Primary and Secondary Resources" Minerals 6, no. 2: 54. https://doi.org/10.3390/min6020054
APA StylePollmann, K., Kutschke, S., Matys, S., Kostudis, S., Hopfe, S., & Raff, J. (2016). Novel Biotechnological Approaches for the Recovery of Metals from Primary and Secondary Resources. Minerals, 6(2), 54. https://doi.org/10.3390/min6020054