
Bioavailability of Colloidal Iron to Heterotrophic Bacteria in Sediments, and Effects on the Mobility of Colloid-Associated Metal(loid)s

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sediment Sampling and Characteristics
2.2. Colloid Extraction and Characterization
2.3. Bacterial Strain Extraction and Identification
2.4. Alteration Experiments: The Bacteria-Colloid-Solution Simplified Micro-Ecosystem
2.4.1. Colloid Immobilization in Silica Gel
2.4.2. Experimental Conditions
2.4.3. Solution Analysis
3. Results
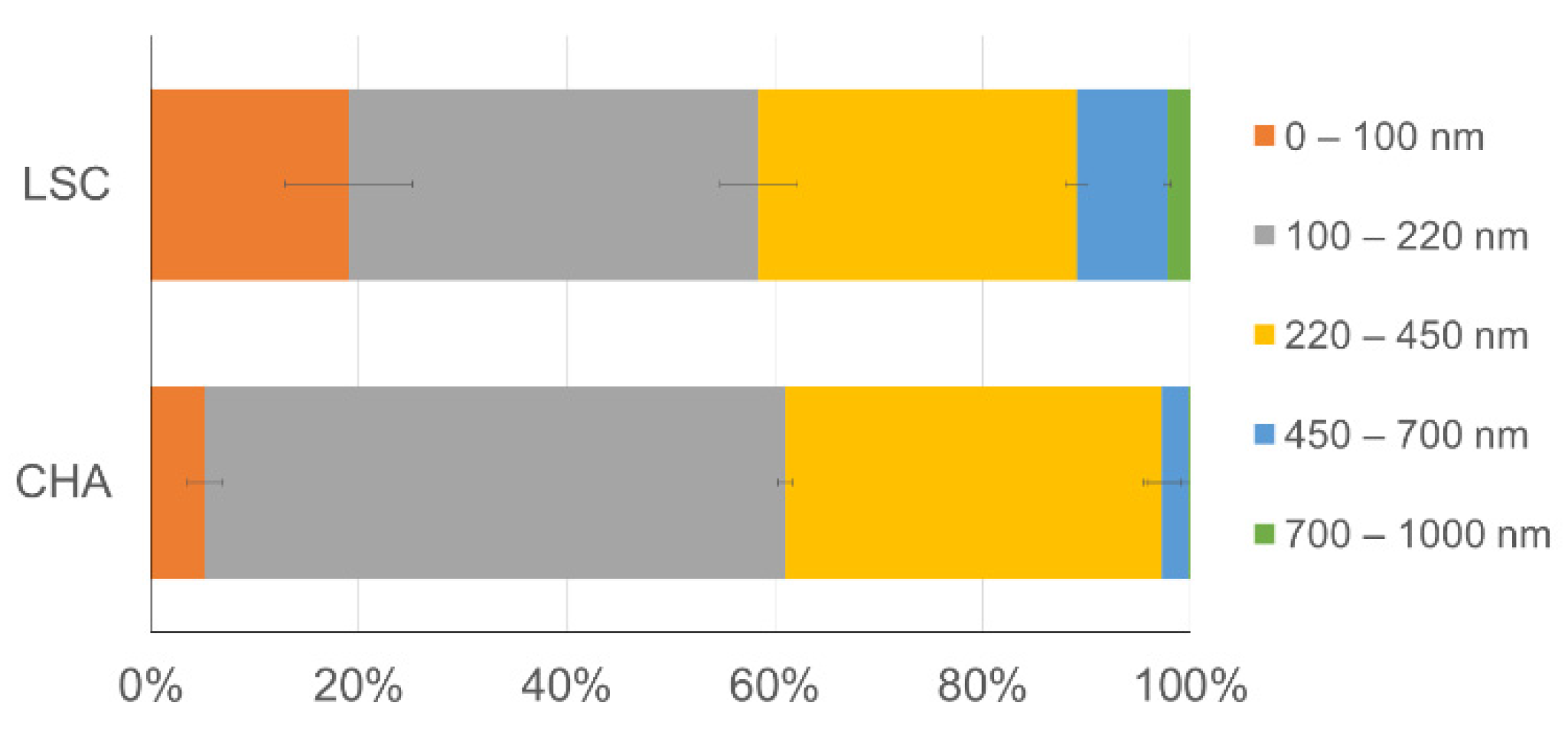
3.1. Size and Chemical Composition of the Water-Mobilizable Submicrometric Fraction of Sediments
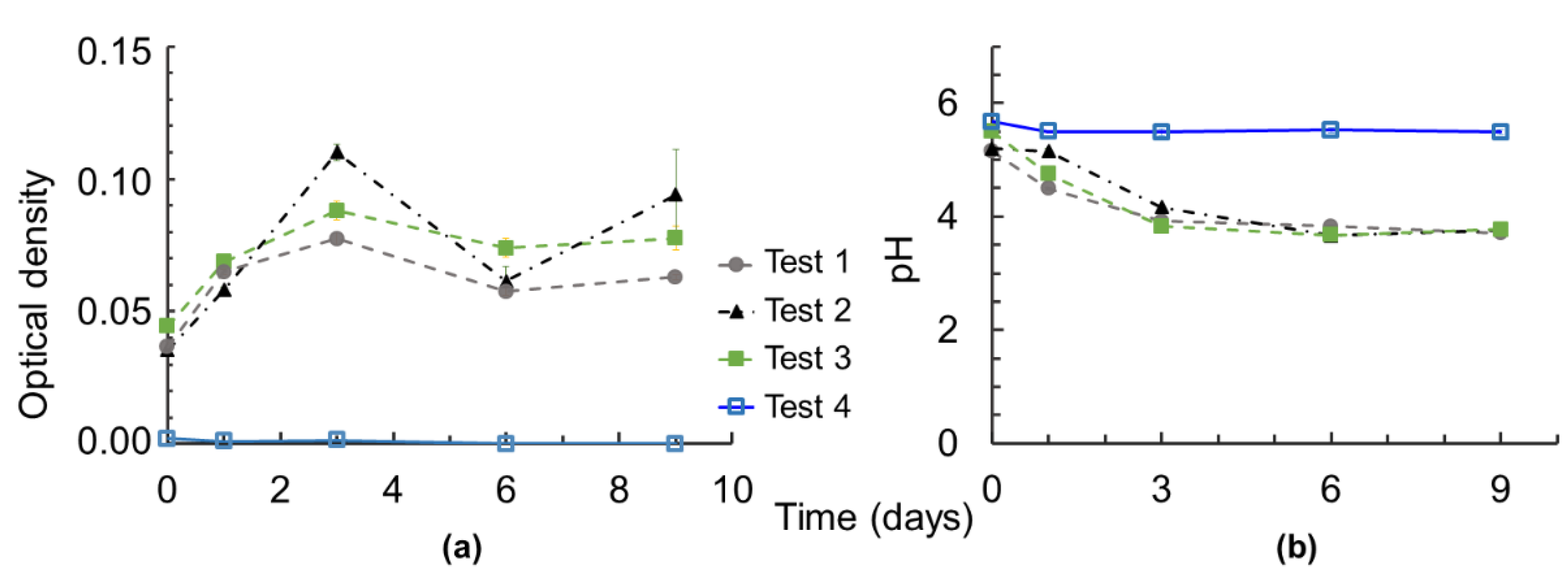
3.2. Set-Up Validation
3.3. Microbial Solubilization of Sedimentary Colloids with Time
3.3.1. Bacterial Growth
3.3.2. Iron Release in the Solution as a Function of Time
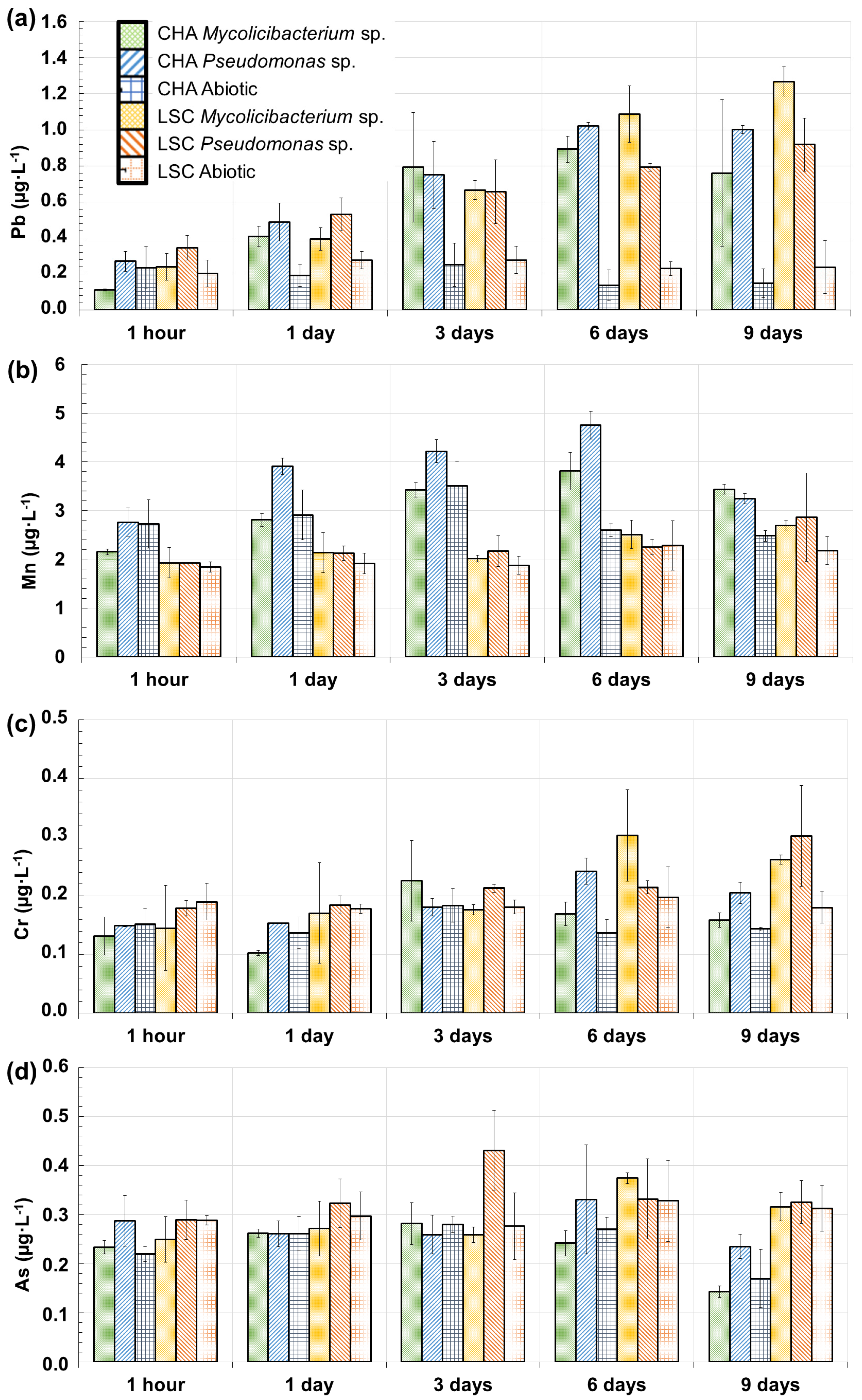
3.3.3. Trace Elements Released in the Solution as a Function of Time: Mn, Pb, Cr and As
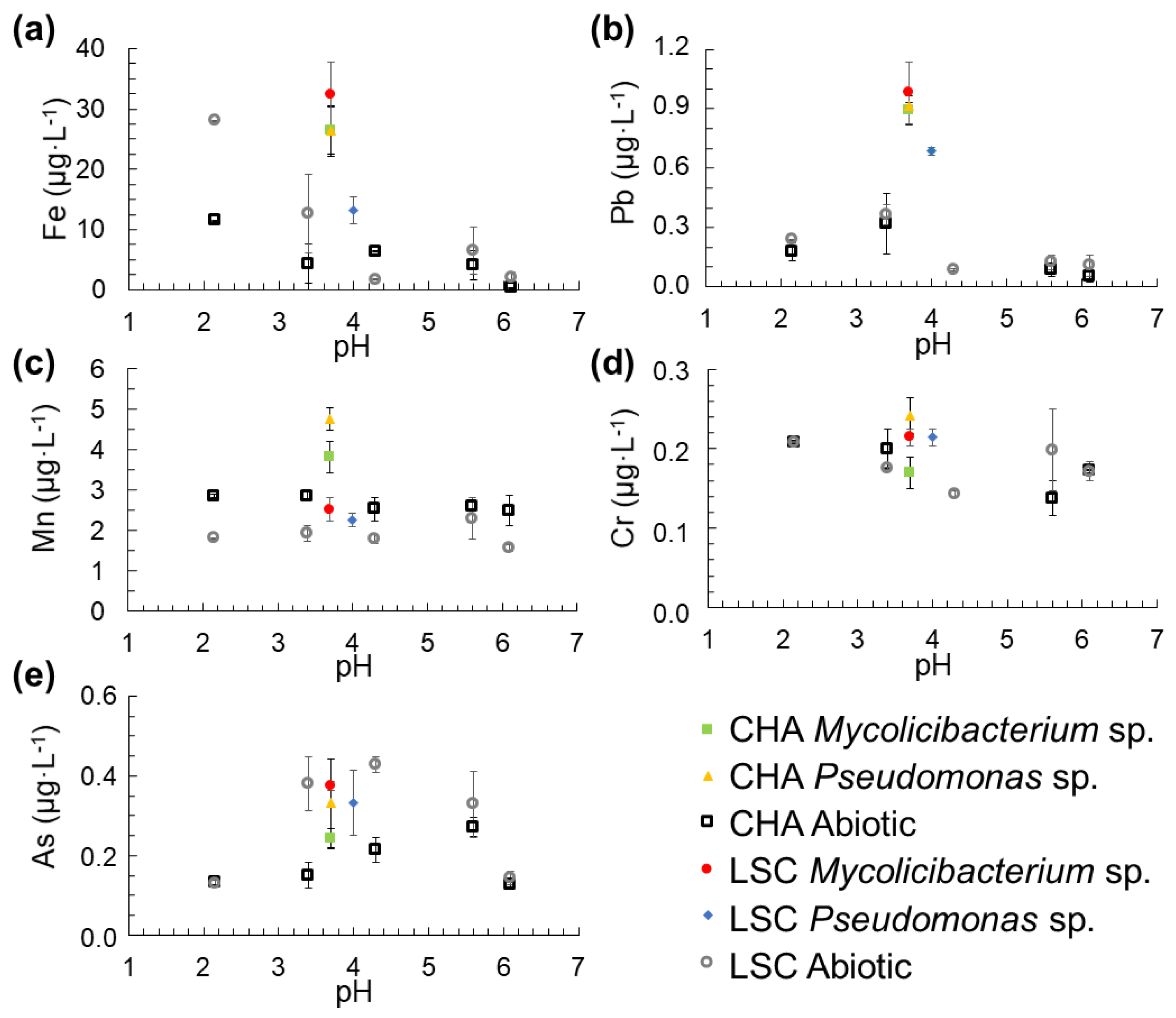
3.4. Release of Fe, Mn, Pb, Cr and As into the Solution from the Colloids as a Function of pH
4. Discussions
4.1. Quality of the Submicrometric Fraction Recovered from Bottom Sediments
4.2. Encapsulation of Sedimentary Colloids in Porous Silica Gel
- (a)
- No microbial growth and no acidification of the growth media were observed under “abiotic” conditions (Test 4, Table 1 and Figure 2), reflecting the assured sterility of the bioassay during incubation. The sterility had to be checked, as recovered sedimentary colloids (fraction <1 µm) may have contained indigenous microorganisms that settled with the sediments in the aquatic system. It has been reported that the concentration of bacteria in bottom sediments can be 10–1000 times higher than in the overlying water, and that bacterial activity and abundance are generally highest in the surface sediment and decrease with sediment depth [41,42,43,44]. Since surface sediments were collected for colloid extraction and bacteria tend to attach to smaller particles because a larger surface area provides them with more attachment sites [45,46,47], inaccurate sterilization may therefore impair the effect of the tested bacteria by the action of the bacteria recovered during colloid extraction.
- (b)
- The silica gels were confirmed to be biocompatible. Silica gels were synthesized by acid-catalyzed hydrolysis and base catalyzed condensation processes of tetraethyl orthosilicate in a closed system, which involves the release of ethanol, which can be toxic for bacteria. The presence of such residue (if any) after rinsing had to be checked. However, the greater bacterial growth in the presence of SG and HSG rather than with no solid (comparison of Tests 2 and 3 with Test 1), suggests a nontoxicity of the gels, and even a promoting effect of the presence of solid surfaces, such as silica gels in this case.
- (c)
- There was no negative effect of the presence of colloids embedded in silica gels was observed on the bacterial growth (comparison of Test 2 with Test 3, Table 1), so no toxicity effect of metal(loid)s appeared at this level and time of exposure. This has to be checked, as the tested colloids contained potentially toxic elements such as Pb and As (Table 2, Figure 6).
- (d)
- No negative effect on bacterial growth was measured under restricted dissolved Fe conditions in the presence of HSG (comparison of Test 2 with Test 3, Table 1), indicating that bacteria can manage their growth using elements coming from sedimentary colloids.
- (e)
- There was no spontaneous release of elements from colloids embedded in silica gel. Silica gel is considered to be chemically and mechanically inert and non-degradable by the microorganisms [48]. The use of a sol-gel process allows for the creation of a mineral hybrid material whose porosity depends on the synthesis parameters. In our study, TEOS hydrolysis was obtained at highly acidic conditions (pH < 2) and through polymerization by increasing the pH to about 5. We did not assess the pore’s size and its connectivity, but no spontaneous release of elements from the three-dimensional silica network was observed under abiotic conditions (Test 4, Table 1 and Figure 2). Since encapsulated suspensions consisted of fractions 0.2–1 µm (large colloids) and <0.2 m (small colloids and dissolved) in size, the lack of the spontaneous release of elements from the silica matrix suggests that the silica gels used had a pore size that did not allow colloids to be released from them, and that the dissolved fractions (if there were any) was washed out during the gel synthesis procedure.
- (f)
- Note that due to the diffusion through the gel, equilibration with the solution may not be instantaneous and this may have an impact on the kinetics of the elements released to the solution. However, the equilibration appeared relatively fast, within an hour, as indicated by the stable pH and elemental concentrations under abiotic conditions (Figure 2 and Figure 6).
4.3. Bioavailability of Colloidal Iron for Bacteria
4.4. Solubilization of Colloids and the Impact on the Release of Pb, Mn, Cr and As
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Kristensen, P.; Whalley, C.; Zal, F.N.N.; Christiansen, T. European Waters Assessment of Status and Pressures 2018. EEA Rep. 2018, 7, 85. [Google Scholar]
- Jeppesen, E.; Søndergaard, M.; Jensen, J.P.; Havens, K.E.; Anneville, O.; Carvalho, L.; Coveney, M.F.; Deneke, R.; Dokulil, M.T.; Foy, B. Lake Responses to Reduced Nutrient Loading—An Analysis of Contemporary Long-term Data from 35 Case Studies. Freshw. Biol. 2005, 50, 1747–1771. [Google Scholar] [CrossRef]
- Søndergaard, M.; Jeppesen, E.; Lauridsen, T.L.; Skov, C.; Van Nes, E.H.; Roijackers, R.; Lammens, E.; Portielje, R. Lake Restoration: Successes, Failures and Long-term Effects. J. Appl. Ecol. 2007, 44, 1095–1105. [Google Scholar] [CrossRef]
- Søndergaard, M.; Jensen, J.P.; Jeppesen, E. Role of Sediment and Internal Loading of Phosphorus in Shallow Lakes. Hydrobiologia 2003, 506, 135–145. [Google Scholar] [CrossRef]
- Schroeder, H.; Duester, L.; Fabricius, A.-L.; Ecker, D.; Breitung, V.; Ternes, T.A. Sediment Water (Interface) Mobility of Metal(Loid)s and Nutrients under Undisturbed Conditions and during Resuspension. J. Hazard. Mater. 2020, 394, 122543. [Google Scholar] [CrossRef]
- Maavara, T.; Chen, Q.; Van Meter, K.; Brown, L.E.; Zhang, J.; Ni, J.; Zarfl, C. River Dam Impacts on Biogeochemical Cycling. Nat. Rev. Earth Environ. 2020, 1, 103–116. [Google Scholar] [CrossRef]
- Frémion, F.; Courtin-Nomade, A.; Bordas, F.; Lenain, J.-F.; Jugé, P.; Kestens, T.; Mourier, B. Impact of Sediments Resuspension on Metal Solubilization and Water Quality during Recurrent Reservoir Sluicing Management. Sci. Total Environ. 2016, 562, 201–215. [Google Scholar] [CrossRef]
- Nguyen, N.D.; Grybos, M.; Rabiet, M.; Deluchat, V. Spatial Distribution of Water-Mobilizable Colloids and Phosphorus from Dam Reservoir Sediment. In Proceedings of the Copernicus Meetings, Virtual, 19–30 April 2021. [Google Scholar]
- Lead, J.R.; Wilkinson, K.J. Aquatic Colloids and Nanoparticles: Current Knowledge and Future Trends. Environ. Chem. 2006, 3, 159–171. [Google Scholar] [CrossRef]
- Xu, H.; Ji, L.; Kong, M.; Xu, M.; Lv, X. Abundance, Chemical Composition and Lead Adsorption Properties of Sedimentary Colloids in a Eutrophic Shallow Lake. Chemosphere 2019, 218, 534–539. [Google Scholar] [CrossRef]
- Nguyen, D.N.; Grybos, M.; Rabiet, M.; Deluchat, V. How Do Colloid Separation and Sediment Storage Methods Affect Water-Mobilizable Colloids and Phosphorus? An Insight into Dam Reservoir Sediment. Colloids Surf. A: Physicochem. Eng. Asp. 2020, 606, 125505. [Google Scholar] [CrossRef]
- Saeed, H.; Hartland, A.; Lehto, N.; Baalousha, M.; Sikder, M.; Sandwell, D.; Mucalo, M.; Hamilton, D.P. Regulation of Phosphorus Bioavailability by Iron Nanoparticles in a Monomictic Lake. Sci. Rep. 2018, 8, 17736. [Google Scholar] [PubMed] [Green Version]
- Eggleton, J.; Thomas, K.V. A Review of Factors Affecting the Release and Bioavailability of Contaminants during Sediment Disturbance Events. Environ. Int. 2004, 30, 973–980. [Google Scholar] [CrossRef] [PubMed]
- Frémion, F.; Mourier, B.; Courtin-Nomade, A.; Lenain, J.-F.; Annouri, A.; Fondanèche, P.; Hak, T.; Bordas, F. Key Parameters Influencing Metallic Element Mobility Associated with Sediments in a Daily-Managed Reservoir. Sci. Total Environ. 2017, 605–606, 666–676. [Google Scholar] [CrossRef]
- Sun, W.; Cheng, K.; Sun, K.Y.; Ma, X. Microbially Mediated Remediation of Contaminated Sediments by Heavy Metals: A Critical Review. Curr. Pollut. Rep. 2021, 7, 201–212. [Google Scholar]
- Gu, Y.-G. Heavy Metal Fractionation and Ecological Risk Implications in the Intertidal Surface Sediments of Zhelin Bay, South China. Mar. Pollut. Bull. 2018, 129, 905–912. [Google Scholar]
- Ren, J.; Williams, P.N.; Luo, J.; Ma, H.; Wang, X. Sediment Metal Bioavailability in Lake Taihu, China: Evaluation of Sequential Extraction, DGT, and PBET Techniques. Environ. Sci. Pollut. Res. 2015, 22, 12919–12928. [Google Scholar]
- Cortada, U.; Hidalgo, M.C.; Martínez, J.; de la Torre, M.J. Mobility and Bioavailability of Metal(Loid)s in a Fluvial System Affected by the Mining and Industrial Processing of Pb. Geosciences 2021, 11, 167. [Google Scholar] [CrossRef]
- Zhang, C.; Ding, S.; Xu, D.; Tang, Y.; Wong, M.H. Bioavailability Assessment of Phosphorus and Metals in Soils and Sediments: A Review of Diffusive Gradients in Thin Films (DGT). Environ. Monit. Assess. 2014, 186, 7367–7378. [Google Scholar] [CrossRef]
- Lei, K.; Han, X.; Zhao, J.; Qiao, F.; Li, Z.; Yu, T. Characterization of Metal Kinetics and Bioavailability Using Diffusive Gradients in Thin Films Technique in Sediments of Taihu Lake, China. Ecotoxicol. Environ. Saf. 2016, 128, 153–160. [Google Scholar] [CrossRef]
- Gu, Y.-G. Risk Assessment of Eight Metals and Their Mixtures to Aquatic Biota in Sediments with Diffusive Gradients in Thin Films (DGT): A Case Study in Pearl River Intertidal Zone. Environ. Sci. Eur. 2021, 33, 122. [Google Scholar] [CrossRef]
- Ikem, A.; Egiebor, N.; Nyavor, K. Trace Elements in Water, Fish and Sediment from Tuskegee Lake, Southeastern USA. Water Air Soil Pollut. 2003, 149, 51–75. [Google Scholar] [CrossRef]
- Grybos, M.; Billard, P.; Desobry-Banon, S.; Michot, L.J.; Lenain, J.-F.; Mustin, C. Bio-Dissolution of Colloidal-Size Clay Minerals Entrapped in Microporous Silica Gels. J. Colloid. Interface Sci. 2011, 362, 317–324. [Google Scholar] [CrossRef] [PubMed]
- Murphy, J.; Riley, J.P. A Modified Single Solution Method for the Determination of Phosphate in Natural Waters. Anal. Chim. Acta 1962, 27, 31–36. [Google Scholar] [CrossRef]
- Ruban, V.; López-Sánchez, J.; Pardo, P.; Rauret, G.; Muntau, H.; Quevauviller, P. Harmonized Protocol and Certified Reference Material for the Determination of Extractable Contents of Phosphorus in Freshwater Sediments–A Synthesis of Recent Works. Fresenius J. Anal. Chem. 2001, 370, 224–228. [Google Scholar] [CrossRef] [PubMed]
- United States Environmental Protection Agency. Determination of Phosphorus by Semi-Automated Colorimetry, Revision 2.0; USEPA Method 365.1; United States Environmental Protection Agency: Washington, DC, USA, 1993. [Google Scholar]
- DuBois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric Method for Determination of Sugars and Related Substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
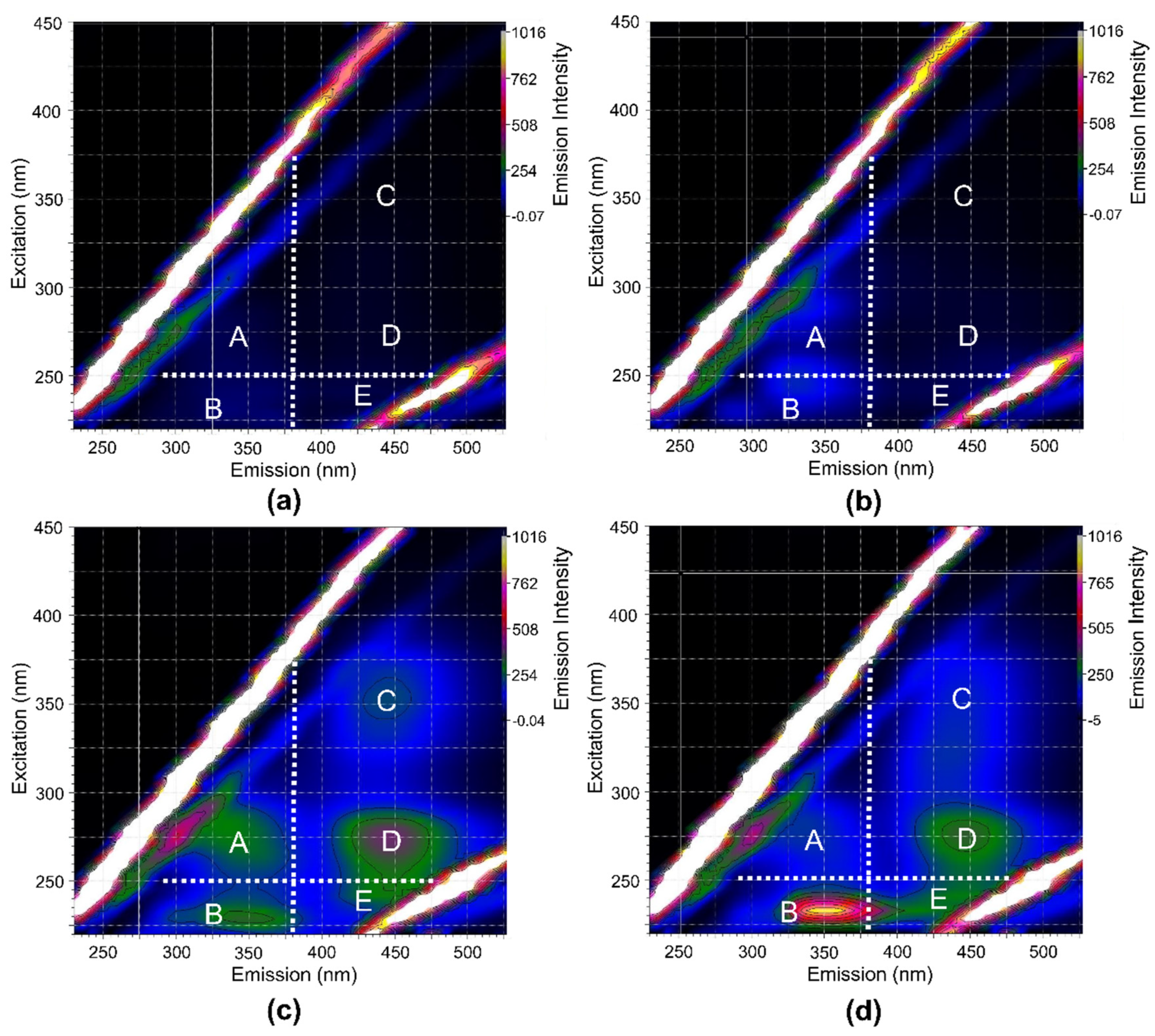
- Chen, W.; Westerhoff, P.; Leenheer, J.A.; Booksh, K. Fluorescence Excitation−Emission Matrix Regional Integration to Quantify Spectra for Dissolved Organic Matter. Environ. Sci. Technol. 2003, 37, 5701–5710. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Wang, Y.; Zhang, W.; Yu, S.; Wang, X.; Gao, N. New Advances in Fluorescence Excitation-Emission Matrix Spectroscopy for the Characterization of Dissolved Organic Matter in Drinking Water Treatment: A Review. Chem. Eng. J. 2020, 381, 122676. [Google Scholar] [CrossRef]
- Kondolf, G.M.; Gao, Y.; Annandale, G.W.; Morris, G.L.; Jiang, E.; Zhang, J.; Cao, Y.; Carling, P.; Fu, K.; Guo, Q.; et al. Sustainable Sediment Management in Reservoirs and Regulated Rivers: Experiences from Five Continents. Earths Future 2014, 2, 256–280. [Google Scholar] [CrossRef]
- Guo, X.; Zhu, X.; Yang, Z.; Ma, J.; Xiao, S.; Ji, D.; Liu, D. Impacts of Cascade Reservoirs on the Longitudinal Variability of Fine Sediment Characteristics: A Case Study of the Lancang and Nu Rivers. J. Hydrol. 2020, 581, 124343. [Google Scholar] [CrossRef]
- Rapin, A.; Rabiet, M.; Mourier, B.; Grybos, M.; Deluchat, V. Sedimentary Phosphorus Accumulation and Distribution in the Continuum of Three Cascade Dams (Creuse River, France). Environ. Sci. Pollut. Res. 2020, 27, 6526–6539. [Google Scholar] [CrossRef]
- Mahmood, Q.; Khan, A.F.; Khan, A. Chapter 25—Colloids in the Environmental Protection—Current and Future Trends. In The Role of Colloidal Systems in Environmental Protection; Fanun, M., Ed.; Elsevier: Amsterdam, The Netherlands, 2014; pp. 635–677. ISBN 978-0-444-63283-8. [Google Scholar]
- Kretzschmar, R.; Barmettler, K.; Grolimund, D.; Yan, Y.; Borkovec, M.; Sticher, H. Experimental Determination of Colloid Deposition Rates and Collision Efficiencies in Natural Porous Media. Water Resour. Res. 1997, 33, 1129–1137. [Google Scholar] [CrossRef]
- Kaplan, D.I.; Sumner, M.E.; Bertsch, P.M.; Adriano, D.C. Chemical Conditions Conducive to the Release of Mobile Colloids from Ultisol Profiles. Soil Sci. Soc. Am. J. 1996, 60, 269–274. [Google Scholar] [CrossRef]
- Klitzke, S.; Lang, F.; Kaupenjohann, M. Increasing PH Releases Colloidal Lead in a Highly Contaminated Forest Soil. Eur. J. Soil Sci. 2008, 59, 265–273. [Google Scholar] [CrossRef]
- Sun, Y.; Pan, D.; Wei, X.; Xian, D.; Wang, P.; Hou, J.; Xu, Z.; Liu, C.; Wu, W. Insight into the Stability and Correlated Transport of Kaolinite Colloid: Effect of PH, Electrolytes and Humic Substances. Environ. Pollut. 2020, 266, 115189. [Google Scholar] [CrossRef] [PubMed]
- Maximova, N.; Dahl, O. Environmental Implications of Aggregation Phenomena: Current Understanding. Curr. Opin. Colloid. Interface Sci. 2006, 11, 246–266. [Google Scholar] [CrossRef]
- Parrello, D.; Zegeye, A.; Mustin, C.; Billard, P. Siderophore-Mediated Iron Dissolution from Nontronites Is Controlled by Mineral Cristallochemistry. Front. Microbiol. 2016, 7, 423. [Google Scholar] [CrossRef]
- Oulkadi, D.; Balland-Bolou-Bi, C.; Billard, P.; Kitzinger, G.; Parrello, D.; Mustin, C.; Banon, S. Interactions of Three Soil Bacteria Species with Phyllosilicate Surfaces in Hybrid Silica Gels. FEMS Microbiol. Lett. 2014, 354, 37–45. [Google Scholar] [CrossRef]
- Fischer, H.; Wanner, S.C.; Pusch, M. Bacterial Abundance and Production in River Sediments as Related to the Biochemical Composition of Particulate Organic Matter (POM). Biogeochemistry 2002, 61, 37–55. [Google Scholar] [CrossRef]
- Haglund, A.-L.; Törnblom, E.; Boström, B.; Tranvik, L. Large Differences in the Fraction of Active Bacteria in Plankton, Sediments, and Biofilm. Microb. Ecol. 2002, 43, 232–241. [Google Scholar] [CrossRef]
- Haglund, A.-L.; Lantz, P.; Törnblom, E.; Tranvik, L. Depth Distribution of Active Bacteria and Bacterial Activity in Lake Sediment. FEMS Microb. Ecol. 2003, 46, 31–38. [Google Scholar] [CrossRef]
- Pachepsky, Y.; Shelton, D. Escherichia Coli and Fecal Coliforms in Freshwater and Estuarine Sediments. Crit. Rev. Environ. Sci. Technol. 2011, 41, 1067–1110. [Google Scholar] [CrossRef]
- Goulder, R. Attached and Free Bacteria in an Estuary with Abundant Suspended Solids. J. Appl. Bacteriol. 1977, 43, 399–405. [Google Scholar] [CrossRef]
- Guber, A.K.; Pachepsky, Y.A.; Shelton, D.R.; Yu, O. Effect of Bovine Manure on Fecal Coliform Attachment to Soil and Soil Particles of Different Sizes. Appl. Environ. Microbiol. 2007, 73, 3363–3370. [Google Scholar] [CrossRef] [Green Version]
- Wu, T.; Zhai, C.; Zhang, J.; Zhu, D.; Zhao, K.; Chen, Y. Study on the Attachment of Escherichia Coli to Sediment Particles at a Single-Cell Level: The Effect of Particle Size. Water 2019, 11, 819. [Google Scholar] [CrossRef] [Green Version]
- Meunier, C.F.; Dandoy, P.; Su, B.-L. Encapsulation of Cells within Silica Matrixes: Towards a New Advance in the Conception of Living Hybrid Materials. J. Colloid. Interface Sci. 2010, 342, 211–224. [Google Scholar] [CrossRef] [PubMed]
- Kalinowski, B.E.; Liermann, L.J.; Givens, S.; Brantley, S.L. Rates of Bacteria-Promoted Solubilization of Fe from Minerals: A Review of Problems and Approaches. Chem. Geol. 2000, 169, 357–370. [Google Scholar] [CrossRef]
- Hider, R.C.; Kong, X. Chemistry and Biology of Siderophores. Nat. Prod. Rep. 2010, 27, 637. [Google Scholar] [CrossRef]
- Rogers, J.R.; Bennett, P.C. Mineral Stimulation of Subsurface Microorganisms: Release of Limiting Nutrients from Silicates. Chem. Geol. 2004, 203, 91–108. [Google Scholar] [CrossRef]
- Hersman, L.; Lloyd, T.; Sposito, G. Siderophore-Promoted Dissolution of Hematite. Geochim. Cosmochim. Acta 1995, 59, 3327–3330. [Google Scholar] [CrossRef]
- Oulkadi, D.; Balland-Bolou-Bi, C.; Michot, L.J.; Grybos, M.; Billard, P.; Mustin, C.; Banon, S. Bioweathering of Nontronite Colloids in Hybrid Silica Gel: Implications for Iron Mobilization. J. Appl. Microbiol. 2014, 116, 325–334. [Google Scholar] [CrossRef]
- Saha, R.; Saha, N.; Donofrio, R.S.; Bestervelt, L.L. Microbial Siderophores: A Mini Review. J. Basic Microbiol. 2013, 53, 303–317. [Google Scholar] [CrossRef] [PubMed]
- Perez, A.; Rossano, S.; Trcera, N.; Verney-Carron, A.; Rommevaux, C.; Fourdrin, C.; Agnello, A.C.; Huguenot, D.; Guyot, F. Direct and Indirect Impact of the Bacterial Strain Pseudomonas Aeruginosa on the Dissolution of Synthetic Fe(III)- and Fe(II)-Bearing Basaltic Glasses. Chem. Geol. 2019, 523, 9–18. [Google Scholar] [CrossRef] [Green Version]
- Gottselig, N.; Nischwitz, V.; Meyn, T.; Amelung, W.; Bol, R.; Halle, C.; Vereecken, H.; Siemens, J.; Klumpp, E. Phosphorus Binding to Nanoparticles and Colloids in Forest Stream Waters. Vadose Zone J. 2017, 16, 1–12. [Google Scholar] [CrossRef]
- Kumari, S.; Mahapatra, S.; Das, S. Ca-Alginate as a Support Matrix for Pb (II) Biosorption with Immobilized Biofilm Associated Extracellular Polymeric Substances of Pseudomonas Aeruginosa N6P6. Chem. Eng. J. 2017, 328, 556–566. [Google Scholar] [CrossRef]
- Mullen, M.; Wolf, D.; Ferris, F.; Beveridge, T.; Flemming, C.; Bailey, G. Bacterial Sorption of Heavy Metals. Appl. Environ. Microbiol. 1989, 55, 3143–3149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tikhonov, V.; Drozdova, O.; Cheptsov, V.; Demin, V. Sorption of Dissolved Organic Matter by Freshwater Bacterioplankton. EDP Sciences. In Proceedings of the E3S Web of Conferences, Virtual, 3 June 2021; Volume 265. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Test N° | Presence of Bacteria 1 | BHM 2 | HSG or SG 3 | Fe Source |
|---|---|---|---|---|
| 1 | YES | YES | No HSG or SG | Dissolved Fe 4 |
| 2 | YES | YES | SG | Dissolved Fe 4 |
| 3 | YES | YES | HSG | Colloids 5 |
| 4 | NO | YES | HSG | Colloids 5 |
| Fraction | Al (mg·L−1) | Fe (mg·L−1) | Mn (mg·L−1) | Cr (µg·L−1) | As (µg·L−1) | Pb (µg·L−1) | Corg (mg·L−1) | Ntot (mg·L−1) | Ptot (mg·L−1) |
|---|---|---|---|---|---|---|---|---|---|
| CHA < 1 µm | 3.20 ± 0.18 | 1.95 ± 0.28 | 0.26 ± 0.10 | 2.72 ± 0.20 | 3.3 ± 0.7 | 5.04 ± 1.05 | 4.09 ± 0.17 | 3.04 ± 1.26 | 0.29 ± 0.01 |
| CHA < 0.2 µm | 0.76 ± 0.12 | 0.60 ± 0.16 | 0.25 ± 0.12 | 0.8 ± 0.1 | 2.22 ± 0.32 | 0.95 ± 0.24 | 3.94 ± 0.47 | 3.21 ± 0.95 | 0.10 ± 0.01 |
| CHA 0.2–1 µm | 2.44 ± 0.16 | 1.34 ± 0.15 | <LQ | 1.91 ± 0.11 | 1.10 ± 0.53 | 4.08 ± 0.84 | 0.14 ± 0.31 | <LQ | 1.0 ± 0.1 |
| LSC < 1 µm | 27.16 ± 3.56 | 14.12 ± 1.84 | 0.09 ± 0.01 | 26.82 ± 3.36 | 8.55 ± 1.14 | 13.8 ± 1.73 | 12.35 ± 0.8 | 1.69 ± 0.31 | 5.20 ± 0.4 |
| LSC < 0.2 µm | 0.03 ± 0.11 | 0.06 ± 0.07 | 0.00 ± 0.00 | 0.17 ± 0.15 | 1.13 ± 0.05 | 0.07 ± 0.07 | 3.78 ± 0.80 | 0.91 ± 0.07 | 0.04 ± 0.01 |
| LSC 0.2–1 µm | 27.06 ± 3.54 | 14.06 ± 1.84 | 0.09 ± 0.01 | 26.64 ± 3.41 | 7.42 ± 1.16 | 13.73 ± 1.66 | 8.57 ± 1.14 | 0.78 ± 0.3 | 5.2 ± 0.4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grybos, M.; Masson, D.; Gorgeon, P.; Fondanèche, P.; Martin, N.; Dupuy, F.; Joussein, E.; Robin, V. Bioavailability of Colloidal Iron to Heterotrophic Bacteria in Sediments, and Effects on the Mobility of Colloid-Associated Metal(loid)s. Minerals 2022, 12, 812. https://doi.org/10.3390/min12070812
Grybos M, Masson D, Gorgeon P, Fondanèche P, Martin N, Dupuy F, Joussein E, Robin V. Bioavailability of Colloidal Iron to Heterotrophic Bacteria in Sediments, and Effects on the Mobility of Colloid-Associated Metal(loid)s. Minerals. 2022; 12(7):812. https://doi.org/10.3390/min12070812
Chicago/Turabian StyleGrybos, Malgorzata, Delphine Masson, Pauline Gorgeon, Patrice Fondanèche, Nicolas Martin, Fabrice Dupuy, Emmanuel Joussein, and Valentin Robin. 2022. "Bioavailability of Colloidal Iron to Heterotrophic Bacteria in Sediments, and Effects on the Mobility of Colloid-Associated Metal(loid)s" Minerals 12, no. 7: 812. https://doi.org/10.3390/min12070812
APA StyleGrybos, M., Masson, D., Gorgeon, P., Fondanèche, P., Martin, N., Dupuy, F., Joussein, E., & Robin, V. (2022). Bioavailability of Colloidal Iron to Heterotrophic Bacteria in Sediments, and Effects on the Mobility of Colloid-Associated Metal(loid)s. Minerals, 12(7), 812. https://doi.org/10.3390/min12070812