Asymmetry and Symmetry in the Beauty of Human Faces

{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Methods
2.1. Participants
2.2. Stimuli
2.3. Procedures
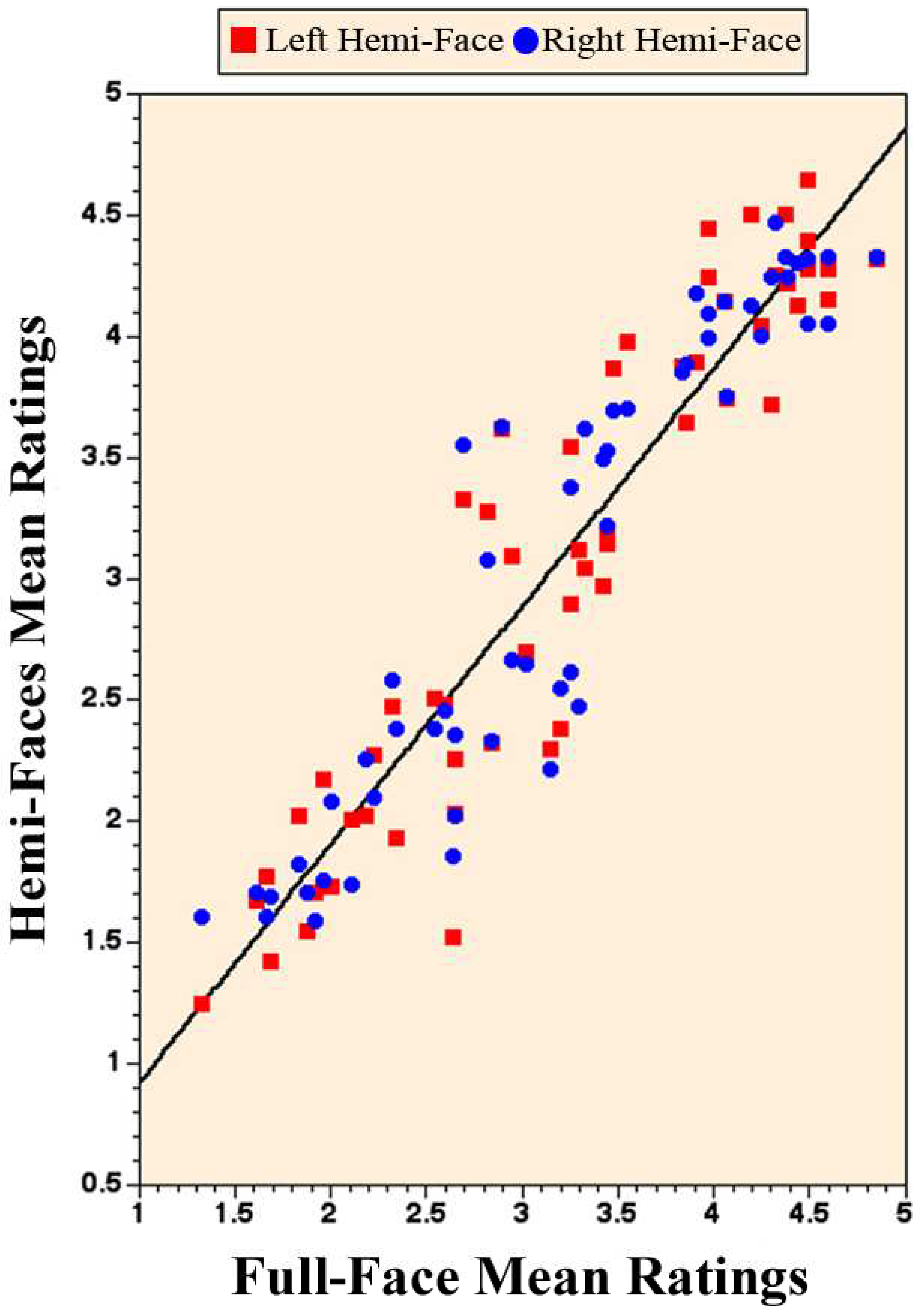
3. Results
4. Discussion
5. Conclusions
Acknowledgments
References and Notes
- Kosso, P. Symmetry arguments in physics. Stud. Hist. Philos. Sci. A 1999, 30, 479–492. [Google Scholar] [CrossRef]
- Engler, G. Aesthetics in science and art. Br. J. Aesthetics 1990, 30, 24–34. [Google Scholar] [CrossRef]
- Zee, A. Fearful symmetry: The search for beauty in modern physics; Princeton University Press: Princeton, NJ, USA, 2007. [Google Scholar]
- Engler, G. Einstein, his theories, and his aesthetic considerations. Int. Stud. Philos. Sci. 2005, 19, 21–30. [Google Scholar] [CrossRef]
- Muller, A. Chemistry: The beauty of symmetry. Science 2003, 300, 749–750. [Google Scholar] [CrossRef]
- Gould, J.L.; Gould, C.G. Sexual selection; Scientific American Library: New York, 1989. [Google Scholar]
- Cronin, H. The ant and the peacock; Cambridge University Press: Cambridge, UK, 1992. [Google Scholar]
- Jacobsen, T.; Schubotz, R.I.; Hofel, L.; Cramon, D.Y.V. Brain correlates of aesthetic judgment of beauty. NeuroImage 2006, 29, 276–285. [Google Scholar] [CrossRef]
- Saito, Y. The Japanese aesthetics of imperfection and insufficiency. J. Aesthet. Art Critic. 1997, 55, 377–385. [Google Scholar] [CrossRef]
- Wicks, R. The idealization of contingency in traditional Japanese aesthetics. J. Aesthet. Edu. 2005, 39, 88–101. [Google Scholar] [CrossRef]
- Juniper, A. Wabi Sabi: The Japanese art of impermanence; Tuttle Publishing: Tokyo, Japan, 2003. [Google Scholar]
- Marra, M.F. A history of modern Japanese aesthetics; University of Hawaii Press: Honolulu, HI, USA, 2001. [Google Scholar]
- Grammer, K.; Thornhill, R. Human (Homo sapiens) facial attractiveness and sexual selection: The role of symmetry and averageness. J. Com. Psychol. 1994, 108, 233–242. [Google Scholar] [CrossRef]
- Zaidel, D.W.; Chen, A.C.; German, C. She is not a beauty even when she smiles: Possible evolutionary basis for a relationship between facial attractiveness and hemispheric specialization. Neuropsychologia 1995, 33, 649–655. [Google Scholar] [CrossRef]
- Zaidel, D.W.; Deblieck, C. Attractiveness of natural faces compared to computer constructed perfectly symmetrical faces. Int. J. Neurosci. 2007, 117, 423–431. [Google Scholar] [CrossRef]
- Babcock, L.E. Asymmetry in the fossil record. Euro. Rev. 2005, 13, 135–143. [Google Scholar] [CrossRef]
- Palmer, A.R. Waltzing with asymmetry. Biol. Sci. 1996, 46, 518–532. [Google Scholar] [CrossRef]
- Stevens, M.; Castor-Perry, S.A.; Price, J.R.F. The protective value of conspicuous signals is not impaired by shape, size, or position asymmetry. Behav. Ecol. 2008, 20, 96–102. [Google Scholar] [CrossRef]
- Takahashi, M.; Arita, H.; Hiraiwa-Hasegawa, M.; Hasegawa, T. Peahens do not prefer peacocks with more elaborate trains. Anim. Behav. 2008, 75, 1209–1219. [Google Scholar] [CrossRef]
- Dufour, K.W.; Weatherhead, P.J. Reproductive consequences of bilateral asymmetry for individual male red-winged blackbirds. Behav. Ecol. 1998, 9, 232–242. [Google Scholar] [CrossRef]
- Midgley, J.J.; Johnson, S.D. Some pollinators do not prefer symmetrically marked or shaped daisy (Asteraceae) flowers. Evol. Ecol. 1998, 12, 123–126. [Google Scholar] [CrossRef]
- Neville, A.C. Animal asymmetry; Edward Arnold: London, UK, 1976. [Google Scholar]
- Sperry, R.W. Lateral specialization in the surgically separated hemispheres. In Neurosciences Third Study Program; Schmitt, F., Worden, F., Eds.; MIT Press: Cambridge, MA, USA, 1974; Vol. 3. [Google Scholar]
- Zaidel, D.W. Different concepts and meaning systems in the left and right hemispheres. Psychol. Learn. Motiv. 2000, 40, 1–21. [Google Scholar]
- Serrien, D.J.; Ivry, R.B.; Swinnen, S.P. Dynamics of hemispheric specialization and integration in the context of motor control. Nat. Rev. Neurosci. 2006, 7, 160–167. [Google Scholar] [CrossRef]
- Zilhao, J. The emergence of ornaments and art: An archaeological perspective on the origins of “behavioral modernity”. J. Archaeol. Res. 2007, 15, 1–54. [Google Scholar] [CrossRef]
- Mace, R. On becoming modern. Science 2009, 324, 1280–1281. [Google Scholar] [CrossRef]
- Zaidel, D.W. Art and brain: Insights from neuropsychology, biology and evolution. J. Anat. 2010, 216, 20–27. [Google Scholar] [CrossRef] [PubMed]
- Farkas, L.G.; Posnick, J.C.; Hreczko, T.M. Growth patterns of the face: A morphometric study. Cleft Palate Craniofac. J. 1992, 29, 308–315. [Google Scholar] [CrossRef] [PubMed]
- Ferrario, V.F.; Sforza, C.; Pogio, C.E.; Tartaglia, G. Distance from symmetry: A three-dimensional evaluation of facial asymmetry. J. Oral Maxillofac. Surg. 1994, 52, 1126–1132. [Google Scholar] [CrossRef]
- Ferrario, V.F.; Sforza, C.; Miani, A., Jr.; Serrao, G. A three-dimensional evaluation of human facial asymmetry. J. Anat. 1995, 186 (Pt 1), 103–110. [Google Scholar]
- Ferrario, V.F.; Sforza, C.; Ciusa, V.; Dellavia, C.; Tartaglia, G.M. The effect of sex and age on facial asymmetry in healthy subjects: A cross-sectional study from adolescence to mid-adulthood. J. Oral Maxillofac. Surg. 2001, 59, 382–388. [Google Scholar] [CrossRef] [PubMed]
- Ferrario, V.F.; Sforza, C.; Dellavia, C.; Tartaglia, G.M.; Colombo, A.; Caru, A. A quantitative three-dimensional assessment of soft tissue facial asymmetry of cleft lip and palate adult patients. J. Craniofac. Surg. 2003, 14, 739–746. [Google Scholar] [CrossRef] [PubMed]
- Drummond, P.D.; Mirco, N. Staring at one side of the face increases blood flow on that side of the face. Psychophysiol. 2004, 41, 281–287. [Google Scholar] [CrossRef]
- Benedicic, M.; Dolenk, V.V.; Stefanovska, A.; Bosnjak, R. Left-right asymmetry of the facial microvascular control. Clin. Auton. Res. 2006, 16, 58–60. [Google Scholar] [CrossRef]
- Sackheim, H.A.; Weiman, A.L.; Forman, B.D. Asymmetry of the face at rest: Size, area and emotional expression. Cortex 1984, 20, 165–178. [Google Scholar] [CrossRef]
- Dopson, W.G.; Beckwith, B.E.; Tucker, D.M.; Bullard-Bates, P.C. Asymmetry of facial expression in spontaneous emotion. Cortex 1984, 20, 243–251. [Google Scholar] [CrossRef]
- Borod, J.C.; Haywood, C.S.; Koff, E. Neuropsychological aspects of facial asymmetry during emotional expression: A review of the normal adult literature. Neuropsych. Rev. 1997, 7, 41–60. [Google Scholar] [CrossRef]
- Skvarilova, B. Facial asymmetry: An X-ray study. Acta Chir. Plast. 1994, 36, 89–91. [Google Scholar] [PubMed]
- Peck, S.; Peck, L.; Kataia, M. Skeletal asymmetry in esthetically pleasing faces. Angle Orthod. 1991, 61, 43–48. [Google Scholar] [PubMed]
- Wylie, D.R.; Goodale, M.A. Left sided oral asymmetries in spontaneous but not posed smiles. Neuropsychologia 1988, 26, 823–832. [Google Scholar] [CrossRef]
- Jordan, T.R.; Thomas, S.M. Hemiface contributions to hemispheric dominance in visual speech perception. Neuropsychology 2007, 21, 721–731. [Google Scholar] [CrossRef] [PubMed]
- Nicholls, M.E.; Searle, D.A. Asymmetries for the visual expression and perception of speech. Brain Lang. 2006, 97, 322–331. [Google Scholar] [CrossRef] [PubMed]
- Zaidel, D.W.; Cohen, J.A. The face, beauty, and symmetry: Perceiving asymmetry in beautiful faces. Int. J. Neurosci. 2005, 115, 1165–1173. [Google Scholar] [CrossRef]
- Wagemans, J. Characteristics and models of human symmetry detection. Trends Cogn. Sci. 1997, 1, 346–352. [Google Scholar] [CrossRef]
- Swaddle, J.P. Visual signaling by asymmetry: A review of perceptual processes. Proc. R. Soc. Lond. B Biol. Sci. 1999, 354, 1383–1393. [Google Scholar] [CrossRef]
- Enquist, M.; Arak, A. Symmetry, beauty, and evolution. Nature 1994, 372, 169–172. [Google Scholar] [CrossRef]
- Johnstone, R.A. Female preference for symmetrical males as a by-product of selection for mate recognition. Nature 1994, 372, 172–175. [Google Scholar] [CrossRef] [PubMed]
- Reis, V.A.; Zaidel, D.W. Functional asymmetry in the human face: Perception of health in the left and right sides of the face. Laterality 2001, 6, 225–231. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Zaidel, D.W.; Aarde, S.M.; Baig, K. Appearance of symmetry, beauty, and health in human faces. Brain Cogn. 2005, 57, 261–263. [Google Scholar] [CrossRef] [PubMed]
- Little, A.C.; Jones, B.C. Evidence against perceptual bias views for symmetry preferences in human faces. Proc. R. Soc. Lond. B Biol. Sci. 2003, 270, 1759–1763. [Google Scholar] [CrossRef] [PubMed]
- Rhodes, G.; Proffitt, F.; Grady, J.M.; Sumich, A. Facial symmetry and the perception of beauty. Psychon. Bull. Rev. 1998, 5, 659–669. [Google Scholar] [CrossRef]
- Knowner, R. Facial asymmetry and attractiveness judgment in developmental perspective. J. Exp. Psychol. 1996, 22, 662–675. [Google Scholar]
- Samuels, C.A.; Butterworth, G.; Roberts, T.; Graupner, L.; Hole, G. Facial aesthetics: Babies prefer attractiveness to symmetry. Perception 1994, 23, 823–831. [Google Scholar] [CrossRef]
- Scheib, J.E.; Gangestad, S.W.; Thornhill, R. Facial attractiveness, symmetry and cues of good genes. Proc. R. Soc. Lond. B Biol. Sci. 1999, 266, 1913–1917. [Google Scholar] [CrossRef]
- Springer, I.N.; Wannicke, B.; Warnke, P.H.; Zernial, O.; Wiltfang, J.; Russo, P.A.J.; Terheyden, H.; Reinhardt, A.; Wolfart, S. Visual attractiveness: Visual impact of symmetry increases significantly towards the midline. Ann. Plast. Surg. 2007, 59, 156–162. [Google Scholar] [CrossRef]
- Chen, A.C.; German, C.; Zaidel, D.W. Brain asymmetry and facial attractiveness: Beauty is not simply in the eye of the beholder. Neuropsychologia 1997, 35, 471–476. [Google Scholar] [CrossRef]
- Moller, A.P. Bumblebee preference for symmetrical flowers. Proc. Nat. Acad. Sci. USA 1995, 92, 2288–2292. [Google Scholar] [CrossRef] [PubMed]
- Langlois, J.H.; Roggman, L.A. Attractive faces are only average. Psychol. Sci. 1990, 1, 115–121. [Google Scholar] [CrossRef]
- Swaddle, J.P.; Cuthill, J.C. Asymmetry and human facial attractiveness: Symmetry may not always be beautiful. Proc. R. Soc. Lond. B Biol. Sci. 1995, 261, 111–116. [Google Scholar]
- Ritchie, M.G. The shape of female mating preferences. Proc. Nat. Acad. Sci. USA 1996, 93, 14628–14631. [Google Scholar] [CrossRef] [PubMed]
- Ellison, P.T. Beauty: In the gonads of the beholder - and the beheld. Horm. Behav. 2008, 53, 11–13. [Google Scholar] [CrossRef] [PubMed]
- Penton-Voak, I.S.; Little, A.C.; Jones, B.C.; Burt, D.M.; Tiddeman, B.P.; Perrett, D.I. Female condition influences preferences for sexual dimorphism in faces of male humans (Homo sapiens). J. Comp. Psychol. 2003, 117, 264–271. [Google Scholar] [CrossRef]
- Swaddle, J.P.; Reierson, G.W. Testosterone increases perceived dominance but not attractiveness in human males. Pro. Biol. Sci. 2002, 22, 2285–2289. [Google Scholar] [CrossRef]
- Welling, L.L.; Jones, B.C.; DeBruine, L.M.; Conway, C.A.; Law Smith, M.J.; Little, A.C.; Feinberg, D.R.; Sharp, M.A.; Al-Dujaili, E.A. Raised salivary testosterone in women is associated with increased attraction to masculine faces. Horm. Behav. 2007, 52, 156–161. [Google Scholar] [CrossRef]
- Madden, J. Sex, bowers and brains. Proc. R. Soc. Lond. B Biol. Sci. 2001, 268, 833–838. [Google Scholar] [CrossRef]
- Keagy, J.; Savard, J.-F.; Borgia, G. Male satin bowerbird problem-solving ability predicts mating success. Anim. Behav. 2009, 100, 1–9. [Google Scholar] [CrossRef]
- Cuthill, I.C.; Hiby, E.; Lloyd, E. The predation costs of symmetrical cryptic coloration. Proc. R. Soc. Lond. B Biol. Sci. 2006, 273, 1267–1271. [Google Scholar] [CrossRef] [PubMed]
- Hauser, M.D.; Akre, K. Asymmetries in the timing of facial and vocal expressions in rhesus monkeys: Implications for hemispheric specialization. Anim. Behav. 2001, 61, 391–408. [Google Scholar] [CrossRef]
- Weiss, D.; Ghazanfar, A.; Miller, C.T.; Hauser, M.D. Specialized processing of primate facial and vocal expressions: Evidence for cerebral asymmetries. In Cerebral vertebrate lateralization; Andrews, L.R.R., Ed.; Cambridge University Press: New York, NY, USA, 2002. [Google Scholar]
- Hook-Costigan, M.A.; Rogers, L.J. Lateralised use of the mouth in production of vocalizations by marmosets. Neuropsychologia 1998, 36, 1265–1273. [Google Scholar] [CrossRef]
- Fernandez-Cariba, S.; Loeches, A.; Morcillo, A.; Hopkins, W.D. Asymmetry in facial expression of emotions by chimpanzees. Neuropsychologia 2002, 40, 1523–1533. [Google Scholar] [CrossRef]
- Vallortigara, G.; Rogers, L.J. Survival with an asymmetrical brain: Advantages and disadvantages of cerebral lateralization. Behav. Brain Sci. 2005, 28, 575–589. [Google Scholar] [CrossRef] [PubMed]
- Buss, D.M. Strategies of human mating. Psychol. Topics 2006, 15, 239–260. [Google Scholar]
- Rhodes, G. The evolutionary psychology of facial beauty. Annu. Rev. Psychol. 2006, 57, 199–226. [Google Scholar] [CrossRef] [PubMed]
- Yorzinski, J.L.; Platt, M.L. Same-sex gaze attraction influences mate-choice copying in humans. PLoS ONE 2010, 5, e9115. [Google Scholar] [CrossRef]
- Little, A.C.; Jones, B.C.; Waitt, C.; Tiddman, B.P.; Feinberg, D.R.; Perrett, D.I.; Apicella, C.L.; Marlowe, F.W. Symmetry is related to sexual dimorphism in faces: Data across culture and species. PLoS ONE 2008, 7, e2106. [Google Scholar] [CrossRef] [PubMed]
- Fink, B.; Neave, N.; Manning, J.T.; Grammer, K. Facial symmetry and judgements of attractiveness, health and personality. Pers. Individ. Dif. 2006, 41, 491–499. [Google Scholar] [CrossRef]
- Jones, B.C.; DeBruine, L.M.; Little, A.C. The role of symmetry in attraction to average faces. Percept. Psychophys. 2007, 69, 1273–1277. [Google Scholar] [CrossRef] [PubMed]
- Zaidel, D.W. Brain and art: Neuro-clues from intersection of disciplines. In Neuroaesthetics; Skov, M., Vartanian, O., Eds.; Baywood: Amityville, NY, USA, 2009. [Google Scholar]



© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Zaidel, D.W.; Hessamian, M. Asymmetry and Symmetry in the Beauty of Human Faces. Symmetry 2010, 2, 136-149. https://doi.org/10.3390/sym2010136
Zaidel DW, Hessamian M. Asymmetry and Symmetry in the Beauty of Human Faces. Symmetry. 2010; 2(1):136-149. https://doi.org/10.3390/sym2010136
Chicago/Turabian StyleZaidel, Dahlia W., and Marjan Hessamian. 2010. "Asymmetry and Symmetry in the Beauty of Human Faces" Symmetry 2, no. 1: 136-149. https://doi.org/10.3390/sym2010136
APA StyleZaidel, D. W., & Hessamian, M. (2010). Asymmetry and Symmetry in the Beauty of Human Faces. Symmetry, 2(1), 136-149. https://doi.org/10.3390/sym2010136