

No Sex Differences in the Attentional Bias for the Right Side of Human Bodies

 ,
,  and
and
Abstract
1. Introduction
2. Materials and Method
2.1. Participants
2.2. Stimuli
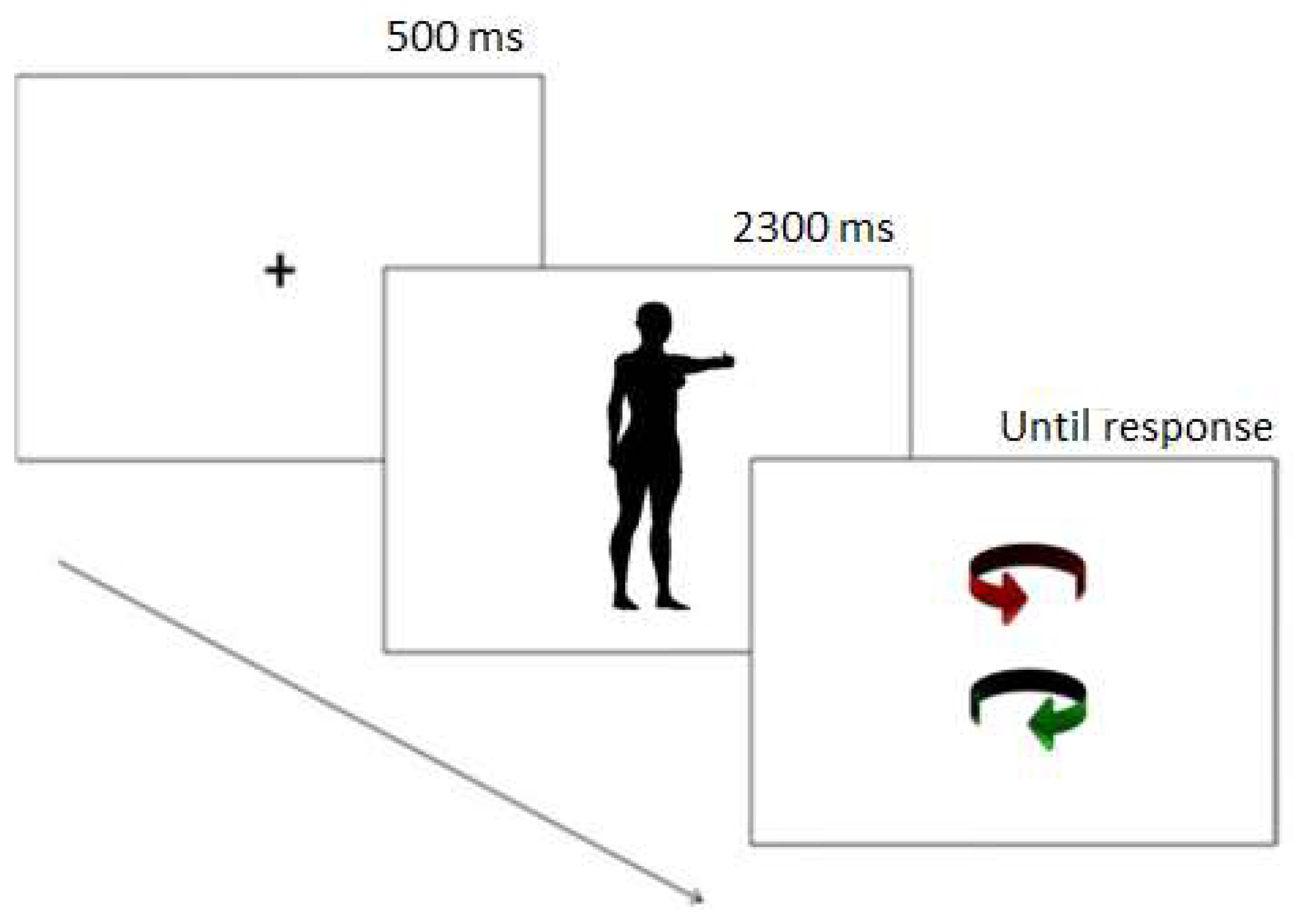
2.3. Procedure
2.4. Data Analysis
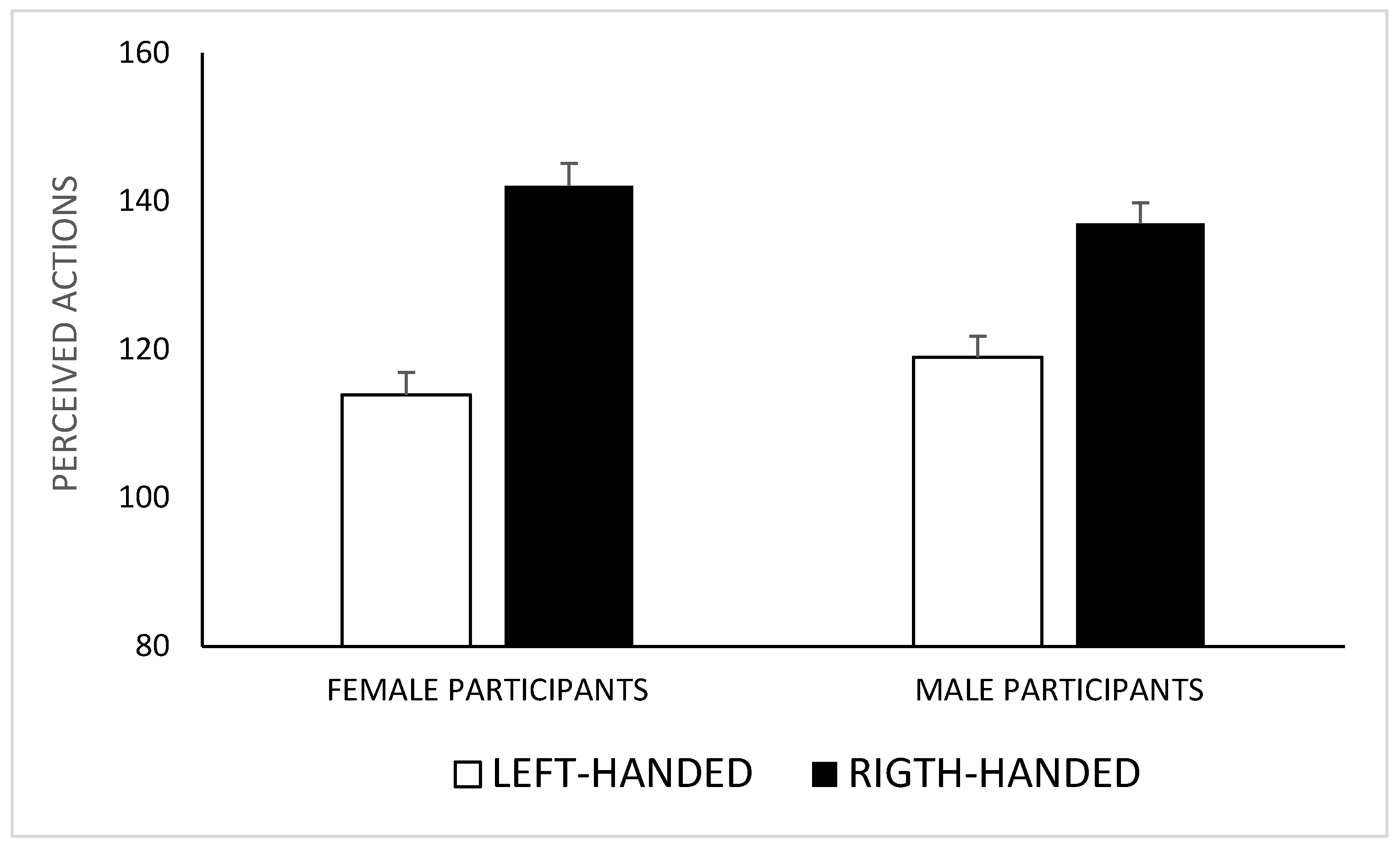
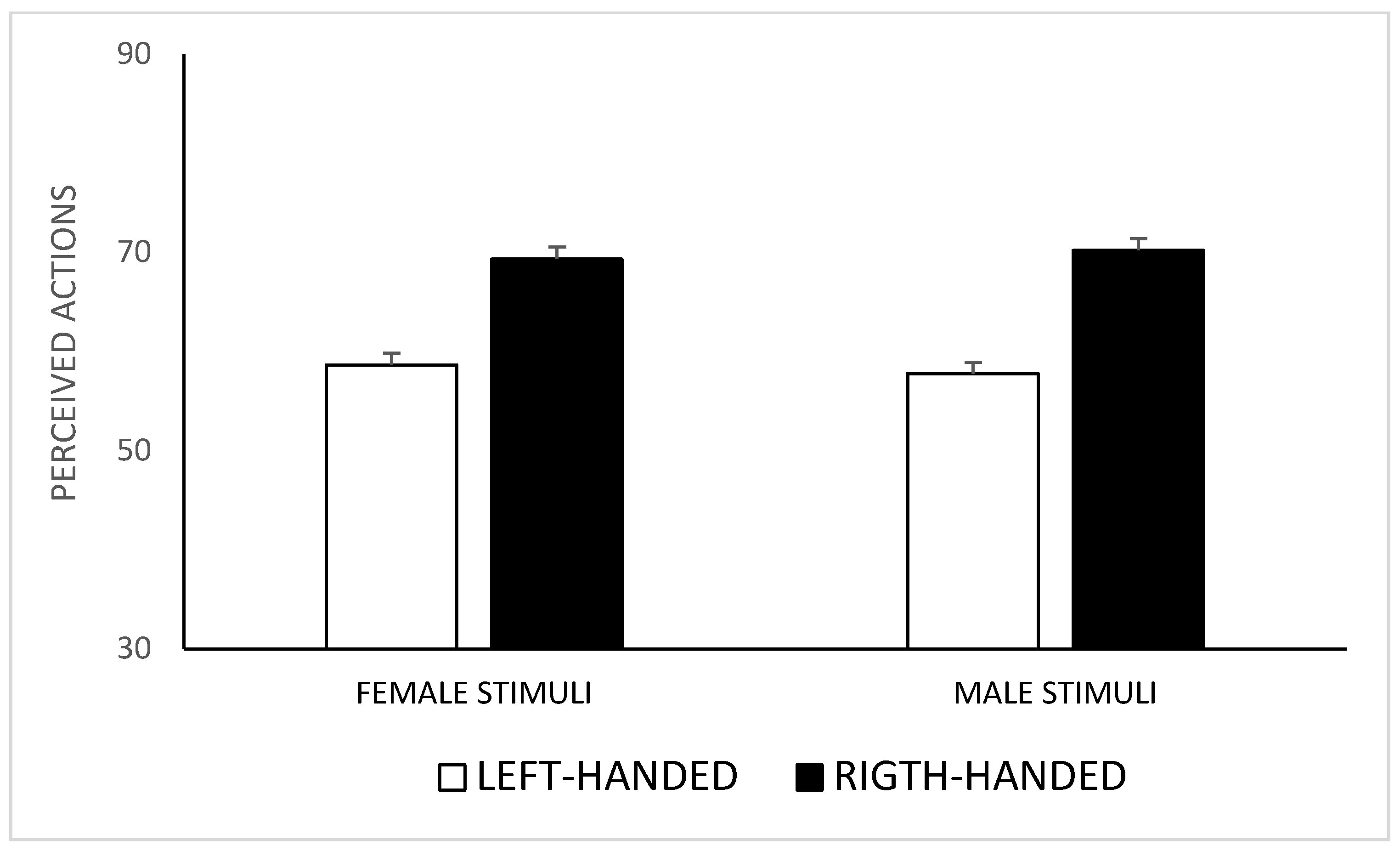
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Marzoli, D.; Menditto, S.; Lucafò, C.; Tommasi, L. Imagining Others’ Handedness: Visual and Motor Processes in the Attribution of the Dominant Hand to an Imagined Agent. Exp. Brain Res. 2013, 229, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Marzoli, D.; Lucafò, C.; Rescigno, C.; Mussini, E.; Padulo, C.; Prete, G.; D’Anselmo, A.; Malatesta, G.; Tommasi, L. Sex-Specific Effects of Posture on the Attribution of Handedness to an Imagined Agent. Exp. Brain Res. 2017, 235, 1163–1171. [Google Scholar] [CrossRef] [PubMed]
- Marzoli, D.; Mitaritonna, A.; Moretto, F.; Carluccio, P.; Tommasi, L. The Handedness of Imagined Bodies in Action and the Role of Perspective Taking. Brain Cogn. 2011, 75, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Marzoli, D.; Palumbo, R.; Di Domenico, A.; Penolazzi, B.; Garganese, P.; Tommasi, L. The Relation between Self-Reported Empathy and Motor Identification with Imagined Agents. PLoS ONE 2011, 6, e14595. [Google Scholar] [CrossRef]
- Lucafò, C.; Marzoli, D.; Prete, G.; Tommasi, L. Laterality Effects in the Spinning Dancer Illusion: The Viewing-from-above Bias Is Only Part of the Story. Br. J. Psychol. Lond. Engl. 1953 2016, 107, 698–709. [Google Scholar] [CrossRef]
- Lucafò, C.; Marzoli, D.; Padulo, C.; Troiano, S.; Pelosi Zazzerini, L.; Malatesta, G.; Amodeo, I.; Tommasi, L. Hemifield-Specific Rotational Biases during the Observation of Ambiguous Human Silhouettes. Symmetry 2021, 13, 1349. [Google Scholar] [CrossRef]
- Lucafò, C.; Marzoli, D.; Zdybek, P.; Malatesta, G.; Smerilli, F.; Ferrara, C.; Tommasi, L. The Bias toward the Right Side of Others Is Stronger for Hands than for Feet. Symmetry 2021, 13, 146. [Google Scholar] [CrossRef]
- Marzoli, D.; Lucafò, C.; Pagliara, A.; Cappuccio, R.; Brancucci, A.; Tommasi, L. Both Right- and Left-Handers Show a Bias to Attend Others’ Right Arm. Exp. Brain Res. 2015, 233, 415–424. [Google Scholar] [CrossRef]
- Marzoli, D.; Pagliara, A.; Prete, G.; Malatesta, G.; Lucafò, C.; Padulo, C.; Brancucci, A.; Tommasi, L. Lateralized Embodiment of Ambiguous Human Silhouettes: Data on Sex Differences. Data Brief 2019, 25, 104009. [Google Scholar] [CrossRef]
- Marzoli, D.; Lucafò, C.; Padulo, C.; Prete, G.; Giacinto, L.; Tommasi, L. Inversion Reveals Perceptual Asymmetries in the Configural Processing of Human Body. Front. Behav. Neurosci. 2017, 11, 126. [Google Scholar] [CrossRef]
- Marzoli, D.; Pagliara, A.; Prete, G.; Malatesta, G.; Lucafò, C.; Padulo, C.; Brancucci, A.; Tommasi, L. Hemispheric Asymmetries in the Processing of Body Sides: A Study with Ambiguous Human Silhouettes. Neurosci. Lett. 2017, 656, 114–119. [Google Scholar] [CrossRef]
- Faurie, C.; Raymond, M. Handedness, Homicide and Negative Frequency-Dependent Selection. Proc. Biol. Sci. 2005, 272, 25–28. [Google Scholar] [CrossRef]
- Grouios, G.; Tsorbatzoudis, H.; Alexandris, K.; Barkoukis, V. Do Left-Handed Competitors Have an Innate Superiority in Sports? Percept. Mot. Skills 2000, 90, 1273–1282. [Google Scholar] [CrossRef]
- Grouios, G.; Koidou, E.; Tsormpatzoudis, C.; Alexandris, K. Handedness in Sport. J. Hum. Mov. Stud. 2002, 43, 347–361. [Google Scholar]
- Raymond, M.; Pontier, D.; Dufour, A.B.; Møller, A.P. Frequency-Dependent Maintenance of Left Handedness in Humans. Proc. Biol. Sci. 1996, 263, 1627–1633. [Google Scholar] [CrossRef]
- Groothuis, T.G.G.; McManus, I.C.; Schaafsma, S.M.; Geuze, R.H. The Fighting Hypothesis in Combat: How Well Does the Fighting Hypothesis Explain Human Left-Handed Minorities? Ann. N. Y. Acad. Sci. 2013, 1288, 100–109. [Google Scholar] [CrossRef]
- Gursoy, R. Effects of Left- or Right-Hand Preference on the Success of Boxers in Turkey. Br. J. Sports Med. 2009, 43, 142–144. [Google Scholar] [CrossRef]
- Loffing, F.; Hagemann, N. Pushing through Evolution? Incidence and Fight Records of Left-Oriented Fighters in Professional Boxing History. Laterality Asymmetries Body Brain Cogn. 2015, 20, 270–286. [Google Scholar] [CrossRef]
- Richardson, T.; Gilman, R.T. Left-Handedness Is Associated with Greater Fighting Success in Humans. Sci. Rep. 2019, 9, 15402. [Google Scholar] [CrossRef]
- Cingoz, Y.E.; Gursoy, R.; Ozan, M.; Hazar, K.; Dalli, M. Research on the Relation between Hand Preference and Success in Karate and Taekwondo Sports with Regards to Gender. Adv. Phys. Educ. 2018, 8, 308. [Google Scholar] [CrossRef]
- Ziyagil, M.A.; Gursoy, R.; Dane, S.; Yuksel, R. Left-Handed Wrestlers Are More Successful. Percept. Mot. Skills 2010, 111, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Dochtermann, N.A.; Gienger, C.M.; Zappettini, S. Born to Win? Maybe, but Perhaps Only against Inferior Competition. Anim. Behav. 2014, 96, e1–e3. [Google Scholar] [CrossRef]
- Pollet, T.V.; Stulp, G.; Groothuis, T.G. Born to Win? Testing the Fighting Hypothesis in Realistic Fights: Left-Handedness in the Ultimate Fighting Championship. Anim. Behav. 2013, 86, 839–843. [Google Scholar] [CrossRef]
- Bisiacchi, P.S.; Ripoll, H.; Stein, J.; Simonet, P.; Azemar, G. Left-Handedness in Fencers: An Attentional Advantage? Percept. Mot. Skills 1985, 61, 507–513. [Google Scholar] [CrossRef]
- Harris, L.J. In Fencing, What Gives Left-Handers the Edge? Views from the Present and the Distant Past. Laterality 2010, 15, 15–55. [Google Scholar] [CrossRef]
- Lawler, T.P.; Lawler, F.H. Left-Handedness in Professional Basketball: Prevalence, Performance, and Survival. Percept. Mot. Skills 2011, 113, 815–824. [Google Scholar] [CrossRef]
- Goldstein, S.R.; Young, C.A. “ Evolutionary” Stable Strategy of Handedness in Major League Baseball. J. Comp. Psychol. 1996, 110, 164. [Google Scholar] [CrossRef]
- Brooks, R.; Bussière, L.F.; Jennions, M.D.; Hunt, J. Sinister Strategies Succeed at the Cricket World Cup. Proc. Biol. Sci. 2004, 271 (Suppl. 3), S64–S66. [Google Scholar] [CrossRef]
- Breznik, K. On the Gender Effects of Handedness in Professional Tennis. J. Sports Sci. Med. 2013, 12, 346–353. [Google Scholar]
- Hagemann, N. The Advantage of Being Left-Handed in Interactive Sports. Atten. Percept. Psychophys. 2009, 71, 1641–1648. [Google Scholar] [CrossRef]
- Holtzen, D.W. Handedness and Professional Tennis. Int. J. Neurosci. 2000, 105, 101–119. [Google Scholar] [CrossRef]
- Loffing, F.; Hagemann, N.; Strauss, B. The Serve in Professional Men’s Tennis: Effects of Players’ Handedness. Int. J. Perform. Anal. Sport 2009, 9, 255–274. [Google Scholar] [CrossRef]
- Loffing, F.; Hagemann, N.; Strauss, B. Automated Processes in Tennis: Do Left-Handed Players Benefit from the Tactical Preferences of Their Opponents? J. Sports Sci. 2010, 28, 435–443. [Google Scholar] [CrossRef]
- Loffing, F.; Hagemann, N.; Strauss, B. Left-Handedness in Professional and Amateur Tennis. PLoS ONE 2012, 7, e49325. [Google Scholar] [CrossRef]
- Wood, C.J.; Aggleton, J.P. Handedness in “fast Ball” Sports: Do Left-Handers Have an Innate Advantage? Br. J. Psychol. Lond. Engl. 1953 1989, 80, 227–240. [Google Scholar] [CrossRef]
- Coren, S. The Left-Hander Syndrome: The Causes and Consequences of Left-Handedness; Free Press: New York, NY, USA, 1992; p. 308. ISBN 978-0-02-906682-9. [Google Scholar]
- Papadatou-Pastou, M.; Ntolka, E.; Schmitz, J.; Martin, M.; Munafò, M.R.; Ocklenburg, S.; Paracchini, S. Human Handedness: A Meta-Analysis. Psychol. Bull. 2020, 146, 481–524. [Google Scholar] [CrossRef]
- Grouios, G. Motoric Dominance and Sporting Excellence: Training versus Heredity. Percept. Mot. Skills 2004, 98, 53–66. [Google Scholar] [CrossRef]
- Marzoli, D.; Prete, G.; Tommasi, L. Perceptual Asymmetries and Handedness: A Neglected Link? Front. Psychol. 2014, 5, 163. [Google Scholar] [CrossRef]
- Marzoli, D.; D’Anselmo, A.; Malatesta, G.; Lucafò, C.; Prete, G.; Tommasi, L. The Intricate Web of Asymmetric Processing of Social Stimuli in Humans. Symmetry 2022, 14, 1096. [Google Scholar] [CrossRef]
- Schorer, J.; Loffing, F.; Hagemann, N.; Baker, J. Human Handedness in Interactive Situations: Negative Perceptual Frequency Effects Can Be Reversed! J. Sports Sci. 2012, 30, 507–513. [Google Scholar] [CrossRef]
- Daly, M.; Wilson, M. Killing the Competition: Female/Female and Male/Male Homicide. Hum. Nat. Hawthorne N 1990, 1, 81–107. [Google Scholar] [CrossRef] [PubMed]
- Baker, J.; Kungl, A.-M.; Pabst, J.; Strauß, B.; Büsch, D.; Schorer, J. Your Fate Is in Your Hands? Handedness, Digit Ratio (2D:4D), and Selection to a National Talent Development System. Laterality 2013, 18, 710–718. [Google Scholar] [CrossRef] [PubMed]
- Barrenetxea-Garcia, J.; Torres-Unda, J.; Esain, I.; Gil, S.M. Relative Age Effect and Left-Handedness in World Class Water Polo Male and Female Players. Laterality 2019, 24, 259–273. [Google Scholar] [CrossRef] [PubMed]
- Connor, J.D.; Mann, D.L.; Gomez, M.-A.; Leicht, A.S.; Doma, K. Performance Advantages of Left-Handed Cricket Batting Talent. Front. Psychol. 2020, 11, 1654. [Google Scholar] [CrossRef]
- Schouten, B.; Troje, N.F.; Brooks, A.; van der Zwan, R.; Verfaillie, K. The Facing Bias in Biological Motion Perception: Effects of Stimulus Gender and Observer Sex. Atten. Percept. Psychophys. 2010, 72, 1256–1260. [Google Scholar] [CrossRef]
- Troje, N.F.; Sadr, J.; Geyer, H.; Nakayama, K. Adaptation Aftereffects in the Perception of Gender from Biological Motion. J. Vis. 2006, 6, 850–857. [Google Scholar] [CrossRef]
- van der Zwan, R.; Machatch, C.; Kozlowski, D.; Troje, N.F.; Blanke, O.; Brooks, A. Gender Bending: Auditory Cues Affect Visual Judgements of Gender in Biological Motion Displays. Exp. Brain Res. 2009, 198, 373–382. [Google Scholar] [CrossRef]
- Vanrie, J.; Dekeyser, M.; Verfaillie, K. Bistability and Biasing Effects in the Perception of Ambiguous Point-Light Walkers. Perception 2004, 33, 547–560. [Google Scholar] [CrossRef]
- Brooks, A.; Schouten, B.; Troje, N.F.; Verfaillie, K.; Blanke, O.; van der Zwan, R. Correlated Changes in Perceptions of the Gender and Orientation of Ambiguous Biological Motion Figures. Curr. Biol. CB 2008, 18, R728–R729. [Google Scholar] [CrossRef]
- Simmons, J.P.; Nelson, L.D.; Simonsohn, U. A 21 Word Solution. Dialogue Off. Newsl. Soc. Personal. Soc. Psychol. 2012, 26, 4–7. [Google Scholar] [CrossRef]
- Liu, C.-H.; Tzeng, O.J.L.; Hung, D.L.; Tseng, P.; Juan, C.-H. Investigation of Bistable Perception with the “Silhouette Spinner”: Sit Still, Spin the Dancer with Your Will. Vision Res. 2012, 60, 34–39. [Google Scholar] [CrossRef]
- Salmaso, D.; Longoni, A.M. Problems in the Assessment of Hand Preference. Cortex J. Devoted Study Nerv. Syst. Behav. 1985, 21, 533–549. [Google Scholar] [CrossRef]
- Watling, D.; Woekman, L.; Bourne, V.J. Emotion Lateralisation: Developments throughout the Lifespan. Laterality 2012, 17, 2012. [Google Scholar]
- Behrmann, M.; Avidan, G.; Leonard, G.L.; Kimchi, R.; Luna, B.; Humphreys, K.; Minshew, N. Configural Processing in Autism and Its Relationship to Face Processing. Neuropsychologia 2006, 44, 110–129. [Google Scholar] [CrossRef]
- Reed, C.L.; Stone, V.E.; Bozova, S.; Tanaka, J. The Body-Inversion Effect. Psychol. Sci. 2003, 14, 302–308. [Google Scholar] [CrossRef]
- Reed, C.L.; Stone, V.E.; Grubb, J.D.; McGoldrick, J.E. Turning Configural Processing Upside down: Part and Whole Body Postures. J. Exp. Psychol. Hum. Percept. Perform. 2006, 32, 73–87. [Google Scholar] [CrossRef]
- Dundas, E.M.; Best, C.A.; Minshew, N.J.; Strauss, M.S. A Lack of Left Visual Field Bias When Individuals with Autism Process Faces. J. Autism Dev. Disord. 2012, 42, 1104–1111. [Google Scholar] [CrossRef]
- Taylor, S.; Workman, L.; Yeomans, H. Abnormal Patterns of Cerebral Lateralisation as Revealed by the Universal Chimeric Faces Task in Individuals with Autistic Disorder. Laterality 2012, 17, 428–437. [Google Scholar] [CrossRef]
- Gilbert, C.; Bakan, P. Visual Asymmetry in Perception of Faces. Neuropsychologia 1973, 11, 355–362. [Google Scholar] [CrossRef]
- Wolff, W. The Experimental Study of Forms of Expression. Character Personal. Q. Psychodiagnostic Allied Stud. 1933, 2, 168–176. [Google Scholar] [CrossRef]
- Malatesta, G.; Marzoli, D.; Tommasi, L. Keep a Left Profile, Baby! The Left-Cradling Bias Is Associated with a Preference for Left-Facing Profiles of Human Babies. Symmetry 2020, 12, 911. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factors | df | F | Sig. | ηp2 |
|---|---|---|---|---|
| participant’s sex | 1,50 | 1.501 | 0.226 | 0.029 |
| figure’s sex | 1,50 | 0.556 | 0.459 | 0.011 |
| figure’s sex X participant’s sex | 1,50 | 2.322 | 0.134 | 0.044 |
| type of rotation | 1,50 | 2.342 | 0.132 | 0.045 |
| type of rotation X participant’s sex | 1,50 | 0.042 | 0.839 | 0.001 |
| figure’s sex X type of rotation | 1,50 | 0.440 | 0.510 | 0.009 |
| figure’s sex X type of rotation X participant’s sex | 1,50 | 3.581 | 0.064 | 0.067 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lucafò, C.; Marzoli, D.; Ferrara, C.; Bertollo, M.; Tommasi, L. No Sex Differences in the Attentional Bias for the Right Side of Human Bodies. Symmetry 2023, 15, 466. https://doi.org/10.3390/sym15020466
Lucafò C, Marzoli D, Ferrara C, Bertollo M, Tommasi L. No Sex Differences in the Attentional Bias for the Right Side of Human Bodies. Symmetry. 2023; 15(2):466. https://doi.org/10.3390/sym15020466
Chicago/Turabian StyleLucafò, Chiara, Daniele Marzoli, Cosimo Ferrara, Maurizio Bertollo, and Luca Tommasi. 2023. "No Sex Differences in the Attentional Bias for the Right Side of Human Bodies" Symmetry 15, no. 2: 466. https://doi.org/10.3390/sym15020466
APA StyleLucafò, C., Marzoli, D., Ferrara, C., Bertollo, M., & Tommasi, L. (2023). No Sex Differences in the Attentional Bias for the Right Side of Human Bodies. Symmetry, 15(2), 466. https://doi.org/10.3390/sym15020466