

Phenotypic Variation in a Species Range: Another Look (Developmental Stability Study of the Meristic Variation in the Sand Lizard Lacerta agilis)


{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
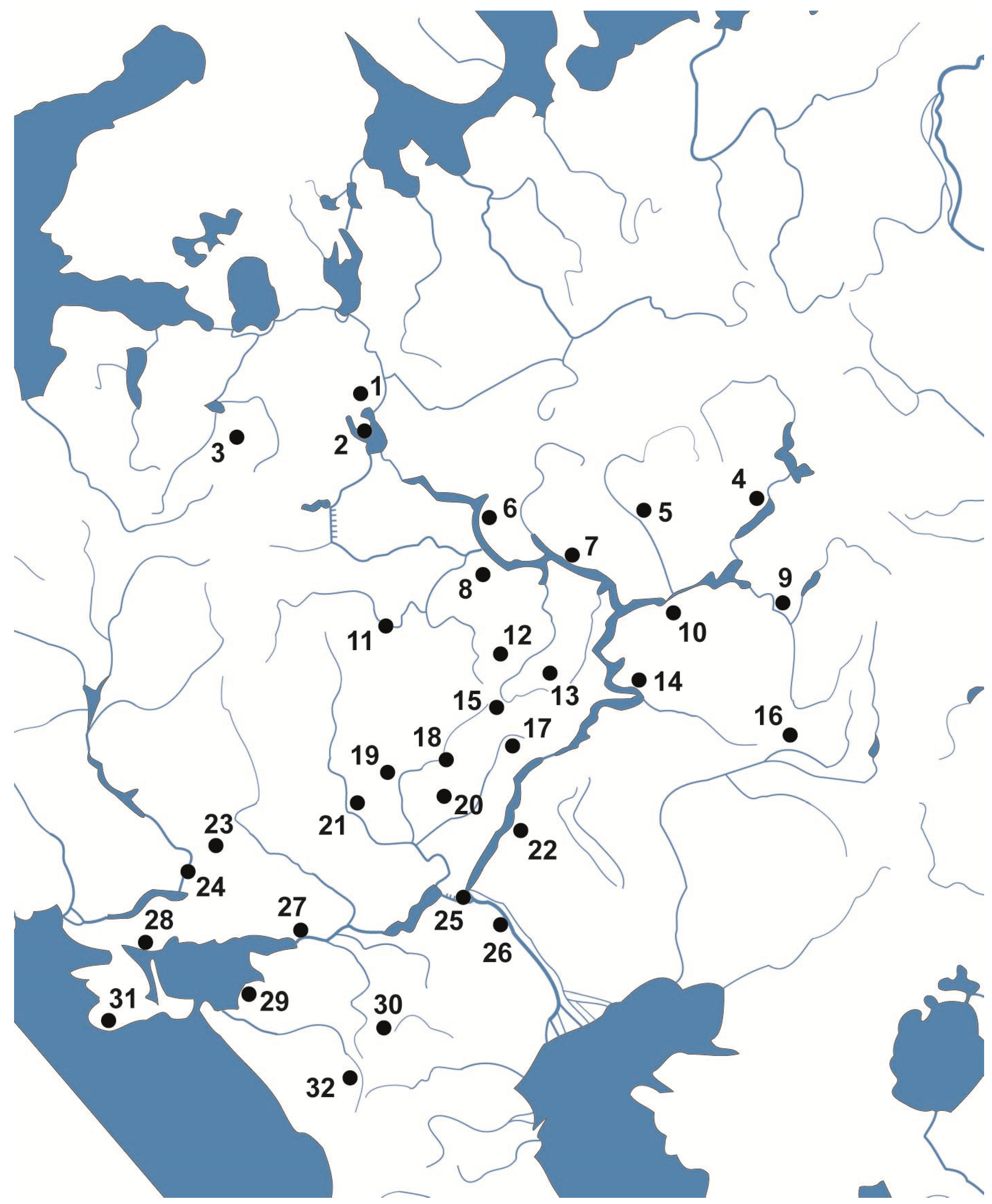
2. Material and Methods
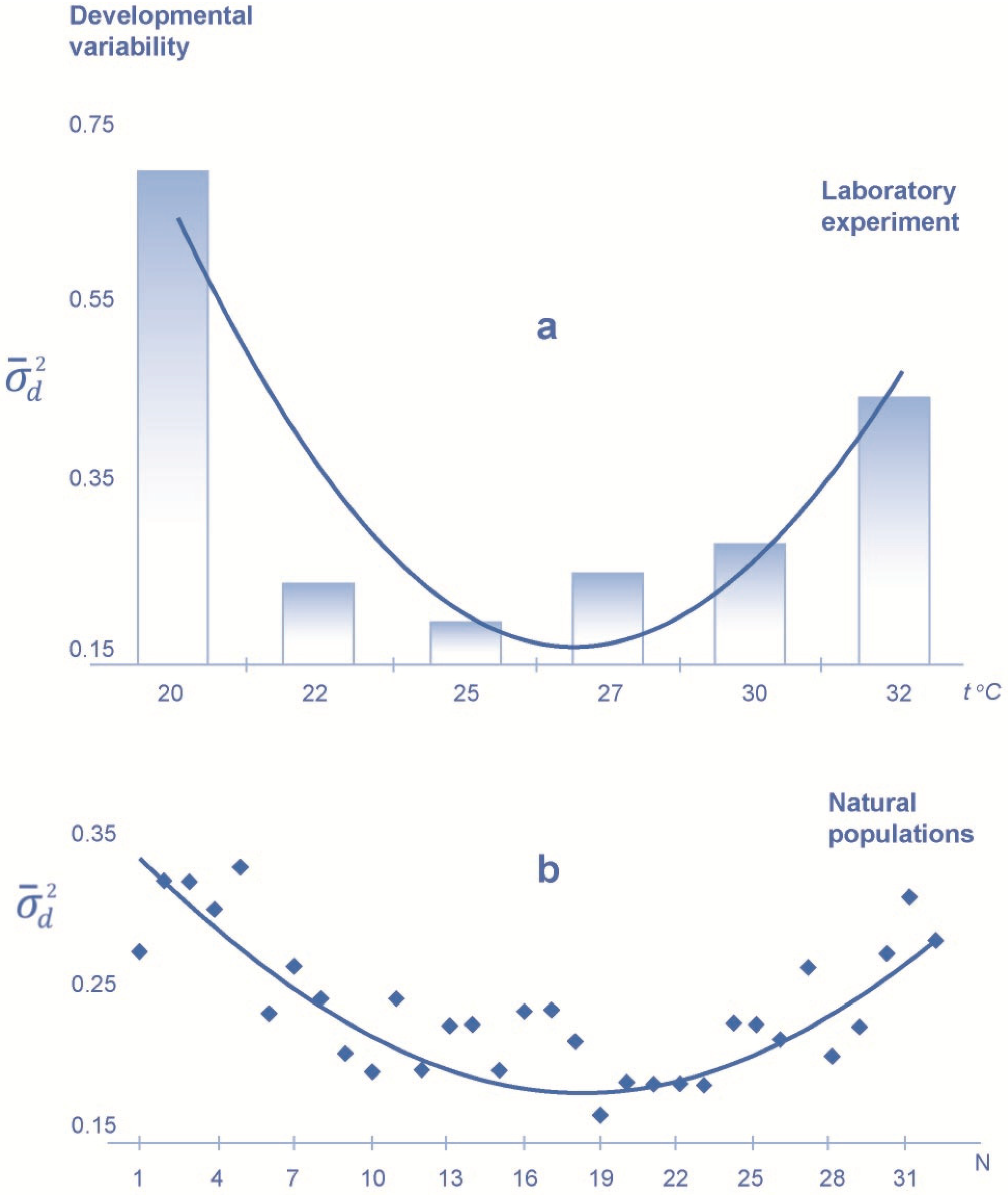
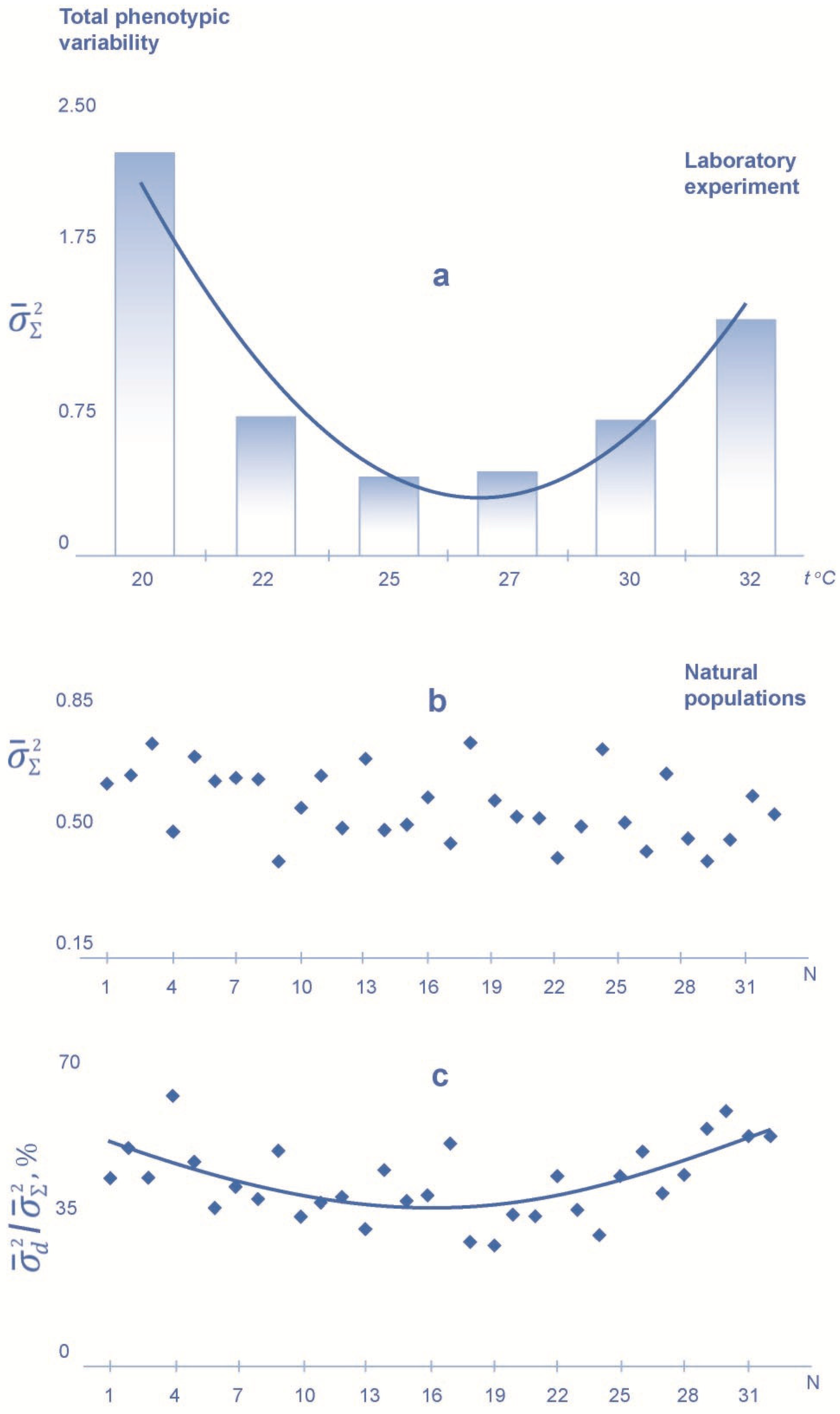
3. Results
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Leary, R.F.; Allendorf, F.W.; Knudsen, K.L. Genetic, environmental, and developmental causes of meristic variation in rainbow trout. Acta Zool. Fenn. 1992, 191, 79–95. [Google Scholar]
- Scheiner, S.M. Genetics and evolution of phenotypic plasticity. Annu. Rev. Ecol. Syst. 1993, 24, 35–68. [Google Scholar] [CrossRef]
- Gilbert, S.F. Ecological Developmental Biology: Developmental Biology Meets the Real World. Dev. Biol. 2001, 233, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, S.F.; Barresi, M.J.F. Developmental Biology, 11th ed.; Sinauer Associates: Sunderland, MA, USA, 2016. [Google Scholar]
- Soule, M.; Baker, B. Phenetics of natural populations IV. The population asymmetry parameter in the butterfly Coenonympha tullia. Heredity 1968, 23, 611–614. [Google Scholar] [CrossRef] [PubMed]
- Kat, P.W. The relationship between heterozygosity for enzyme loci and developmental homeostasis in peripheral populations of aquatic bivalves (Unionidae). Am. Nat. 1982, 119, 824–832. [Google Scholar] [CrossRef]
- Parsons, P.A. Fluctuating asymmetry: A biological monitor of environmental and genomic stress. Heredity 1992, 68, 361–364. [Google Scholar] [CrossRef] [PubMed]
- Mather, K. Genetical control of stability in development. Heredity 1953, 7, 297–336. [Google Scholar] [CrossRef]
- Waddington, C.H. The Strategy of the Genes; George Allen & Unwin: London, UK, 1957; 262p. [Google Scholar]
- Yedier, S.; Bostanci, D.; Kontas, S.; Kurucu, G.; Polat, N. Comparison of otolith mass asymmetry in two different Solea solea populations in Mediterranean Sea. Ordu Üniversitesi Bilim Ve Teknol. Derg. 2018, 8, 125–133. [Google Scholar]
- Yedier, S.; Bostanci, D.; Kontas, S.; Kurucu, G.; Polat, N. Fluctuating asymmetry in otolith dimensions of Trachurus mediterraneus collected from the Middle Black Sea. Acta Biol. Turc. 2018, 31, 152–159. [Google Scholar]
- Zakharov, V.M.; Shadrina, E.G.; Trofimov, I.E. Fluctuating Asymmetry, Developmental Noise and Developmental Stability: Future Prospects for the Population Developmental Biology Approach. Symmetry 2020, 12, 1376. [Google Scholar] [CrossRef]
- Graham, J.H. Nature, Nurture, and Noise: Developmental Instability, Fluctuating Asymmetry, and the Causes of Phenotypic Variation. Symmetry 2021, 13, 1204. [Google Scholar] [CrossRef]
- Maldonado-López, Y.; Prieto-Dueñas, I.S.; Tapia-Torres, Y.; Magno, A.Z.B.; Suazo-Ortuño, I.; Cuevas-Reyes, P. Fluctuating asymmetry and oxidative stress indicate environmental stress of Cane toads Rhinella marina. Zool. Anz. 2022, 299, 234–242. [Google Scholar] [CrossRef]
- Zhelev, Z.; Tsonev, S.; Boyadzhiev, P. Using of fluctuating asymmetry in adult Pelophylax ridibundus (Amphibia: Anura: Ranidae) meristic traits as a method for assessing developmental stability of population and environmental quality of their habitat: Industrial area in southern Bulgaria. Turk. J. Zool. 2022, 46, 220–227. [Google Scholar] [CrossRef]
- Zakharov, V.M. Future Prospects for Population Phenogenetics; Soviet Scientific Reviews, Section F, Physiology and General Biology Reviews; Routledge: London, UK, 1989; Volume 4, pp. 1–80. [Google Scholar]
- Van Valen, L. A study of fluctuating asymmetry. Evolution 1962, 16, 125–142. [Google Scholar] [CrossRef]
- Soule, M.E. Phenetics of Natural Populations. II. Asymmetry and Evolution in a Lizard. Am. Nat. 1967, 101, 141–160. [Google Scholar] [CrossRef]
- Palmer, A.R.; Strobeck, C. Fluctuating asymmetry: Measurement, Analysis, Patterns. Annu. Rev. Ecol. Syst. 1986, 17, 391–421. [Google Scholar] [CrossRef]
- Zakharov, V.M.; Pankakoski, E.; Sheftel, B.I.; Peltonen, A.; Hanski, I. Developmental stability and population dynamics in the common shrew, Sorex araneus. Am. Nat. 1991, 138, 797–810. [Google Scholar] [CrossRef]
- Zhivotovsky, L.A. A measure of fluctuating asymmetry for a set of characters. Acta Zool. Fenn. 1992, 191, 73–77. [Google Scholar]
- Zakharov, V.M. Population phenogenetics: Analysis of developmental stability in natural populations. Acta Zool. Fenn. 1992, 191, 7–30. [Google Scholar]
- Graham, J.H.; Raz, S.; Hel-Or, H.; Nevo, E. Fluctuating asymmetry: Methods, theory, and applications. Symmetry 2010, 2, 466–540. [Google Scholar] [CrossRef]
- Astauroff, B.L. Analyse der erblichen Störungsfälle der bilateralen Symmetrie. Z. Ver-Erbungslehre 1930, 55, 183–262. [Google Scholar] [CrossRef]
- Bader, R.S. Fluctuating Asymmetry in the Dentition of the House Mouse. Growth 1965, 29, 291–300. [Google Scholar] [PubMed]
- Angus, R.A. Quantifying Fluctuating Asymmetry: Not All Methods are Equivalent. Growth 1982, 46, 337–342. [Google Scholar]
- Pertoldi, C.; Kristensen, T.N.; Loeschcke, V. A new method for estimating environmental variability for clonal organisms, and the use of fluctuating asymmetry as an indicator of developmental Instability. J. Theor. Biol. 2001, 210, 407–410. [Google Scholar] [CrossRef] [PubMed]
- Lajus, D.L.; Graham, J.H.; Kozhara, A.V. Developmental instability and the stochastic component of total phenotypic variance. In Developmental Instability: Causes and Consequences; Polak, M., Ed.; Oxford University Press: New York, NY, USA, 2003; pp. 343–363. [Google Scholar]
- Osgood, D.W. Effects of temperature on the development of meristic characters in Natrix fasciata. Copeia 1978, 1, 33–47. [Google Scholar] [CrossRef]
- Zakharov, V.M. Appearance, fixation and stabilization of environmentally induced phenotypic changes as a microevoutionary event. Genetica 1993, 89, 227–234. [Google Scholar] [CrossRef]
- Qualls, F.J.; Shine, R. Geographic variation in lizard phenotypes: Importance of the incubation environment. Biol. J. Linn. Soc. 1998, 64, 477–491. [Google Scholar] [CrossRef]
- Beardmore, J.A. Developmental stability in constant and fluctuating temperatures. Heredity 1960, 14, 411–422. [Google Scholar] [CrossRef]
- Freeman, G.H. Statistical methods for the analysis of genotype-environment interactions. Heredity 1973, 31, 339–354. [Google Scholar] [CrossRef]
- Glotov, N.V. Analysis of the genotype-environment interaction in natural populations. Acta Zool. Fenn. 1992, 191, 47–55. [Google Scholar]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zakharov, V.M.; Zhdanova, N.P.; Trofimov, I.E. Phenotypic Variation in a Species Range: Another Look (Developmental Stability Study of the Meristic Variation in the Sand Lizard Lacerta agilis). Symmetry 2022, 14, 2426. https://doi.org/10.3390/sym14112426
Zakharov VM, Zhdanova NP, Trofimov IE. Phenotypic Variation in a Species Range: Another Look (Developmental Stability Study of the Meristic Variation in the Sand Lizard Lacerta agilis). Symmetry. 2022; 14(11):2426. https://doi.org/10.3390/sym14112426
Chicago/Turabian StyleZakharov, Vladimir M., Nadezhda P. Zhdanova, and Ilya E. Trofimov. 2022. "Phenotypic Variation in a Species Range: Another Look (Developmental Stability Study of the Meristic Variation in the Sand Lizard Lacerta agilis)" Symmetry 14, no. 11: 2426. https://doi.org/10.3390/sym14112426
APA StyleZakharov, V. M., Zhdanova, N. P., & Trofimov, I. E. (2022). Phenotypic Variation in a Species Range: Another Look (Developmental Stability Study of the Meristic Variation in the Sand Lizard Lacerta agilis). Symmetry, 14(11), 2426. https://doi.org/10.3390/sym14112426