
Dynamics of an Eco-Epidemic Predator–Prey Model Involving Fractional Derivatives with Power-Law and Mittag–Leffler Kernel



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
- (a)
- In the presence of disease, the prey is divided into two compartments, namely susceptible prey and infected prey . The susceptible prey becomes infected when the individuals have contact with the infected prey. Since the density of prey and predator are assumed large enough, the infection rate due to this contact is bilinear which is symbolized by b.
- (b)
- In the presence of the predator–prey relationship, the interaction between susceptible prey, infected prey and predator is following the Rosenzweig–MacArthur model [38] with a few adjustments. The susceptible prey growth logistically with intrinsic growth rate r and environmental carrying capacity K. The infected prey competes for food with the susceptible prey but has no contribution to the growth rate of susceptible prey. Both susceptible prey and infected prey are predated following Holling type-II with the attack rate of predator on susceptible prey , the attack rate of predator on infected prey , the half-saturation constant of predator for susceptible prey and the half-saturation constant of predator for infected prey . Since both predations contribute to the predator birth, the conversion efficiency consists of two parts, i.e., the conversion efficiency of predator on susceptible prey and the conversion efficiency of predator on infected prey . It is also assumed that both infected prey and predator are reduced due to mortality following exponential decay where d is the death rate of infected prey, and a is the death rate of predator.
2. Fundamental Concepts
3. Eco-Epidemic Model in the Caputo Sense
3.1. Existence and Uniqueness
3.2. Non-Negativity and Boundedness
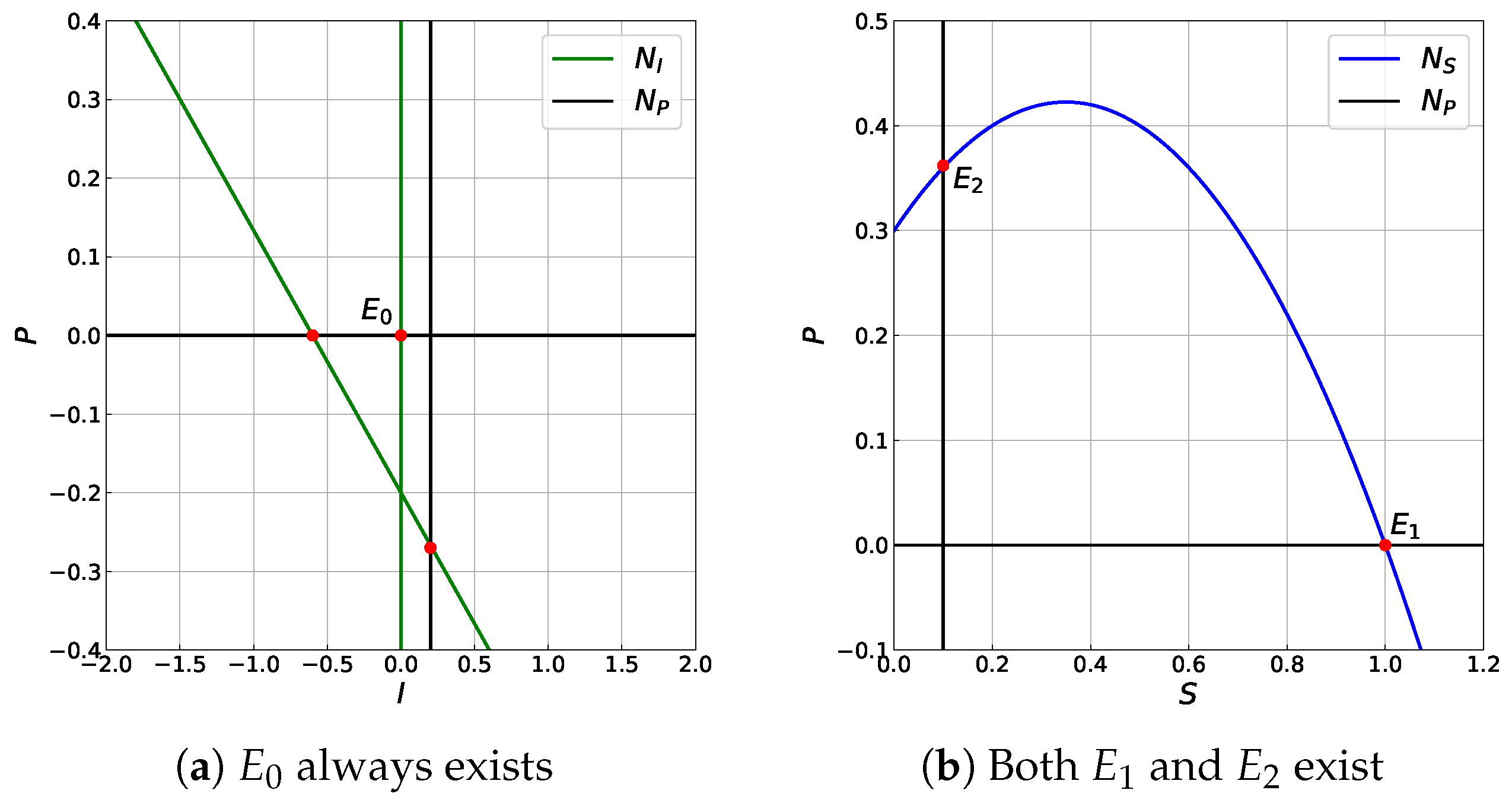
3.3. The Existence of Equilibrium Point
- The extinction of infected prey and predator point: , which always exists.
- The infected prey free point where and which exists if . The condition is equivalent to condition that the conversion rate of susceptible prey predation into the birth rate of predator is larger than the sum of the death rate of predator and its multiplication with half-saturation constant of predation.
- (i)
- If , then the co-existence point does not exist.
- (ii)
- if and
- (a)
- then there are two co-existence points, i.e., and .
- (b)
- then is the unique co-existence point.
- (iii)
- If , then there is a unique co-existence point .
- (i)
- It is clear that if then , and thus the co-existence point does not exist.
- (ii)
- if then . As a result that , we have . Furthermore, if
- (a)
- then . Therefore, we have and and .
- (b)
- then so that and .
- (iii)
- If then is the only solution for . Furthermore, if then .
3.4. Local Stability of Equilibrium Points
- (i)
- locally asymptotically stable if and .
- (ii)
- a saddle point if or .
- (i)
- If and , then and . Due to Matignon condition at Theorem 2, is locally asymptotically stable.
- (ii)
- If then . In addition, if then . Thus, Theorem 2 says that is a saddle point.
- (i)
- locally asymptotically stable if and
- (a)
- , or;
- (b)
- , and .
- (ii)
- a saddle point if
- (a)
- and , or;
- (b)
- , , , and , or;
- (c)
- , , and .
- (i)
- , , , and .
- (ii)
- , , , , and .
- (iii)
- , , , and .
- (iv)
- , , , , and .
3.5. Global Stability of Equilibrium Points
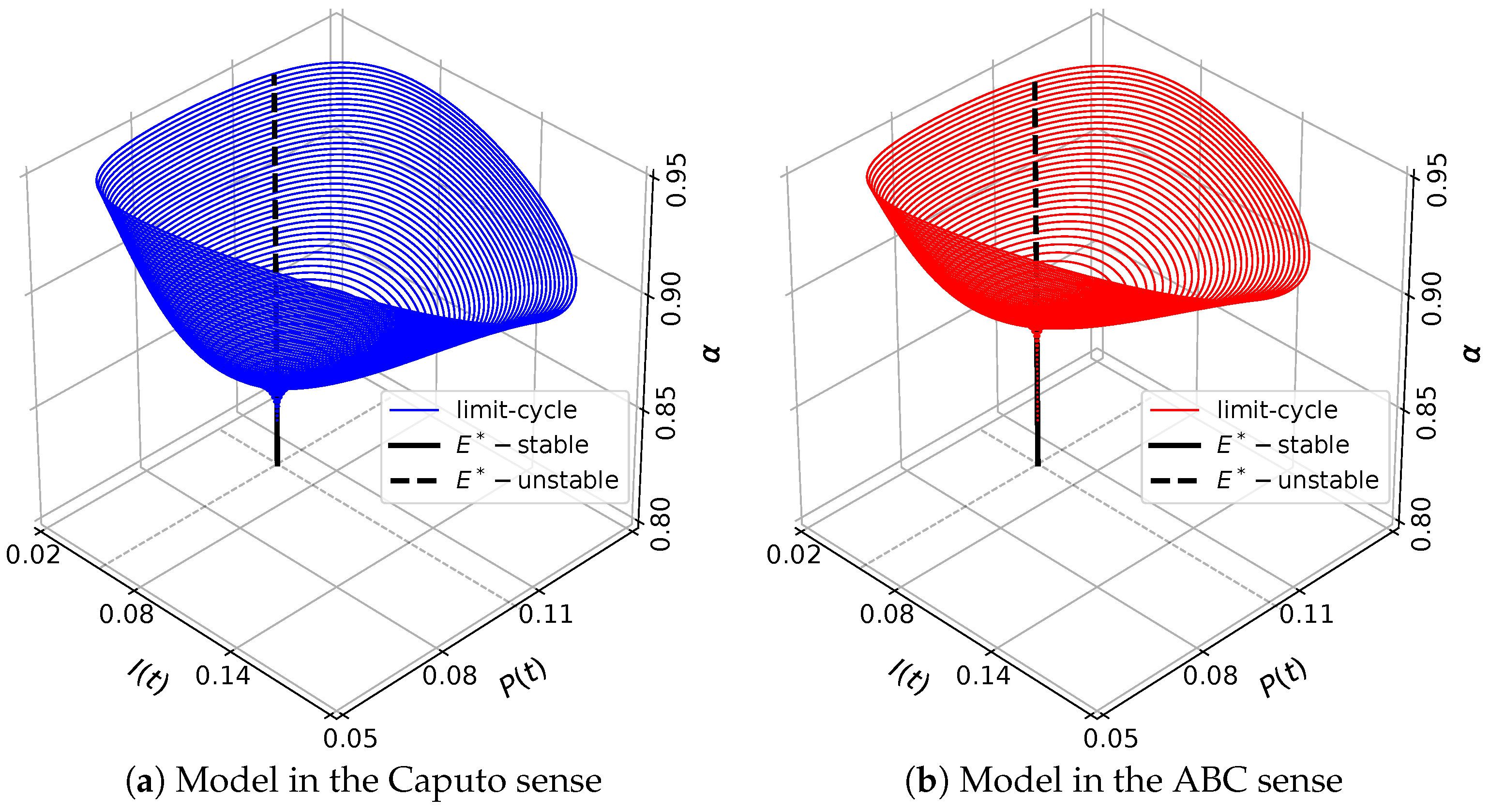
3.6. The Existence of Hopf Bifurcation
- and where ;
- ;
- .
4. Eco-Epidemic Model in the Atangana–Baleanu Sense
Existence and Uniqueness
5. Numerical Simulations
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lotka, A.J. Elements of physical biology. Nature 1925, 116, 461. [Google Scholar]
- Volterra, V. Variations and fluctuations of the number of individuals in animal species living together. ICES J. Mar. Sci. 1928, 3, 3–51. [Google Scholar] [CrossRef]
- Berryman, A.A. The orgins and evolution of predator-prey theory. Ecology 1992, 73, 1530–1535. [Google Scholar] [CrossRef]
- González-Olivares, E.; Tintinago-Ruiz, P.C.; Rojas-Palma, A. A Leslie–Gower-type predator–prey model with sigmoid functional response. Int. J. Comput. Math. 2015, 92, 1895–1909. [Google Scholar] [CrossRef]
- Wei, F.; Fu, Q. Hopf bifurcation and stability for predator-prey systems with Beddington-DeAngelis type functional response and stage structure for prey incorporating refuge. Appl. Math. Model. 2016, 40, 126–134. [Google Scholar] [CrossRef]
- Khajanchi, S. Modeling the dynamics of stage-structure predator-prey system with Monod-Haldane type response function. Appl. Math. Comput. 2017, 302, 122–143. [Google Scholar] [CrossRef]
- Song, Q.; Yang, R.; Zhang, C.; Tang, L. Bifurcation analysis in a diffusive predator–prey system with Michaelis–Menten-type predator harvesting. Adv. Differ. Equ. 2018, 2018, 329. [Google Scholar] [CrossRef]
- Suryanto, A.; Darti, I.; Panigoro, H.S.; Kilicman, A. A fractional-order predator–prey model with ratio-dependent functional response and linear harvesting. Mathematics 2019, 7, 1100. [Google Scholar] [CrossRef]
- Ghanbari, B.; Kumar, D. Numerical solution of predator-prey model with Beddington-DeAngelis functional response and fractional derivatives with Mittag-Leffler kernel. Chaos 2019, 29, 063103. [Google Scholar] [CrossRef] [PubMed]
- Manna, D.; Maiti, A.; Samanta, G.P. A Michaelis–Menten type food chain model with strong Allee effect on the prey. Appl. Math. Comput. 2017, 311, 390–409. [Google Scholar] [CrossRef]
- Dhiman, A.; Poria, S. Allee effect induced diversity in evolutionary dynamics. Chaos Soliton Fract. 2018, 108, 32–38. [Google Scholar] [CrossRef]
- Elaydi, S.; Kwessi, E.; Livadiotis, G. Hierarchical competition models with the Allee effect III: Multispecies. J. Biol. Dyn. 2018, 12, 271–287. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, L.; Bai, Y. Stability and bifurcation analysis on a predator–prey system with the weak Allee effect. Mathematics 2019, 7, 432. [Google Scholar] [CrossRef]
- Rahmi, E.; Darti, I.; Suryanto, A.; Trisilowati; Panigoro, H.S. Stability analysis of a fractional-order Leslie-Gower model with Allee Effect in predator. J. Phys. Conf. Ser. 2021, 1821, 012051. [Google Scholar] [CrossRef]
- Bodine, E.N.; Yust, A.E. Predator–prey dynamics with intraspecific competition and an Allee effect in the predator population. Lett. Biomath. 2017, 4, 23–38. [Google Scholar] [CrossRef]
- Ali, N.; Haque, M.; Venturino, E.; Chakravarty, S. Dynamics of a three species ratio-dependent food chain model with intra-specific competition within the top predator. Comput. Biol. Med. 2017, 85, 63–74. [Google Scholar] [CrossRef]
- Jana, D.; Banerjee, A.; Samanta, G. Degree of prey refuges: Control the competition among prey and foraging ability of predator. Chaos Soliton Fract. 2017, 104, 350–362. [Google Scholar] [CrossRef]
- Sieber, M.; Malchow, H.; Hilker, F.M. Disease-induced modification of prey competition in eco-epidemiological models. Ecol. Complex. 2014, 18, 74–82. [Google Scholar] [CrossRef]
- Sahoo, B. Role of additional food in eco-epidemiological system with disease in the prey. Appl. Math. Comput. 2015, 259, 61–79. [Google Scholar] [CrossRef]
- Saifuddin, M.; Biswas, S.; Samanta, S.; Sarkar, S.; Chattopadhyay, J. Complex dynamics of an eco-epidemiological model with different competition coefficients and weak Allee in the predator. Chaos Soliton Fract. 2016, 91, 270–285. [Google Scholar] [CrossRef]
- Mondal, S.; Lahiri, A.; Bairagi, N. Analysis of a fractional order eco-epidemiological model with prey infection and type 2 functional response. Math. Methods Appl. Sci. 2017, 40, 6776–6789. [Google Scholar] [CrossRef]
- Mondal, A.; Pal, A.K.; Samanta, G.P. On the dynamics of evolutionary Leslie-Gower predator-prey eco-epidemiological model with disease in predator. Ecol. Genet. Genom. 2019, 10, 100034. [Google Scholar] [CrossRef]
- Panigoro, H.S.; Suryanto, A.; Kusumawinahyu, W.M.; Darti, I. Dynamics of a fractional-order predator-prey model with infectious diseases in prey. Commun. Biomath. Sci. 2019, 2, 105–117. [Google Scholar] [CrossRef]
- Wei, C.; Chen, L. Global dynamics behaviors of viral infection model for pest management. Discrete. Dyn. Nat. Soc. 2009, 2009, 1–16. [Google Scholar] [CrossRef]
- Fu, J.; Wang, Y. The Mathematical study of pest management strategy. Discrete. Dyn. Nat. Soc. 2012, 2012, 1–19. [Google Scholar] [CrossRef]
- Sun, K.; Zhang, T.; Tian, Y. Theoretical study and control optimization of an integrated pest management predator–prey model with power growth rate. Math. Biosci. 2016, 279, 13–26. [Google Scholar] [CrossRef]
- Mandal, D.S.; Samanta, S.; Alzahrani, A.K.; Chattopadhyay, J. Study of a predator-prey model with pest management perspective. J. Biol. Syst. 2019, 27, 309–336. [Google Scholar] [CrossRef]
- Suryanto, A.; Darti, I. Dynamics of Leslie-Gower pest-predator model with disease in pest including pest-harvesting and optimal implementation of pesticide. Int. J. Math. Math. Sci. 2019, 2019, 5079171. [Google Scholar] [CrossRef]
- Connole, M.D.; Yamaguchi, H.; Elad, D.; Hasegawa, A.; Segal, E.; Torres-Rodriguez, J.M. Natural pathogens of laboratory animals and their effects on research. Med. Mycol. 2000, 38, 59–65. [Google Scholar] [CrossRef][Green Version]
- Kan, I.; Motro, Y.; Horvitz, N.; Kimhi, A.; Leshem, Y.; Yom-Tov, Y.; Nathan, R. Agricultural rodent control using barnowls: Is it profitable. Am. J. Agric. Econ. 2014, 96, 733–752. [Google Scholar] [CrossRef]
- Kross, S.M.; Bourbour, R.P.; Martinico, B.L. Agricultural land use, barn owl diet, and vertebrate pest control implications. Agric. Ecosyst. Environ. 2016, 223, 167–174. [Google Scholar] [CrossRef]
- Wendt, C.A.; Johnson, M.D. Agriculture, Ecosystems and Environment Multi-scale analysis of barn owl nest box selection on Napa Valley vineyards. Agric. Ecosyst. Environ. 2017, 247, 75–83. [Google Scholar] [CrossRef]
- Solter, L.; Hajek, A.; Lacey, L. Exploration for Entomopathogens. In Microbial Control of Insect and Mite Pests; Lacey, L.A., Ed.; Academic Press: San Diego, CA, USA, 2017; pp. 13–23. [Google Scholar]
- Wang, J.; Qu, X. Qualitative analysis for a ratio-dependent predator-prey model with disease and diffusion. Appl. Math. Comput. 2011, 217, 9933–9947. [Google Scholar] [CrossRef]
- Suryanto, A. Dynamics of an eco-epidemiological model with saturated incidence rate. AIP Conf. Proc. 2017, 1825, 020021. [Google Scholar]
- Upadhyay, R.K.; Roy, P. Spread of a Disease and its effect on population dynamics in an eco-epidemiological system. Comm. Nonlinear. Sci. Numer. Simulat. 2014, 19, 4170–4184. [Google Scholar] [CrossRef]
- Nugraheni, K.; Trisilowati, T.; Suryanto, A. Dynamics of a fractional order eco-epidemiological model. J. Trop. Life Sci. 2017, 7, 243–250. [Google Scholar] [CrossRef]
- Rosenzweig, M.L.; MacArthur, R.H. Graphical representation and stability conditions of predator-prey interactions. Am. Nat. 1963, 97, 209–223. [Google Scholar] [CrossRef]
- Panja, P. Dynamics of a fractional order predator-prey model with intraguild predation. Int. J. Model. Simul. 2019, 39, 256–268. [Google Scholar] [CrossRef]
- Morales-Delgado, V.F.; Gómez-Aguilar, J.F.; Saad, K.; Escobar Jiménez, R.F. Application of the Caputo-Fabrizio and Atangana-Baleanu fractional derivatives to mathematical model of cancer chemotherapy effect. Math. Meth. Appl. Sci. 2019, 42, 1167–1193. [Google Scholar] [CrossRef]
- El-Saka, H.A.; Lee, S.; Jang, B. Dynamic analysis of fractional-order predator–prey biological economic system with Holling type II functional response. Nonlinear Dyn. 2019, 96, 407–416. [Google Scholar] [CrossRef]
- Li, H.L.; Zhang, L.; Hu, C.; Jiang, Y.L.; Teng, Z. Dynamical analysis of a fractional-order predator-prey model incorporating a prey refuge. J. Appl. Math. Comput. 2017, 54, 435–449. [Google Scholar] [CrossRef]
- Supajaidee, N.; Moonchai, S. Stability analysis of a fractional-order two-species facultative mutualism model with harvesting. Adv. Differ. Equ. 2017, 2017, 372. [Google Scholar] [CrossRef]
- Shaikh, A.; Tassaddiq, A.; Nisar, K.S.; Baleanu, D. Analysis of differential equations involving Caputo–Fabrizio fractional operator and its applications to reaction–diffusion equations. Adv. Differ. Equ. 2019, 2019, 178. [Google Scholar] [CrossRef]
- Jajarmi, A.; Yusuf, A.; Baleanu, D.; Inc, M. A new fractional HRSV model and its optimal control: A non-singular operator approach. Phys. A 2020, 547, 123860. [Google Scholar] [CrossRef]
- Baleanu, D.; Jajarmi, A.; Mohammadi, H.; Rezapour, S. A new study on the mathematical modelling of human liver with Caputo–Fabrizio fractional derivative. Chaos Soliton Fract. 2020, 134, 109705. [Google Scholar] [CrossRef]
- Panigoro, H.S.; Suryanto, A.; Kusumawinahyu, W.M.; Darti, I. A Rosenzweig–MacArthur model with continuous threshold harvesting in predator involving fractional derivatives with power law and Mittag–Leffler kernel. Axioms 2020, 9, 122. [Google Scholar] [CrossRef]
- Panigoro, H.S.; Suryanto, A.; Kusumawinahyu, W.M.; Darti, I. Continuous threshold harvesting in a gause-type predator-prey model with fractional-order. AIP Conf. Proc. 2020, 2264, 040001. [Google Scholar]
- Suryanto, A.; Darti, I.; Anam, S. Stability analysis of a fractional order modified Leslie-Gower model with additive Allee effect. Int. J. Math. Math. Sci. 2017, 2017, 1–9. [Google Scholar] [CrossRef]
- Xie, Y.; Lu, J.; Wang, Z. Stability analysis of a fractional-order diffused prey–predator model with prey refuges. Phys. A 2019, 526, 120773. [Google Scholar] [CrossRef]
- Shah, S.A.A.; Khan, M.A.; Farooq, M.; Ullah, S.; Alzahrani, E.O. A fractional order model for Hepatitis B virus with treatment via Atangana–Baleanu derivative. Phys. A 2020, 538, 122636. [Google Scholar] [CrossRef]
- Podlubny, I. Fractional Differential Equations: An Introduction to Fractional Derivatives, Fractional Differential Equations, to Methods of Their Solution and Some of Their Applications; Academic Press: San Diego, CA, USA, 1999. [Google Scholar]
- Caputo, M. Linear models of dissipation whose Q is almost frequency independent–II. Geophys. J. Int. 1967, 13, 529–539. [Google Scholar] [CrossRef]
- Diethelm, K. The Analysis of Fractional Differential Equations: An Application-Oriented Exposition Using Differential Operators of Caputo Type; Springer: Braunschweig, Germany, 2010. [Google Scholar]
- Petras, I. Fractional-Order Nonlinear Systems: Modeling, Analysis and Simulation; Springer: Beijing, China, 2011. [Google Scholar]
- Atangana, A.; Koca, I. Chaos in a simple nonlinear system with Atangana–Baleanu derivatives with fractional order. Chaos Soliton Fract. 2016, 89, 447–454. [Google Scholar] [CrossRef]
- Yadav, S.; Pandey, R.K.; Shukla, A.K. Numerical approximations of Atangana–Baleanu Caputo derivative and its application. Chaos Soliton Fract. 2019, 118, 58–64. [Google Scholar] [CrossRef]
- Bonyah, E.; Atangana, A.; Elsadany, A.A. A fractional model for predator-prey with omnivore. Chaos 2019, 29, 013136. [Google Scholar] [CrossRef] [PubMed]
- Tajadodi, H. A Numerical approach of fractional advection-diffusion equation with Atangana–Baleanu derivative. Chaos Soliton Fract. 2020, 130, 109527. [Google Scholar] [CrossRef]
- Caputo, M.; Fabrizio, M. A new definition of fractional derivative without singular kernel. Progr. Fract. Differ. Appl. 2015, 1, 73–85. [Google Scholar]
- Li, H.; Cheng, J.; Li, H.B.; Zhong, S.M. Stability analysis of a fractional-order linear system described by the Caputo-Fabrizio derivative. Mathematics 2019, 7, 200. [Google Scholar] [CrossRef]
- Khan, S.A.; Shah, K.; Zaman, G.; Jarad, F. Existence theory and numerical solutions to smoking model under Caputo–Fabrizio fractional derivative. Chaos 2019, 29, 013128. [Google Scholar] [CrossRef]
- Atangana, A.; Khan, M.A.; Fatmawati. Modeling and analysis of competition model of bank data with fractal-fractional Caputo-Fabrizio operator. Alex. Eng. J. 2020, 59, 1985–1998. [Google Scholar] [CrossRef]
- Atangana, A.; Baleanu, D. New fractional derivatives with nonlocal and non-singular kernel: Theory and application to heat transfer model. Therm. Sci. 2016, 20, 763–769. [Google Scholar] [CrossRef]
- Fatmawati; Khan, M.A.; Azizah, M.; Windarto; Ullah, S. A fractional model for the dynamics of competition between commercial and rural banks in Indonesia. Chaos Soliton Fract. 2019, 122, 32–46. [Google Scholar] [CrossRef]
- Diethelm, K. A fractional calculus based model for the simulation of an outbreak of dengue fever. Nonlinear Dyn. 2003, 71, 613–619. [Google Scholar] [CrossRef]
- Odibat, Z.M.; Shawagfeh, N.T. Generalized Taylor’s formula. Appl. Math. Comput. 2007, 186, 286–293. [Google Scholar] [CrossRef]
- Matignon, D. Stability results for fractional differential equations with applications to control processing. Comput. Eng. Sys. appl. 1996, 2, 963–968. [Google Scholar]
- Li, Y.; Chen, Y.; Podlubny, I. Stability of fractional-order nonlinear dynamic systems: Lyapunov direct method and generalized Mittag–Leffler stability. Comput. Math. Appl. 2010, 59, 1810–1821. [Google Scholar] [CrossRef]
- Vargas-De-León, C. Volterra-type Lyapunov functions for fractional-order epidemic systems. Comm. Nonlinear Sci. Numer. Simulat. 2015, 24, 75–85. [Google Scholar] [CrossRef]
- Huo, J.; Zhao, H.; Zhu, L. The effect of vaccines on backward bifurcation in a fractional order HIV model. Nonlinear Anal. Real World Appl. 2015, 26, 289–305. [Google Scholar] [CrossRef]
- Ahmed, E.; El-Sayed, A.; El-Saka, H.A. On some Routh–Hurwitz conditions for fractional order differential equations and their applications in Lorenz, Rössler, Chua and Chen systems. Phys. Lett. A 2006, 358, 1–4. [Google Scholar] [CrossRef]
- Baisad, K.; Moonchai, S. Analysis of stability and Hopf bifurcation in a fractional Gauss-type predator–prey model with Allee effect and Holling type-III functional response. Adv. Differ. Equ. 2018, 2018, 82. [Google Scholar] [CrossRef]
- Deshpande, A.S.; Daftardar-Gejji, V.; Sukale, Y.V. On Hopf bifurcation in fractional dynamical systems. Chaos Soliton Fract. 2017, 98, 189–198. [Google Scholar] [CrossRef]
- Kuznetsov, Y.A. Elements of Applied Bifurcation Theory, 3rd ed.; Springer: New York, NY, USA, 2004. [Google Scholar]
- Li, X.; Wu, R. Hopf bifurcation analysis of a new commensurate fractional-order hyperchaotic system. Nonlinear Dyn. 2014, 78, 279–288. [Google Scholar] [CrossRef]
- Moustafa, M.; Mohd, M.H.; Ismail, A.I.; Abdullah, F.A. Stage structure and refuge effects in the dynamical analysis of a fractional order Rosenzweig-MacArthur prey-predator model. Prog. Fract. Differ. Appl. 2019, 5, 49–64. [Google Scholar] [CrossRef]
- Abdelouahab, M.S.; Hamri, N.E.; Wang, J. Hopf bifurcation and chaos in fractional-order modified hybrid optical system. Nonlinear Dyn. 2012, 69, 275–284. [Google Scholar] [CrossRef]
- Tavazoei, M.S.; Haeri, M. A proof for non existence of periodic solutions in time invariant fractional order systems. Automatica 2009, 45, 1886–1890. [Google Scholar] [CrossRef]
- Moustafa, M.; Mohd, M.H.; Ismail, A.I.; Abdullah, F.A. Dynamical analysis of a fractional-order eco-epidemiological model with disease in prey population. Adv. Differ. Equ. 2020, 2020, 48. [Google Scholar] [CrossRef]
- Diethelm, K.; Ford, N.J.; Freed, A.D. A Predictor-corrector approach for the numerical solution of fractional differential equations. Nonlinear Dyn. 2002, 29, 3–22. [Google Scholar] [CrossRef]
- Baleanu, D.; Jajarmi, A.; Hajipour, M. On the nonlinear dynamical systems within the generalized fractional derivatives with Mittag–Leffler kernel. Nonlinear Dyn. 2018, 94, 397–414. [Google Scholar] [CrossRef]













Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Panigoro, H.S.; Suryanto, A.; Kusumawinahyu, W.M.; Darti, I. Dynamics of an Eco-Epidemic Predator–Prey Model Involving Fractional Derivatives with Power-Law and Mittag–Leffler Kernel. Symmetry 2021, 13, 785. https://doi.org/10.3390/sym13050785
Panigoro HS, Suryanto A, Kusumawinahyu WM, Darti I. Dynamics of an Eco-Epidemic Predator–Prey Model Involving Fractional Derivatives with Power-Law and Mittag–Leffler Kernel. Symmetry. 2021; 13(5):785. https://doi.org/10.3390/sym13050785
Chicago/Turabian StylePanigoro, Hasan S., Agus Suryanto, Wuryansari Muharini Kusumawinahyu, and Isnani Darti. 2021. "Dynamics of an Eco-Epidemic Predator–Prey Model Involving Fractional Derivatives with Power-Law and Mittag–Leffler Kernel" Symmetry 13, no. 5: 785. https://doi.org/10.3390/sym13050785
APA StylePanigoro, H. S., Suryanto, A., Kusumawinahyu, W. M., & Darti, I. (2021). Dynamics of an Eco-Epidemic Predator–Prey Model Involving Fractional Derivatives with Power-Law and Mittag–Leffler Kernel. Symmetry, 13(5), 785. https://doi.org/10.3390/sym13050785